1. Introduction
The family Ariidae includes the hardhead catfish,
Ariopsis felis (Linnaeus 1766), and the gafftopsail catfish,
Bagre marinus (Mitchill 1815), both of which are marine catfishes common in tropical and temperate estuaries, coastal bays, and lagoons [
1]. Both species have overlapping distributions in the U.S. southeastern Atlantic and the Gulf of Mexico (GOM). While hardhead catfish and gafftopsail catfish frequent coastal waters from Cape Cod to Mexico, gafftopsail catfish also range as far south as Brazil [
2,
3,
4]. The coastal zone characteristics throughout the GOM favor high abundances of these catfishes [
3]. Accordingly, gafftopsail catfish populations support substantial fisheries across the southern GOM, particularly off Tabasco, Mexico, where the species contributes as much as 44% to the state’s total fisheries production [
5,
6]. Although the two species are relatively similar in size, gafftopsail catfish grow larger and more rapidly compared to hardhead catfish [
7].
Traditional analysis of stomach contents suggests that hardhead catfish and gafftopsail catfish share similar diets. Both are considered opportunistic feeders and omnivorous scavengers, with a diet that includes algae, sea grasses, sea anemones, gastropods, polychaetes, crustaceans, and fishes [
3,
8]. For example, Yanez-Arancibia and Lara-Dominguez [
1] noted ontogenetic shifts in diet for hardhead catfish along the Bay of Campeche, where juveniles feed on small shrimp and crabs, mollusks, and annelids, while adults feed primarily on unidentified organic matter, fishes, and crustaceans. Rudershausen and Locascio [
9] observed that gafftopsail catfish collected off Florida consume numerous prey items, ranging from benthic infauna to pelagic species, with pink shrimp (
Farfantepenaeus duorarum), amphipods, and fishes as the most important food items. In Tabasco, Mexico, Mendoza-Carranza [
5] found that crabs, fishes, stomatopods, and penaeid shrimp were common prey items for gafftopsail catfish.
Methodological advances can reveal dietary nuances often masked during visual identification of stomach contents. Along the west coast of Florida, nitrogen stable isotope analysis showed that hardhead catfish occupy a lower trophic position relative to gafftopsail catfish from the same region [
10], contrary to earlier studies suggesting dietary similarity [
3,
8]. Stable isotope analysis also indicated that hardhead catfish in Florida [
11] and Louisiana [
12] show ontogenetic stability, contrary to the ontogenetic shifts reported for hardhead catfish in Campeche [
1]. Determining whether these conflicting results reflect temporal or spatial heterogeneity in diets, or simply methodological differences, requires additional investigation.
The expansive distribution and high abundance of hardhead catfish and gafftopsail catfish throughout the coastal GOM suggest that these species fill important ecological roles. Food web models are a tool for understanding the roles of these species in coastal estuarine ecosystems. However, given a lack of detailed dietary data, even modern, comprehensive food web models (e.g., [
13]) aggregate hardhead catfish and gafftopsail catfish as “sea catfishes”. Given the shortcomings of traditional dietary analysis (e.g., the prevalence of unidentified prey items, a bias against soft-bodied prey, etc.), the objectives of this study were to: (1) investigate the dietary habits of hardhead and gafftopsail catfish by combining morphological examination and DNA barcoding of stomach contents; (2) describe the spatial and temporal variation in diet for hardhead and gafftopsail catfish. These findings will address fundamental gaps in our understanding of the roles these species fill within northern GOM ecosystems.
4. Discussion
Diet analysis aided by DNA barcoding identified distinct differences between hardhead and gafftopsail catfish in the northern GOM. These findings do not align with previous research that suggests both species share similar diets as opportunistic feeders and omnivorous scavengers. Stomach contents of hardhead catfish were dominated by crustaceans, while gafftopsail catfish showed a significantly broader dietary breadth and were primarily piscivorous. Gafftopsail catfish also consumed prey that varied widely in size and habitat, with food items ranging from benthic infauna and epifauna (such as
Lepidophthalmus louisianensis and
Callinectes sapidus, respectively) to pelagic species such as
Anchoa mitchilli. Previous studies reported that crustaceans were a primary prey item for both hardhead catfish and gafftopsail catfish [
5,
8,
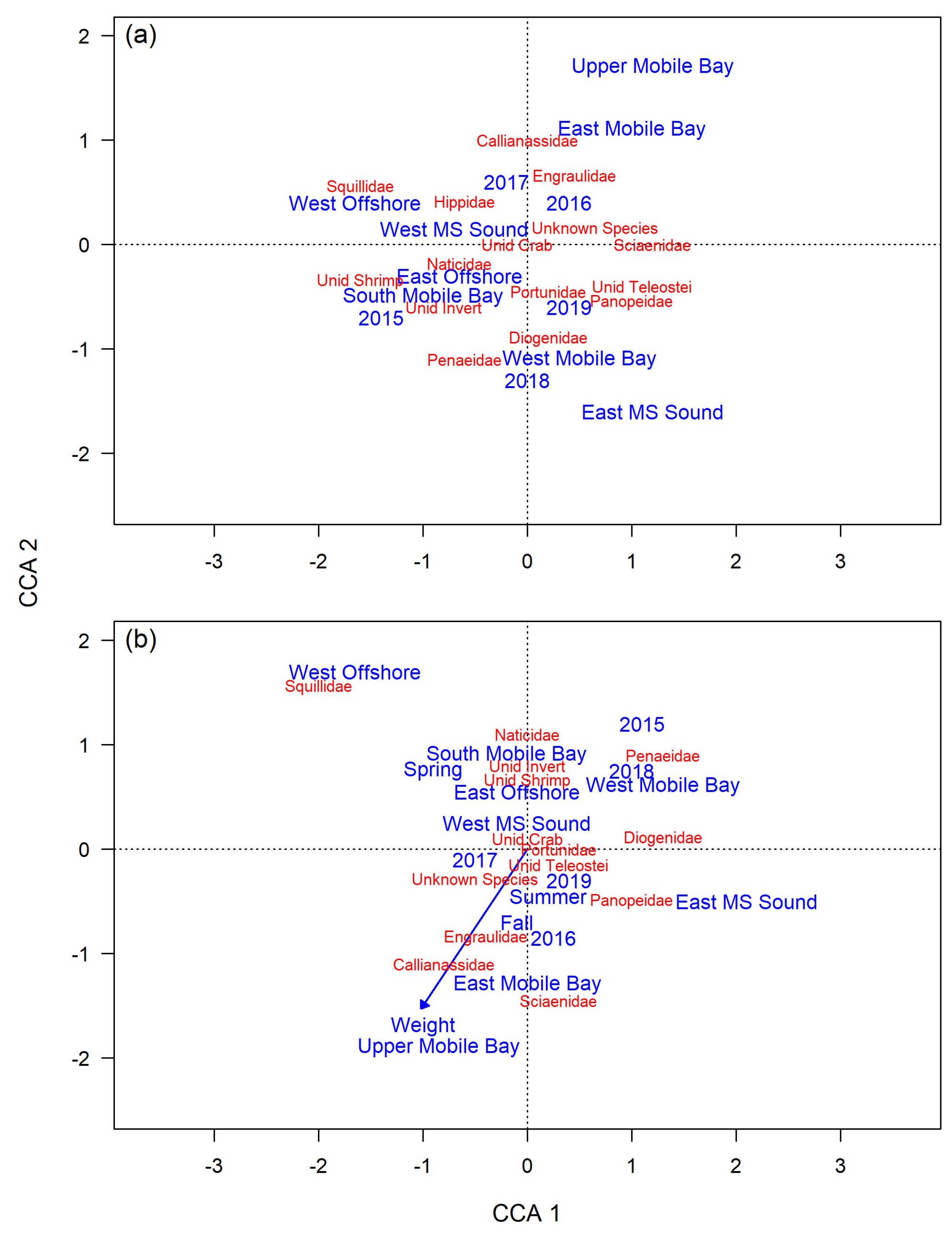
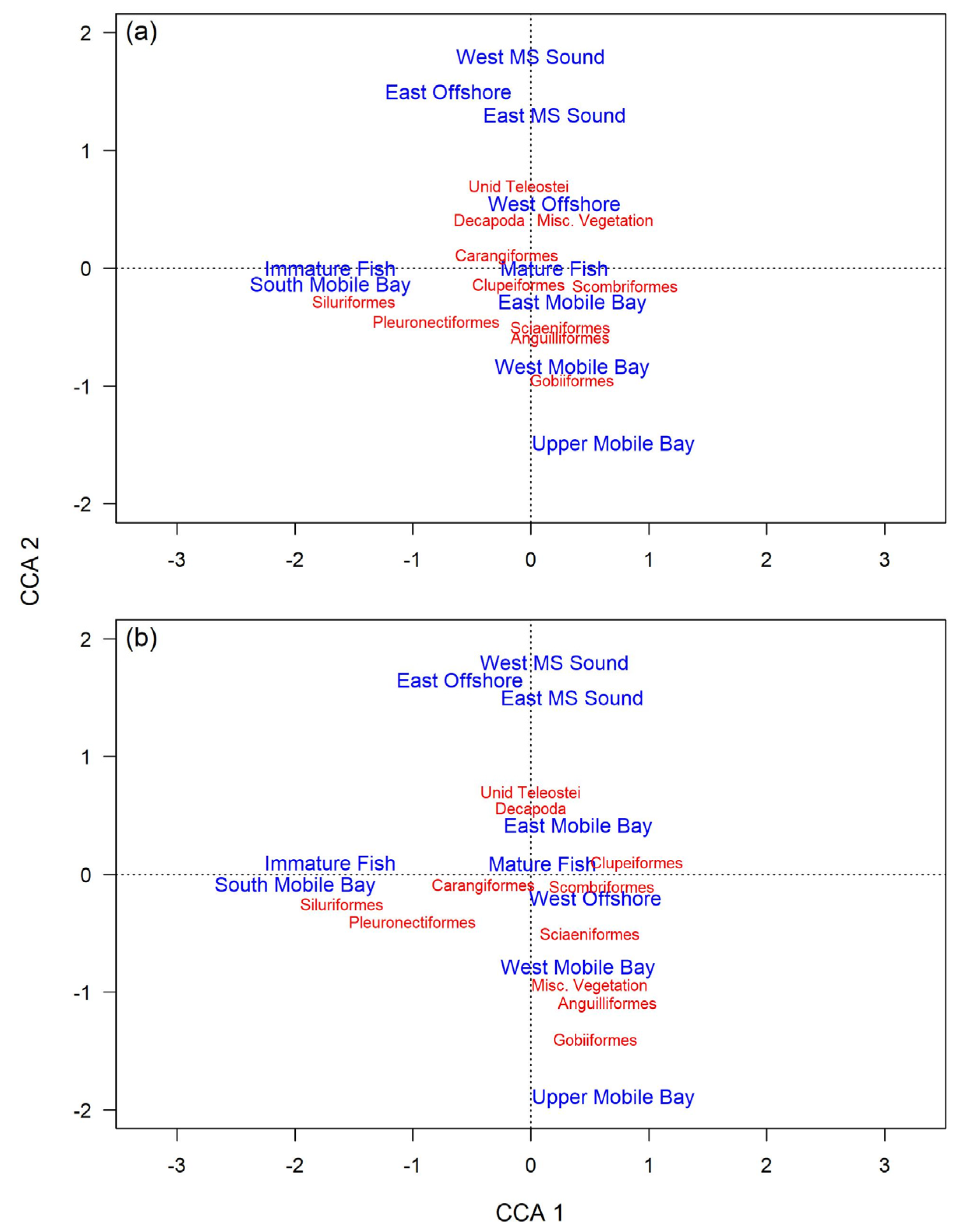
9]. In the present study, hardhead catfish CCA biplots show a high and consistent presence of crustaceans, aligning with these earlier studies. These findings support earlier studies that suggest this species is primarily dependent on meio- and macro-benthic invertebrates and epifaunal decapods. However, past results are at odds with the observed diet of gafftopsail catfish in the present study, which largely prey on fishes.
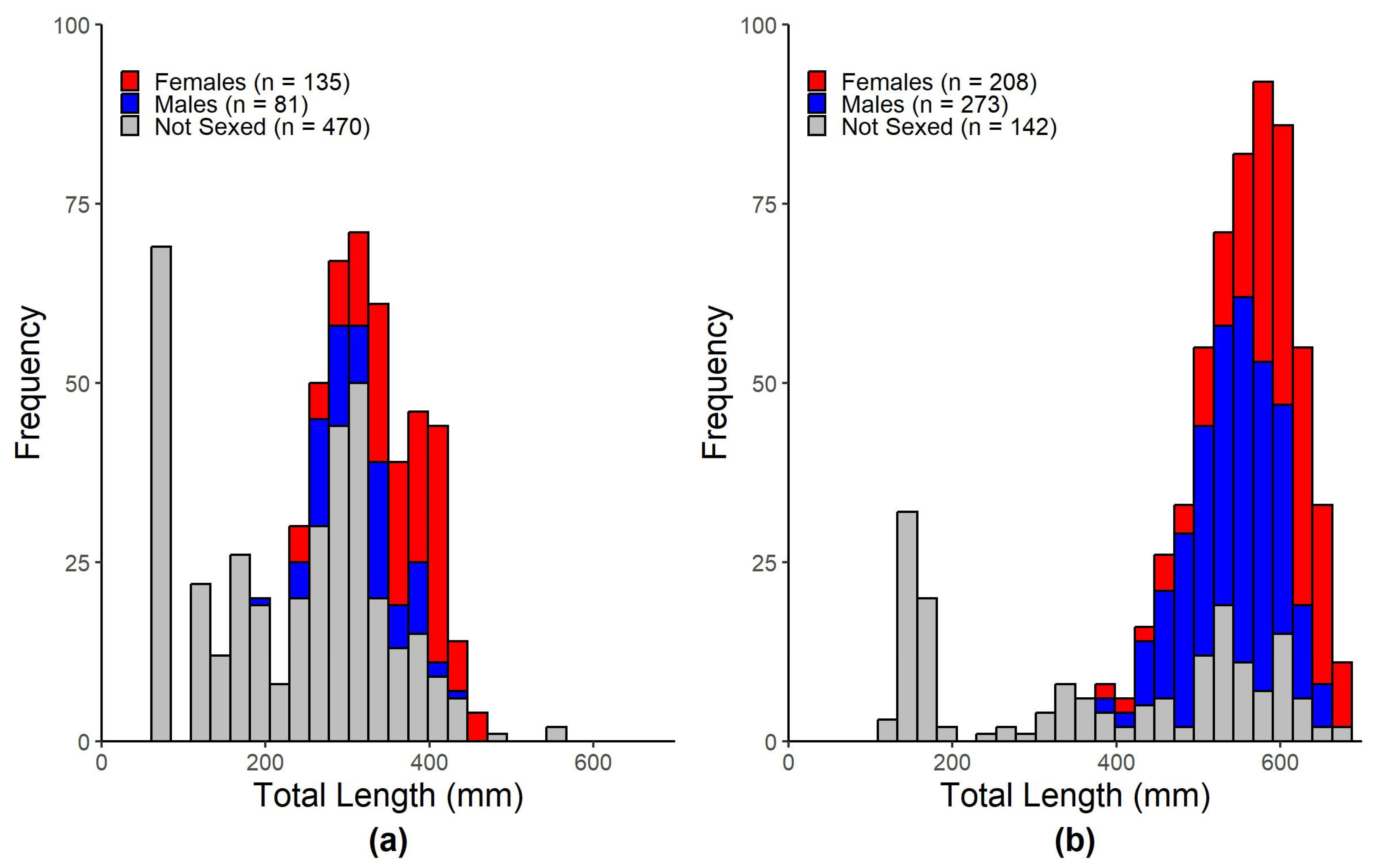
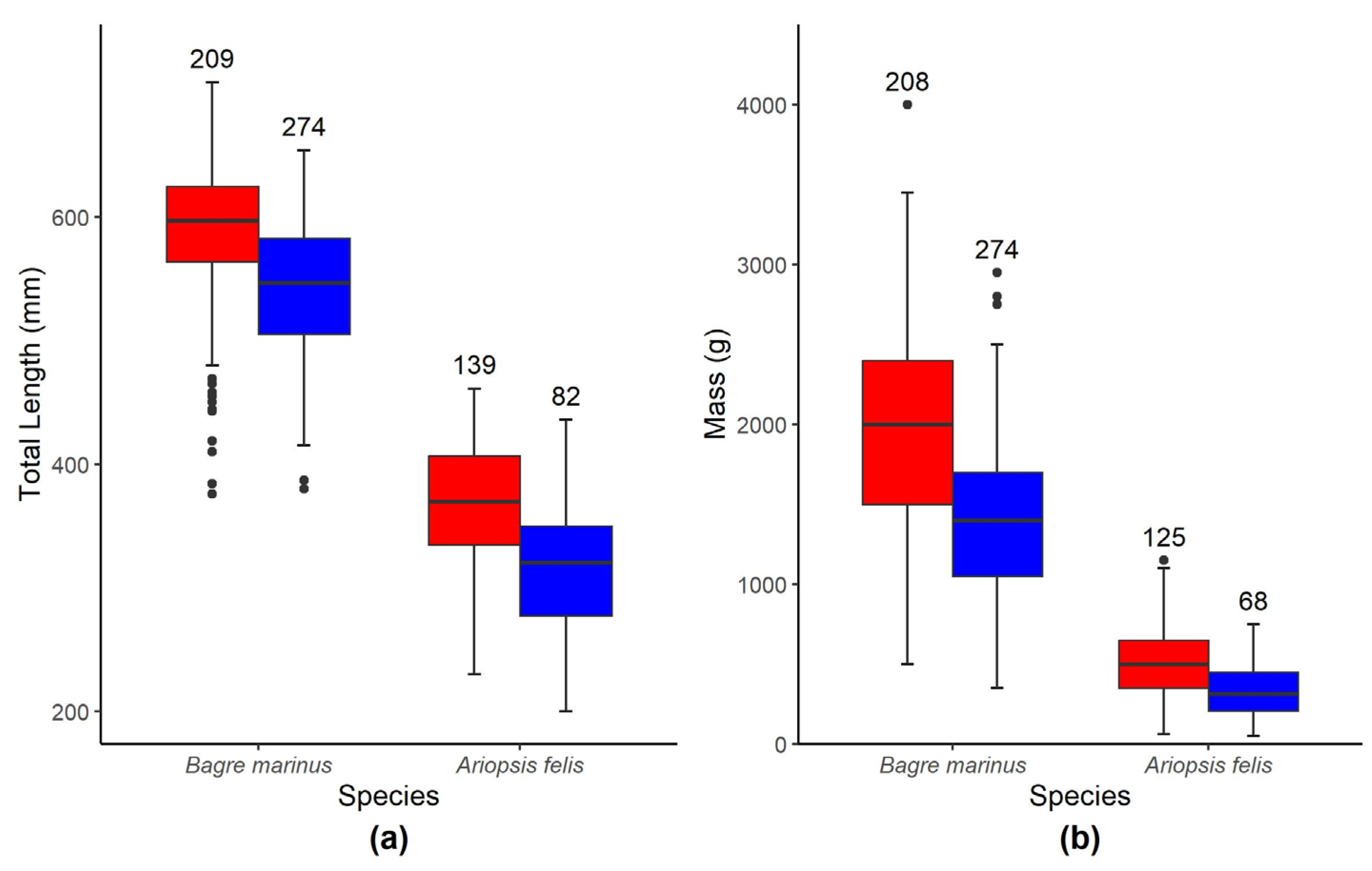
Biological differences could explain the dietary disparities between hardhead catfish and gafftopsail catfish in the present study. The mean length of gafftopsail catfish sampled in this study was more than double that of hardhead catfish, and the mean mass of gafftopsail catfish was more than five-times that of hardhead catfish. Generally, as fish size increases, so do the quantity and size range of its prey items [
34]. Thus, a larger size would explain the wider dietary breadth for gafftopsail catfish compared to hardhead catfish occupying the same habitats. Alternatively, the observed differences in diet between gafftopsail catfish and hardhead catfish in this study could be confounded by the taxonomic resolution of the diet analyses. Specifically, the inability to resolve family-level dietary habits for gafftopsail catfish may have weakened the comparison between the two catfishes. However, given the magnitude of the observed differences in prey items, it is likely that these findings reflect true differences in diet between the two species, rather than biological artifacts.
Findings from this study agree with previous hardhead catfish diet studies from the southern [
1] and eastern [
9] GOM. The significant interaction between location and year in the present study is not surprising, as the extent of the spatial-temporal overlap between hardhead catfish and their prey is reflected in their diet. For example, both estuarine ghost shrimp (
Lepidophthalmus louisianensis) and crabs of the
Callinectes genus were important to hardhead catfish diet; estuarine ghost shrimp exhibit high abundances in the northern GOM, specifically off Louisiana and Alabama [
35], and
Callinectes crabs are among the most dominant benthic macroinvertebrates along the Atlantic coast of both North and South America [
36]. These prey items exhibit broad distributions, which may contribute to the consistency in hardhead catfish diets between this study and past diet studies in other regions.
In contrast, gafftopsail catfish diet showed significant spatial differences compared to previous studies from the southern GOM. For example, Yanez-Arancibia and Lara-Domingues [
1] reported unidentified organic matter as the dominant dietary item for gafftopsail catfish, but only sampled juveniles. Similarly, Mendoza-Carranza [
5] found that fish prey were significantly less frequent than brachyuran prey, which were observed in high and consistent abundances throughout the year. However, brachyurans contributed little to gafftopsail catfish diet in the present study. Gafftopsail catfish prey items reported in the present study more closely align with observations off the west Florida coast, where fishes, amphipods, and shrimp were the most dominant prey species [
9].
A lack of complete dietary information for ariid populations in the GOM could also explain some of the diet differences between past studies and the present study. Previous research evaluating stomach contents of hardhead and gafftopsail catfish often reported large quantities of unidentifiable material or failed to identify prey items to the species level, making it difficult to elucidate differences in the diets between the two catfish species [
1,
9]. Those that did not report these difficulties were either conducted outside of the present study’s geographic range [
5] or lacked the advanced methods used herein to effectively capture diet preferences [
3,
8]. The use of DNA barcoding in the present study enabled a more comprehensive diet characterization for both catfishes by reporting higher sample sizes and more precise prey identification compared to past diet studies for either species to date in the GOM [
1,
5,
9], thus improving our understanding of the trophic importance of these catfishes in this region.
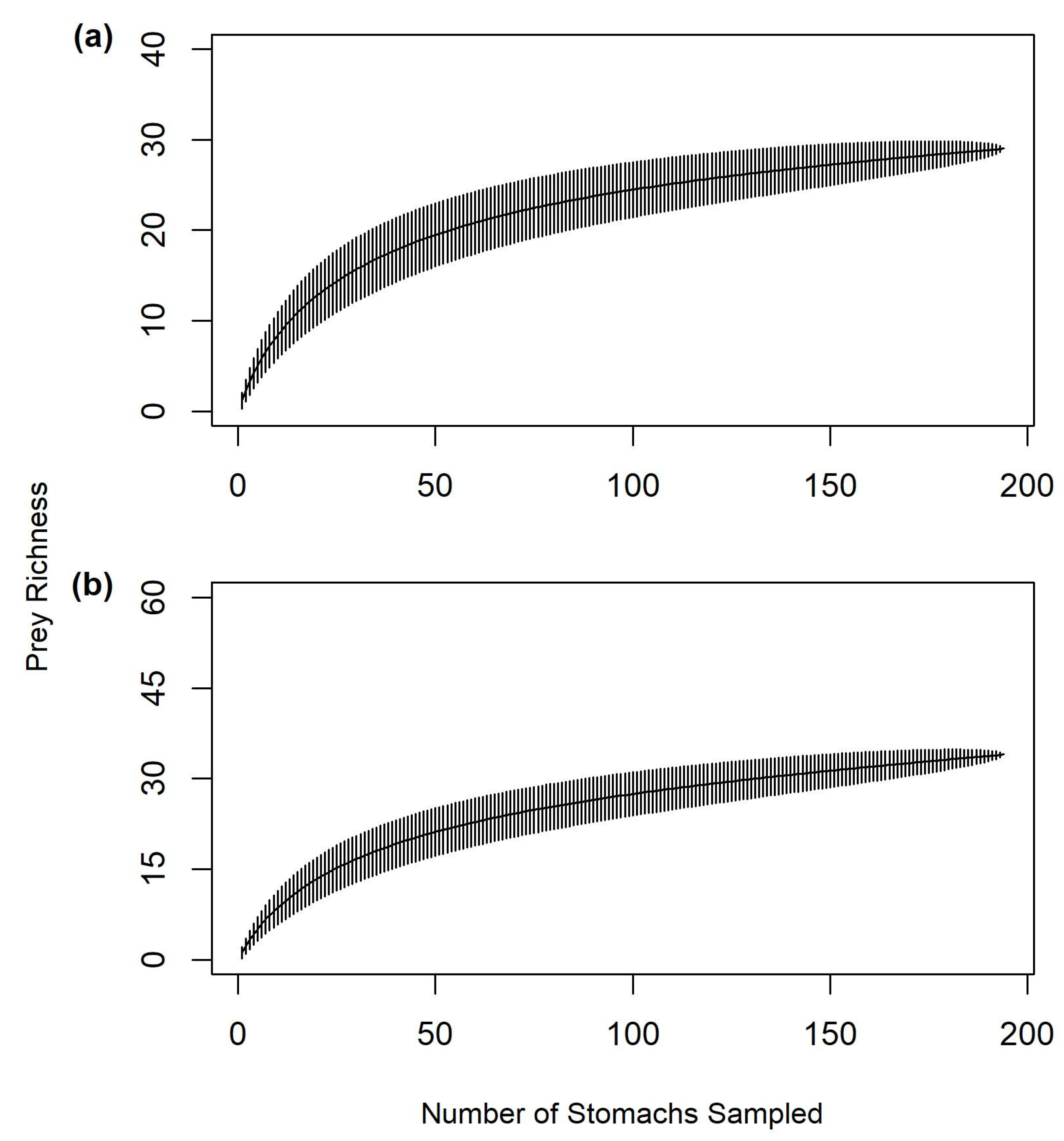
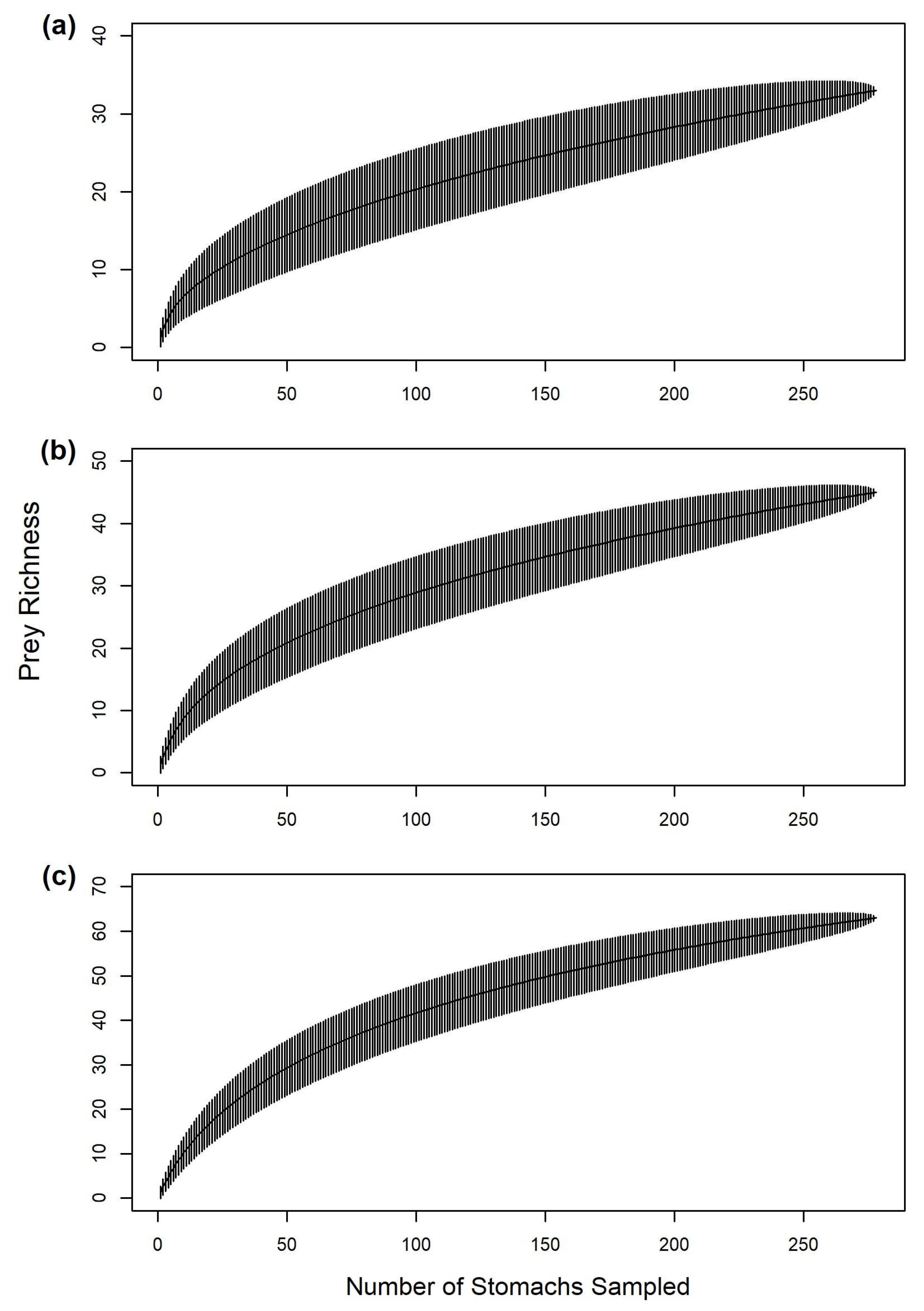
Final OTU assignment was more successful for vertebrate prey than invertebrate prey, allowing more prey items (i.e., fishes) to be identified for gafftopsail catfish than hardhead catfish. However, our ability to explain gafftopsail catfish diet variability was limited compared to hardhead catfish, which is likely due to the extensive dietary breadth observed for gafftopsail catfish in this study. Despite employing advanced techniques, some prey material still could not be identified for both hardhead and gafftopsail catfish. The substantial proportion of empty stomachs and unidentifiable (i.e., highly digested) material observed in this study for both catfishes and in past diet studies may indicate a rapid digestion rate for these species [
9]. Difficulty in identifying prey items could also be due to methodological differences; freezing and thawing samples or preserving them for later analysis [
5], rather than processing them shortly after capture [
9], may limit accurate prey identification. The freezing and thawing of some stomachs in this study likely explains why the smaller hardhead catfish sampled in this study had such a high index of vacuity. Further investigation is needed to determine the digestion rates and feeding frequency of both hardhead and gafftopsail catfish. Variation in the digestion rates could result in incorrect interpretations of dietary importance, particularly for prey items that have a high %FO but are more easily digestible and are likely under-represented in stomach content analysis [
22].
The present study provides some evidence for ontogenetic trophic shifts for gafftopsail catfish, but none for hardhead catfish, in the northern GOM. Curiously, while prey from the family Ariidae were the most important prey for immature gafftopsail catfish, they were less important for mature gafftopsail catfish. However, much of this difference can be explained by a one-time feeding event, as all the immature gafftopsail catfish that consumed other catfish were sampled from trawls performed on the same day. Rudershausen and Locascio [
9] provided evidence of ontogenetic changes in the dietary preferences among gafftopsail catfish off the west coast of Florida, although prey items for immature gafftopsail catfish were different from those observed in the present study. Pink shrimp and amphipods were most important for small gafftopsail catfish, while unidentified fishes and crabs were most important for large gafftopsail catfish. No diet shifts with ontogeny were found for hardhead catfish in the present study, which seemed to rely heavily on decapods as primary prey items regardless of catfish size. In contrast, Yanez-Arancibia and Lara-Dominguez [
1] reported that hardhead catfish diet changed with ontogeny; however, hardhead catfish were sampled outside of the geographic range of this study, the study suffered from a low (i.e., <100 individuals) sample size, and the most important food group was “unidentified organic matter” for both large (>200 mm TL) and small (<200 mm TL) individuals. Pensinger et al. [
12] concluded that these catfish showed consistent trophic niche stability off Louisiana that likely does not change with maturity, aligning with the conclusions of the present study. The high versatility and wide breadth of both hardhead and gafftopsail catfish diets suggested here–along with prey resource partitioning between these two species–likely reduces inter- and intra-species competition, contributing to the high abundances and overlapping distributions of these fishes.
A better understanding of interspecific interactions between hardhead and gafftopsail catfish and their prey–which may be recreationally or commercially valuable–is critical for developing holistic management strategies for the northern GOM [
37]. High abundances of these ariids, combined with their extensive use of estuarine and marine habitats and wide dietary breadth, make both species important contributors to ecosystem connectivity [
1,
3,
12]. Results from this study indicate that hardhead and gafftopsail catfish are closely interconnected as predator and prey, yet show distinct trophic preferences in the northern GOM. Food web models can enhance fisheries management in the northern GOM by improving our understanding of complex ecosystem processes and incorporating multispecies considerations into policy decisions. For example, management efforts aimed at reducing bycatch in the shrimp trawl fishery were unexpectedly predicted to result in decreased productivity of several commercially and recreationally valued species (e.g.,
Brevoortia patronus,
Sciaenops ocellatus, and
Lutjanus campechanus) based on an Ecopath/Ecosim model for the GOM [
37]. Rather than increasing abundances of these valued species, bycatch reduction would allow the recovery of hardhead and gafftopsail catfish and effectively increase the predation on juveniles of valued species [
37]. This counterintuitive prediction highlights the importance of evaluating potential effects of policy changes for interactions that are not yet fully understood. While model projections should be evaluated with caution, ecosystem modeling reinforces the need to establish diet composition and prey preferences for abundant predators such as hardhead and gafftopsail catfish, especially considering that minor shifts in the diets of such predators may significantly increase mortality rates of valuable juvenile species [
37].

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}