
Cleaner Fish Do Not Impact the Pigmentation of Salmon Lice (Lepeophtheirus salmonis) in Commercial Aquaculture Cages

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish, Location and Sampling
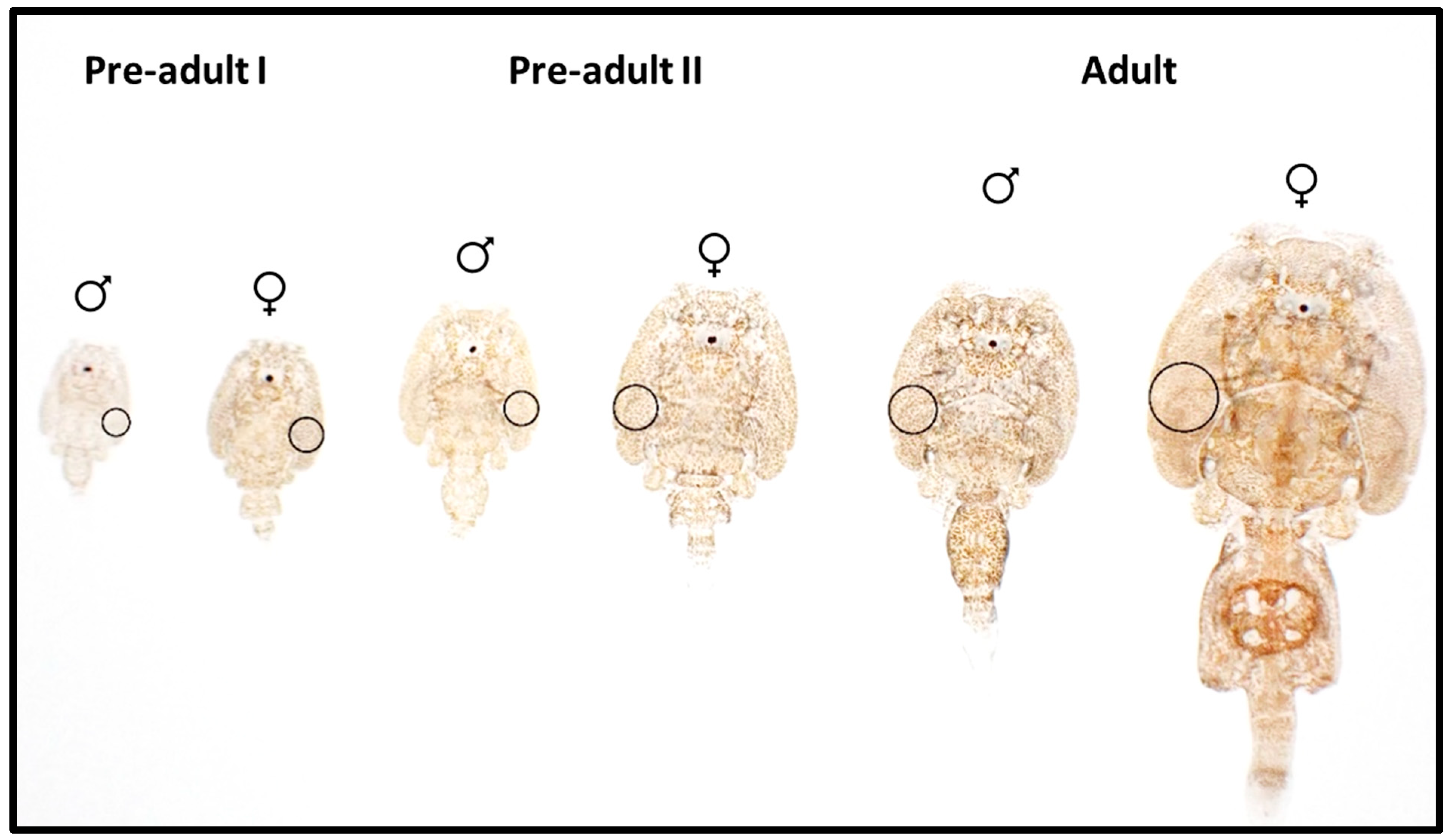
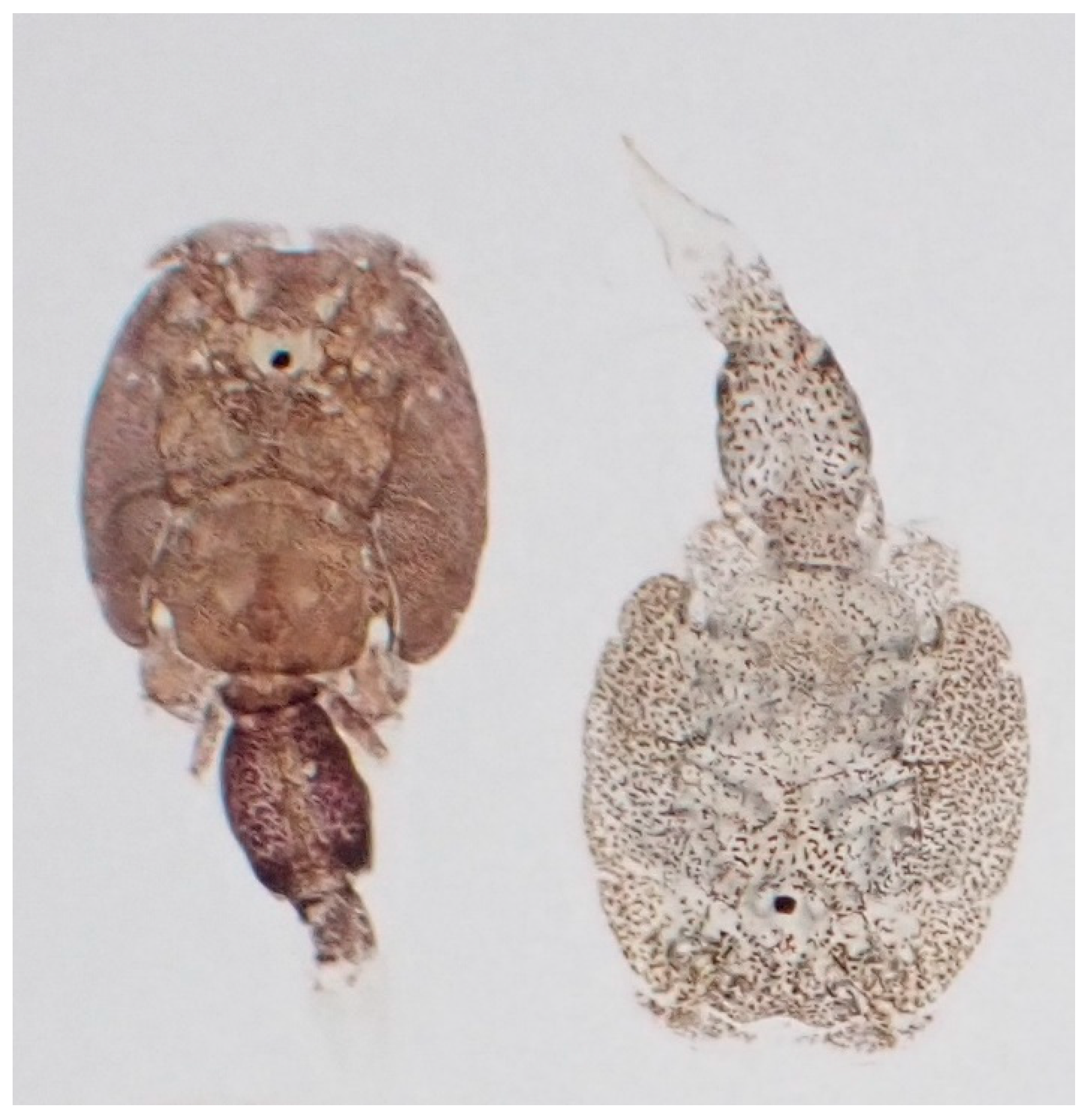
2.2. Photography and Measurement of Pigmentation
2.3. Statistical Analyses
3. Results
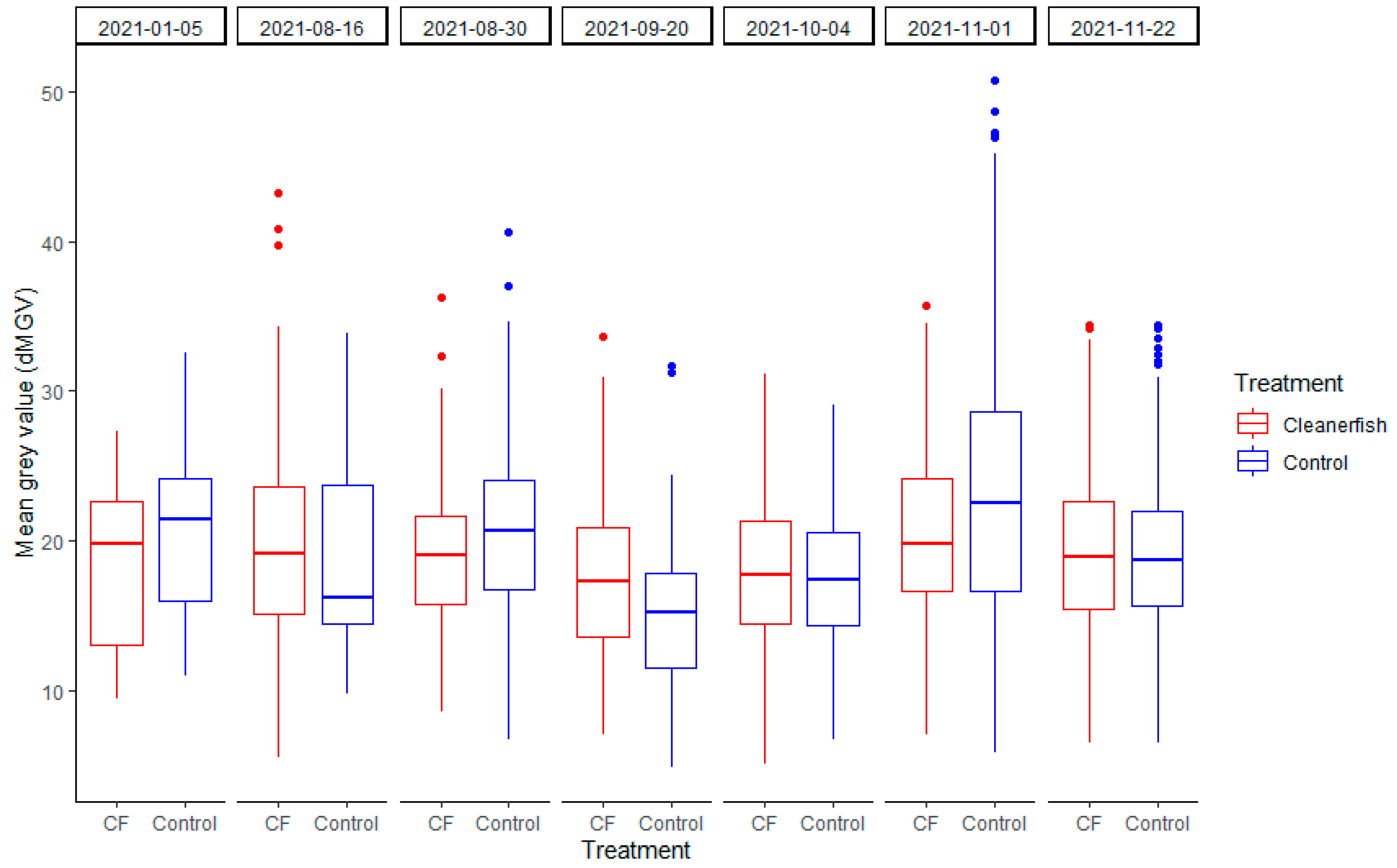
3.1. dMGV by Treatment
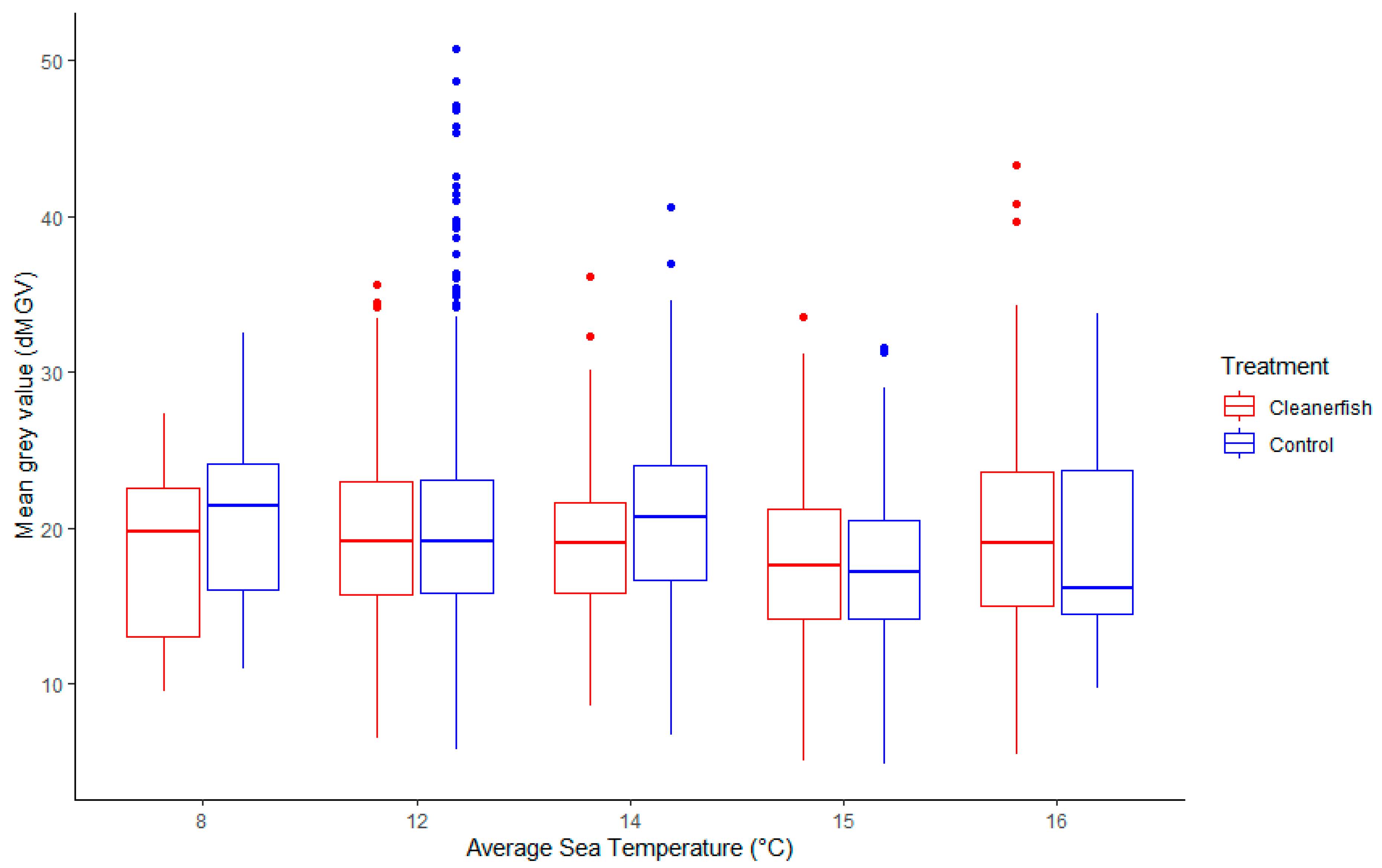
3.2. Seasonal Changes of dMGV
3.2.1. Sea Temperature
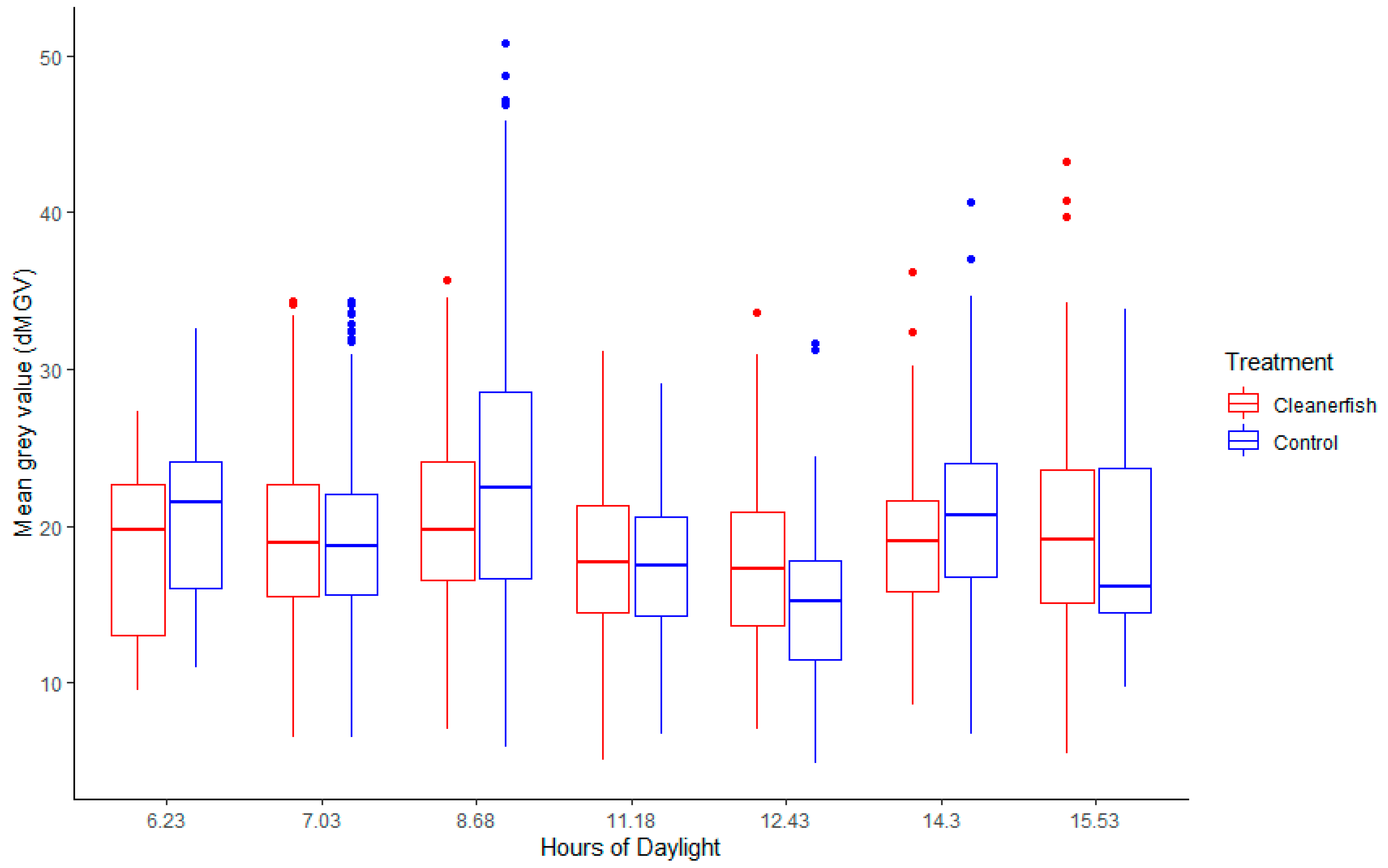
3.2.2. Hours of Daylight
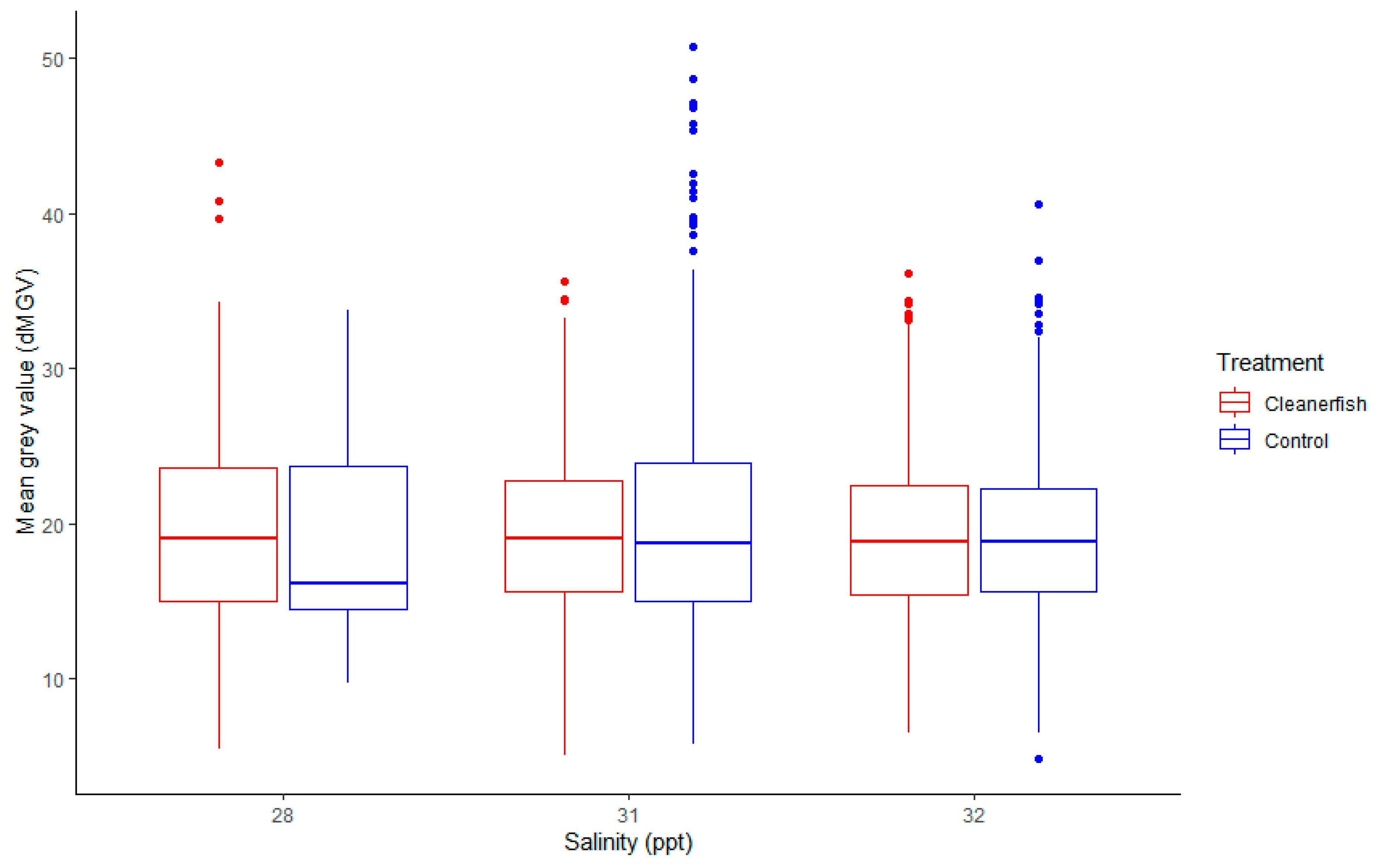
3.2.3. Salinity
3.3. dMGV by Stage and Sex
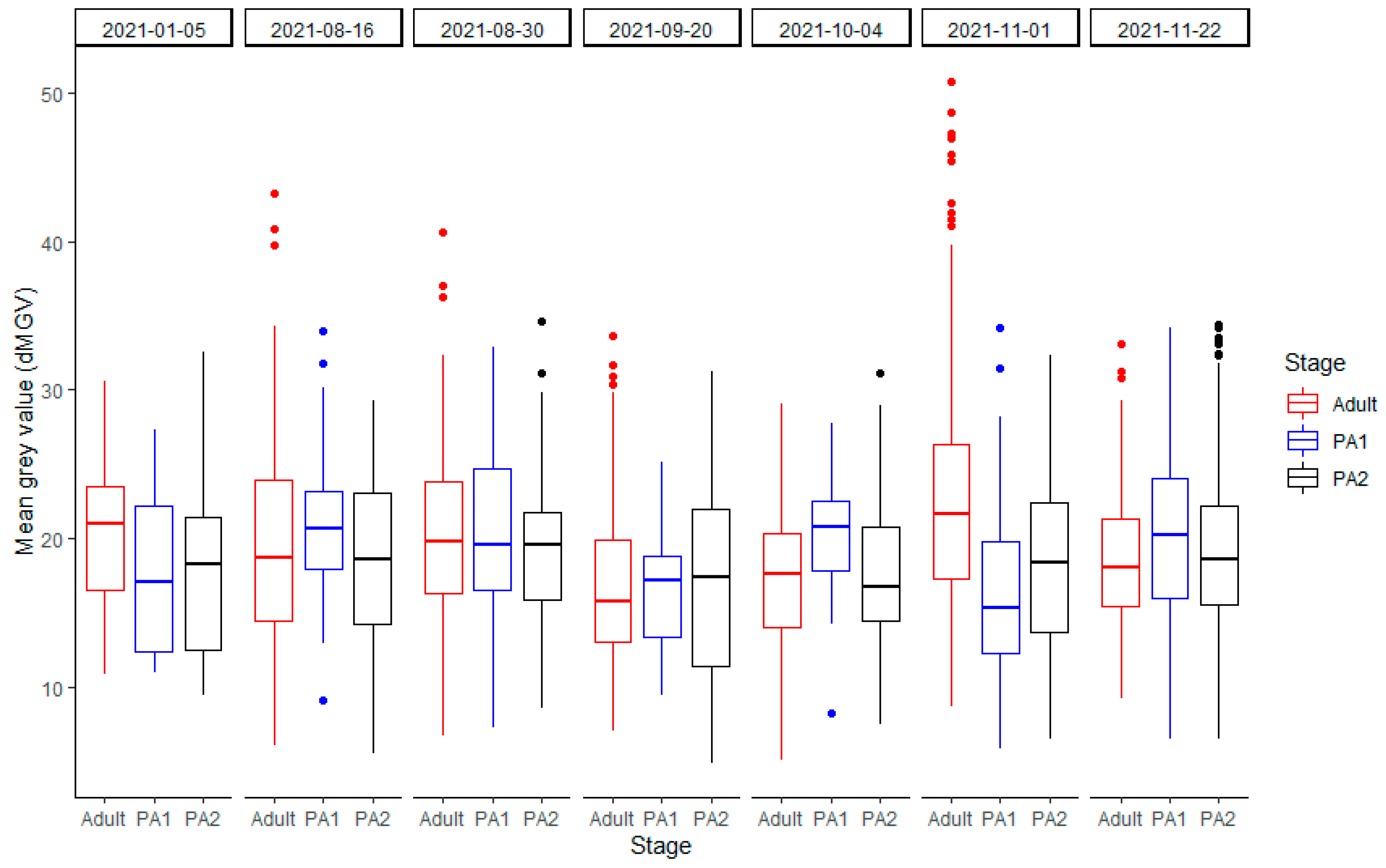
3.3.1. dMGV by Stage
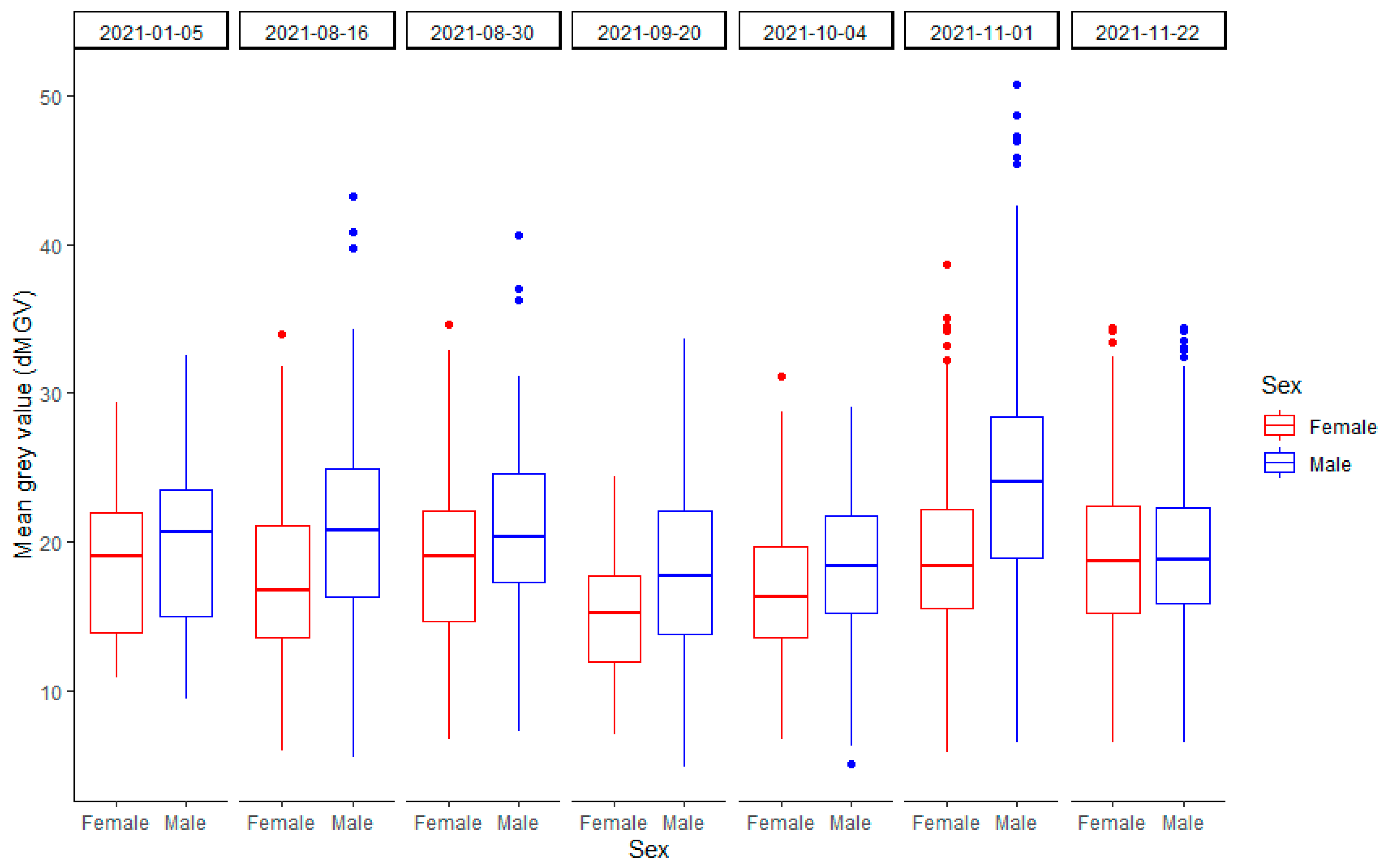
3.3.2. dMGV by Sex
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bui, S.; Oppedal, F.; Nola, V.; Barrett, L.T. Where art thou louse? A snapshot of attachment location preferences in salmon lice on Atlantic salmon hosts in sea cages. J. Fish Dis. 2020, 43, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Torrissen, O.; Jones, S.; Asche, F.; Guttormsen, A.; Skilbrei, O.T.; Nilsen, F.; Horsberg, T.E.; Jackson, D. Salmon lice—Impact on wild salmonids and salmon aquaculture. J. Fish Dis. 2013, 36, 171–194. [Google Scholar] [CrossRef]
- Wootten, R.; Smith, J.W.; Needham, E.A. Aspects of the biology of the parasitic copepods Lepeophtheirus salmonis and Caligus elongatus on farmed salmonids, and their treatment. Proc. Royal Soc. Edinb. 1982, 81, 185–197. [Google Scholar] [CrossRef]
- McEwan, G.F.; Groner, M.L.; Fast, M.D.; Gettinby, G.; Revie, C.W. Using agent-based modelling to predict the role of wild refugia in the evolution of resistance of sea lice to chemotherapeutants. PLoS ONE 2015, 10, e0139128. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.C.; Albright, L.J. The developmental stages of Lepeophtheirus salmonis (Krøyer, 1837) (Copepoda: Caligidae). Can. J. Zool. 1991, 69, 929–950. [Google Scholar] [CrossRef]
- Ljungfeldt, L.E.R.; Quintela, M.; Besnier, F.; Nilsen, F.; Glover, K.A. A pedigree-based experiment reveals variation in salinity and thermal tolerance in the salmon louse, Lepeophtheirus salmonis. Evol. Appl. 2017, 10, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Dempster, T.; Overton, K.; Bui, S.; Stien, L.H.; Oppedal, F.; Karlsen, Ø.; Coates, A.; Phillips, B.L.; Barrett, L.T. Farmed salmonids drive the abundance, ecology and evolution of parasitic salmon lice in Norway. Aquac. Environ. Interact. 2021, 13, 237–248. [Google Scholar] [CrossRef]
- Hamre, L.A.; Oldham, T.; Oppedal, F.; Nilsen, F.; Glover, K.A. The potential for cleaner fish-driven evolution in the salmon louse Lepeophtheirus salmonis: Genetic or environmental control of pigmentation? Ecol. Evol. 2021, 11, 7865–7878. [Google Scholar] [CrossRef]
- Jensen, E.M.; Horsberg, T.E.; Sevatdal, S.; Helgesen, K.O. Trends in de-lousing of Norwegian farmed salmon from 2000–2019—Consumption of medicines, salmon louse resistance and non-medicinal control methods. PLoS ONE 2020, 15, e0240894. [Google Scholar] [CrossRef]
- Fjørtoft, H.B.; Nilsen, F.; Besnier, F.; Espedal, P.G.; Stene, A.; Tveten, A.K.; Bjørn, P.A.; Aspehaug, V.T.; Glover, K.A. Aquaculture-driven evolution: Distribution of pyrethroid resistance in the salmon louse throughout the North Atlantic in the years 2000–2017. ICES J. Mar. Sci. 2020, 77, 1806–1815. [Google Scholar] [CrossRef]
- Besnier, F.; Kent, M.; Skern-Mauritzen, R.; Lien, S.; Malde, K.; Edvardsen, R.B.; Taylor, S.; Ljungfeldt, L.E.R.; Nilsen, F.; Glover, K.A. Human-induced evolution caught in action: SNP-array reveals rapid amphi-atlantic spread of pesticide resistance in the salmon ecotoparasite Lepeophtheirus salmonis. BMC Genom. 2014, 15, 937. [Google Scholar] [CrossRef] [PubMed]
- Ljungfeldt, L.E.R.; Espedal, P.G.; Nilsen, F.; Skern-Mauritzen, M.; Glover, K.A. A common-garden experiment to quantify evolutionary processes in copepods: The case of emamectin benzoate resistance in the parasitic sea louse Lepeophtheirus salmonis. BMC Evol. Biol. 2014, 14, 108. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Foss, A.; Vikingstad, E.; Elvegård, T.A. The use of lumpfish (Cyclopterus lumpus L.) to control sea lice (Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon (Salmo salar L.). Aquaculture 2014, 424–425, 18–23. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Notes on the behaviour of lumpfish with and without Atlantic salmon present. J. Ethol. 2014, 32, 117–122. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Nytrø, A.V.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Assessment of growth and sea lice infection levels in Atlantic salmon stocked in small-scale cages with lumpfish. Aquaculture 2014, 433, 137–142. [Google Scholar] [CrossRef]
- Imsland, A.K.; Hanssen, A.; Reynolds, P.; Nytrø, A.V.; Jonassen, T.M.; Hangstad, T.A.; Elvegård, T.A.; Urskog, T.C.; Mikalsen, B. It works! Lumpfish can significantly lower sea lice infections in large scale salmon farming. Biol. Open 2018, 7, bio036301. [Google Scholar] [CrossRef] [PubMed]
- Overton, K.; Dempster, T.; Oppedal, F.; Kristiansen, T.S.; Gismervik, K.; Stien, L.H. Salmon lice treatments and salmon mortality in Norwegian aquaculture: A review. Rev. Aquac. 2019, 11, 1398–1417. [Google Scholar] [CrossRef]
- Imsland, A.K.D.; Reynolds, P. In lumpfish we trust? The efficacy of lumpfish to control Lepeophtheirus salmonis infestations on farmed Atlantic salmon: A review. Fishes 2022, 7, 220. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Nytrø, A.V.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Feeding preferences of lumpfish (Cyclopterus lumpus L.) maintained in open net-pens with Atlantic salmon (Salmo salar L.). Aquaculture 2015, 436, 47–51. [Google Scholar] [CrossRef]
- Vaughan, D.B.; Grutter, A.S.; Costello, M.J.; Hutson, K.S. Cleaner fishes and shrimp diversity and a re-evaluation of cleaning symbioses. Fish Fish. 2017, 18, 698–716. [Google Scholar] [CrossRef]
- Overton, K.; Dempster, T.; Oppedal, F.; Kristiansen, T.S. Sea lice removal by cleaner fish in salmon aquaculture: A review of the evidence base. Aquac. Environ. Interact. 2020, 12, 31–44. [Google Scholar] [CrossRef]
- Thompson, C.R.S.; Madaro, A.; Nilsson, J.; Stien, L.H.; Oppedal, F.; Øverli, Ø.; Korzan, W.J.; Bui, S. Comparison of non-medicinal delousing strategies for parasite (Lepeophtheirus salmonis) removal efficacy and welfare impact on Atlantic salmon (Salmo salar) hosts. Aquac. Int. 2023. [Google Scholar] [CrossRef]
- Philis, G.; Ziegler, F.; Jansen, M.D.; Gansel, L.C.; Hornborg, S.; Aas, G.H.; Stene, A. Quantifying environmental impacts of cleaner fish used as sea lice treatments in salmon aquaculture with life cycle assessment. J. Ind. Ecol. 2022, 26, 1992–2005. [Google Scholar] [CrossRef]
- Nytrø, A.V.; Vikingstad, E.; Foss, A.; Hangstad, T.A.; Reynolds, P.; Eliassen, G.; Elvegård, T.A.; Falk-Petersen, I.B.; Imsland, A.K. The effect of temperature and fish size on growth of juvenile lumpfish (Cyclopterus lumpus L.). Aquaculture 2014, 434, 296–302. [Google Scholar] [CrossRef]
- Brooker, A.J.; Papadopoulou, A.; Gutierrez, C.; Rey, S.; Davie, A.; Migaud, H. Sustainable production and use of cleaner fish for the biological control of sea lice: Recent advances and current challenges. Vet. Rec. 2018, 183, 383. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.C. Observations on the distribution of lumpsucker (Cyclopterus lumpus, L.) in the Norwegian Sea. Fish. Res. 1993, 17, 369–372. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Mortensen, A.; Hansen, Ø.J.; Puvanendran, V.; Hangstad, T.A.; Jónsdóttir, Ó.D.B.; Emaus, P.A.; Elvegård, T.A.; et al. Is cleaning behavior in lumpfish (Cycloptherus lumpus) parentally controlled? Aquaculture 2016, 459, 156–165. [Google Scholar] [CrossRef]
- Hansson, L.A.; Hylander, S.; Sommaruga, R. Escape from UV threats in zooplankton: A cocktail of behavior and protective pigmentation. Ecology 2007, 88, 1932–1939. [Google Scholar] [CrossRef]
- Oren, A.; Gunde-Cimerman, N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol. Lett. 2007, 269, 1–10. [Google Scholar] [CrossRef]
- Scoville, A.G.; Pfrender, M.E. Phenotypic plasticity facilitates recurrent rapid adaptation to introduced predators. Proc. Natl. Acad. Sci. USA 2007, 107, 4260–4263. [Google Scholar] [CrossRef]
- Imsland, A.K.D.; Reynolds, P.; Jonassen, T.M.; Hangstad, T.A.; Elvegård, T.A.; Urskog, T.C.; Hanssen, A.; Mikalsen, B. Effects of different feeding frequencies on growth, cataract development and histopathology of lumpfish (Cyclopterus lumpus L.). Aquaculture 2019, 501, 161–168. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Nytrø, A.V.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Assessment of suitable substrates for lumpfish in sea pens. Aquac. Int. 2015, 23, 639–645. [Google Scholar] [CrossRef]
- Imsland, A.K.D.; Conlon, H. Money for nothing: The possible use of recycled fish farm material as habitat for lumpfish. Aquac. Eng. 2019, 87, 102015. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Berg-Hansen, L.; Hangstad, T.A.; Elvegård, T.A.; Urskog, T.C.; Mikalsen, B. Assessment of artificial substrates for lumpfish: Effect of material thickness and water current speed. Aquac. Int. 2018, 26, 1469–1479. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. (v4.1.2). 2021. Available online: https://www.r-project.org/ (accessed on 10 April 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 10 April 2022).
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1984; 718p. [Google Scholar]
- Staven, F.R.; Nordeide, J.T.; Gesto, M.; Andersen, P.; Patel, D.M.; Kristensen, T. Behavioural and physiological responses of lumpfish (Cyclopterus lumpus) exposed to Atlantic salmon (Salmo salar) sensory cues. Aquaculture 2021, 544, 737066. [Google Scholar] [CrossRef]
- Brooker, A.J.; Skern-Mauritzen, R.; Bron, J.E. Production, mortality, and infectivity of planktonic larval sea lice, Lepeophtheirus salmonis (Krøyer, 1837): Current knowledge and implications for epidemiological modelling. ICES J. Mar. Sci. 2018, 75, 1214–1234. [Google Scholar] [CrossRef]
- Hvas, M.; Folkedal, O.; Oppedal, F. Fish welfare in offshore salmon aquaculture. Rev. Aquac. 2021, 13, 836–852. [Google Scholar] [CrossRef]
- Geitung, L.; Oppedal, F.; Stien, L.H.; Dempster, T.; Karlsbakk, E.; Nola, V.; Wright, D.W. Snorkel sea-cage technology decreases salmon louse infestation by 75% in a full-cycle commercial test. Int. J. Parasitol. 2019, 49, 843–846. [Google Scholar] [CrossRef]
- Geitung, L.; Wright, D.W.; Oppedal, F.; Stien, L.H.; Vågseth, T.; Madaro, A. Cleaner fish growth, welfare and survival in Atlantic salmon sea cages during an autumn-winter production. Aquaculture 2020, 528, 735623. [Google Scholar] [CrossRef]
- Leclercq, E.; Zerafa, B.; Brooker, A.J.; Davie, A.; Migaud, H. Application of passive-acoustic telemetry to explore the behaviour of ballan wrasse (Labrus bergylta) and lumpfish (Cyclopterus lumpus) in commercial Scottish salmon sea-pens. Aquaculture 2018, 495, 1–12. [Google Scholar] [CrossRef]
- Gentry, K.; Bui, S.; Oppedal, F.; Dempster, T. Sea lice prevention strategies affect cleaner fish delousing efficacy in commercial Atlantic salmon sea cages. Aquac. Environ. Interact. 2020, 12, 67–80. [Google Scholar] [CrossRef]
- Hamre, L.A.; Bui, S.; Oppedal, F.; Skern-Mauritzen, R.; Dalvin, S. Development of the salmon louse Lepeophtheirus salmonis parasitic stages in temperatures ranging from 3 to 24 °C. Aquac. Environ. Interact. 2019, 11, 429–443. [Google Scholar] [CrossRef]
- Samsing, F.; Oppedal, F.; Dalvin, S.; Johnsen, I.; Vågseth, T.; Dempster, T. Salmon lice (Lepeophtheirus salmonis) development times, body size, and reproductive outputs follow universal models of temperature dependence. Can. J. Fish. Aquat. Sci. 2016, 73, 1841–1851. [Google Scholar] [CrossRef]
- Coates, A.; Phillips, B.L.; Bui, S.; Oppedal, F.; Robinson, N.A.; Dempster, T. Evolution of salmon lice in response to management strategies: A review. Rev. Aquac. 2021, 13, 1397–1422. [Google Scholar] [CrossRef]
- Handeland, S.O.; Imsland, A.K.; Stefansson, S.O. The effect of temperature and fish size on growth, feed intake, food conversion efficiency and stomach evacuation rate of Atlantic salmon post-smolts. Aquaculture 2008, 283, 36–42. [Google Scholar] [CrossRef]
- Oppedal, F.; Juell, J.E.; Taranger, G.L.; Hansen, T. Artificial light and season affects vertical distribution and swimming behaviour of post-smolt Atlantic salmon in sea cages. J. Fish Biol. 2001, 58, 1570–1584. [Google Scholar] [CrossRef]
- Hamre, L.A.; Glover, K.A.; Nilsen, F. Establishment and characterisation of salmon louse (Lepeophtheirus salmonis (Krøyer 1837)) laboratory strains. Paras Int. 2009, 58, 451–460. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lumpfish | |||
|---|---|---|---|
| Cage 1 | Cage 4 | Cage 5 | |
| Week 46 2020 | 10697 | 10521 | 10470 |
| Week 8 2021 | 9792 | 9805 | 9817 |
| Ballan wrasse | |||
| Week 21 2021 | 10,654 | 9876 | 9935 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imsland, A.K.D.; Berg, J.P.; Nola, V.; Geitung, L.; Oldham, T. Cleaner Fish Do Not Impact the Pigmentation of Salmon Lice (Lepeophtheirus salmonis) in Commercial Aquaculture Cages. Fishes 2023, 8, 455. https://doi.org/10.3390/fishes8090455
Imsland AKD, Berg JP, Nola V, Geitung L, Oldham T. Cleaner Fish Do Not Impact the Pigmentation of Salmon Lice (Lepeophtheirus salmonis) in Commercial Aquaculture Cages. Fishes. 2023; 8(9):455. https://doi.org/10.3390/fishes8090455
Chicago/Turabian StyleImsland, Albert Kjartan Dagbjartarson, Jon Petter Berg, Velimir Nola, Lena Geitung, and Tina Oldham. 2023. "Cleaner Fish Do Not Impact the Pigmentation of Salmon Lice (Lepeophtheirus salmonis) in Commercial Aquaculture Cages" Fishes 8, no. 9: 455. https://doi.org/10.3390/fishes8090455
APA StyleImsland, A. K. D., Berg, J. P., Nola, V., Geitung, L., & Oldham, T. (2023). Cleaner Fish Do Not Impact the Pigmentation of Salmon Lice (Lepeophtheirus salmonis) in Commercial Aquaculture Cages. Fishes, 8(9), 455. https://doi.org/10.3390/fishes8090455