1. Introduction
Many aquaculture systems harness natural biological productivity to support the production of edible aquatic organisms. The extractive species inside the culture ponds can convert particulate and dissolved materials not ingestible by animals of higher trophic levels into biomass, effectively recovering inaccessible energy and nutrients. This process should be explored to restore natural resources from waste and pollution inside ponds, according to a circular economy approach [
1]. Nevertheless, nutrient limitation may hinder biological productivity in aquaculture ponds. To compensate, organic and inorganic fertilization may be performed to stimulate natural productivity [
2,
3]. Fertilization is frequently used in ponds recently stocked with fingerlings or during early grow-out cultures to boost the development of natural food because most fishes have limitations in ingesting or digesting manufactured feed in the early life stages [
4]. During the grow-out phase, fertilization has been observed to enhance fish productivity in various species, including
Cirrhinus migrala [
5], different carps in polyculture [
6,
7,
8],
Coptodon rendalli [
9], and
Labeo rohita [
10]. This effect is generally attributed to a rise in natural food, mainly phytoplankton, zooplankton, periphyton, and benthic invertebrates.
Most studies have evaluated the effect of fertilization on pond productivity without the supply of formulated feed. This practice results in limited productivity because the enhancement of natural food is insufficient to feed fish stocked in high densities. Productivity is low in fertilized ponds without an allochthonous diet, while intensively fed ponds bear high productivity. An allochthonous diet is the primary source of nutrients in semi-intensive or intensive fish monocultures [
3,
11]. Despite this, natural food can supplement the formulated feed in ponds or partially replace it [
4,
12]. They are even essential in farming certain species for which no suitable and well-balanced diets are available [
13,
14]. Therefore, understanding the role of fertilization associated with an allochthonous diet may provide information for aquaculture efficiency and align with restoration approaches (sensus Alleway [
1]).
The incorporation of ingested food into fish biomass and its contribution to growth is variable. The intake of allochthonous diets or natural biota organisms has been largely studied in fish production by stomach content analysis [
10,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25]. However, this technique does not reveal the contribution of each kind of feed to fish growth because part of the eaten feed is not assimilated. Stable isotope analysis is a tool for quantifying dietary support of fish growth among the ingested feed items with distinct isotope content, since the isotopic compositions of animal tissues resemble their diets [
26,
27,
28,
29,
30,
31]. Since formulated diet is the main cost of production in fish farms [
3], understanding its utilization by fish and how it can be replaced or complemented by natural food would help to improve feeding strategies and decrease production costs.
The tambaqui (
Colossoma macropomum) is an indigenous fish species from the Amazon River Basin and has been framed in Central and South American countries, including Panama, Honduras, Brazil, Colombia, Peru, and Venezuela. This species has also been farmed in China [
32]. This Tambaqui is a low-trophic-level fish that exhibits a remarkable ability to consume natural food sources both in its natural habitat [
33,
34] and in rearing ponds during all life stages [
20,
22]. Tambaqui also shows easy captivity adaptation, a low feed conversion ratio (FCR), and fast growth [
12,
22]. Brazil is the primary producer [
35], where tambaqui is farmed in earthen ponds [
36]. The farming practices include a direct stocking of juveniles weighing between 2 and 5 g or a preliminary culture phase lasting 2 to 3 months until the fish reach a weight of 70 to 80 g, and then, they are transferred to grow-out ponds where they reach 1.5 to 3 kg within a span of 10 months [
12,
37].
Tambaqui culture in ponds may be an interesting model for studying the contribution of natural food in the grow-out phase to low-trophic-level fish growth. Combining the determination of stomach content and stable isotopic analyses enables us to determine both the ingested food by fish and its actual assimilation into their body mass. Therefore, the present study aims to describe the intake of natural food and its contribution to tambaqui growth in fertilized and unfertilized ponds. Furthermore, it aims to understand the impact of fertilization on natural food availability, fish performance, and production costs.
4. Discussion
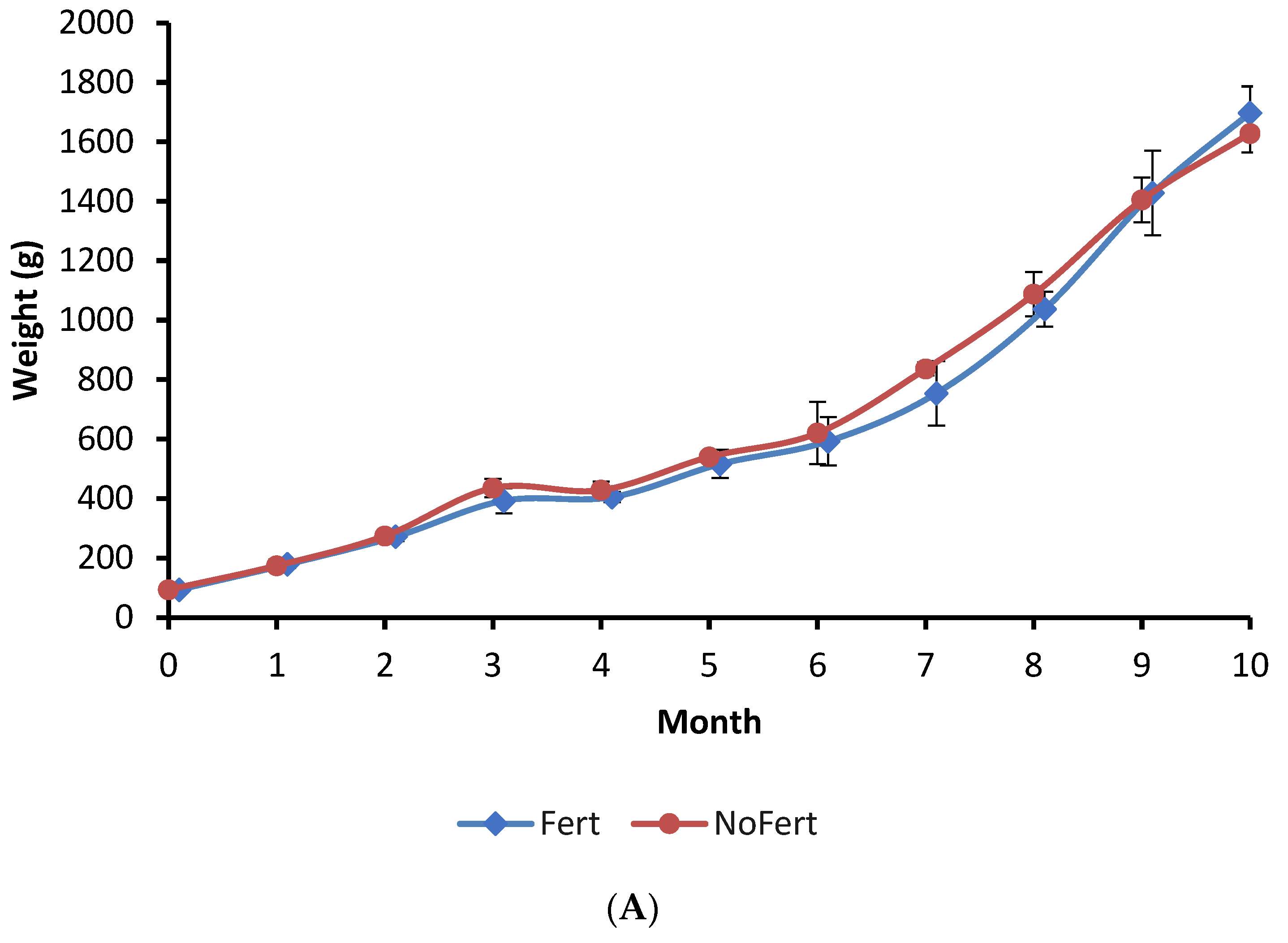
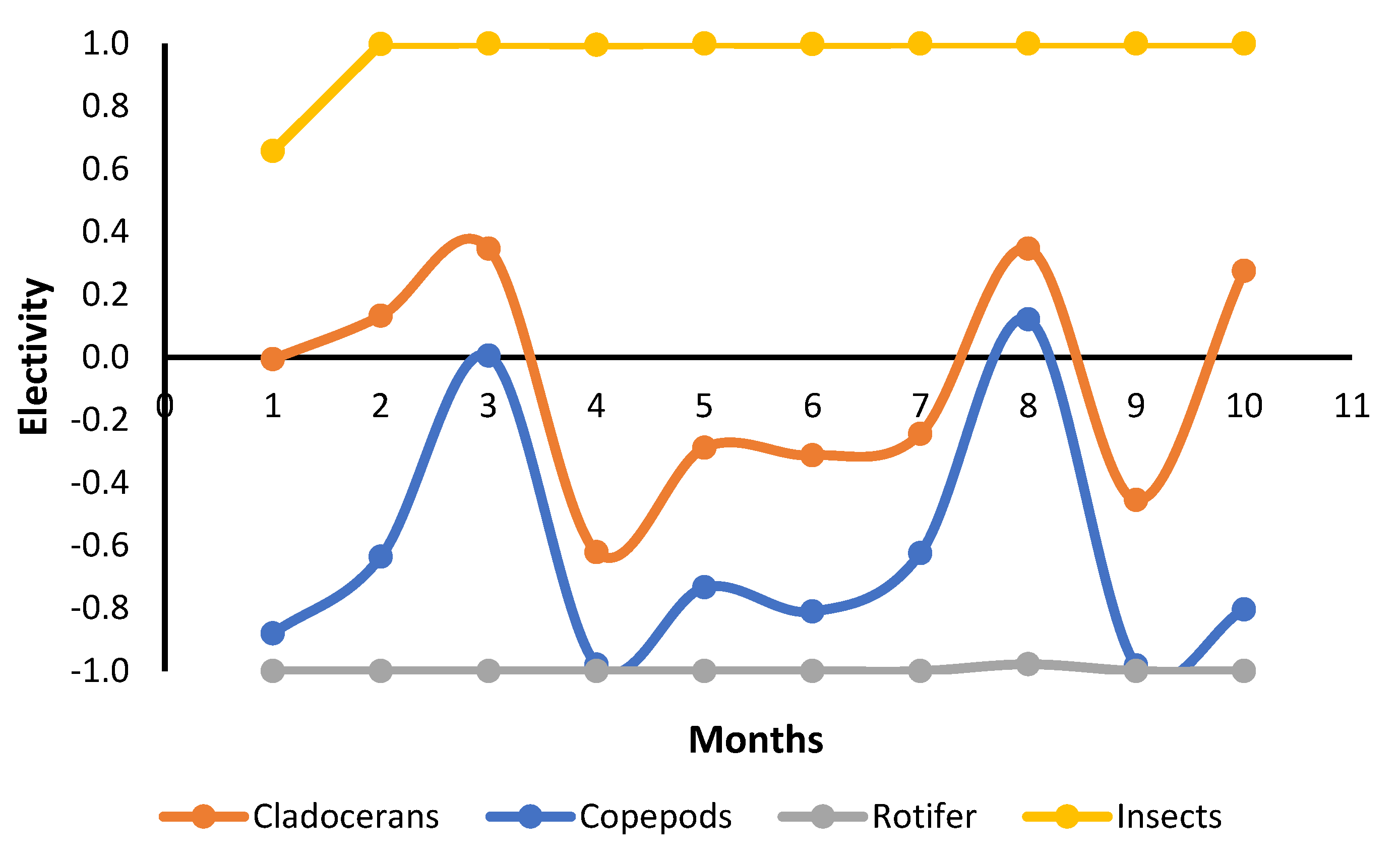
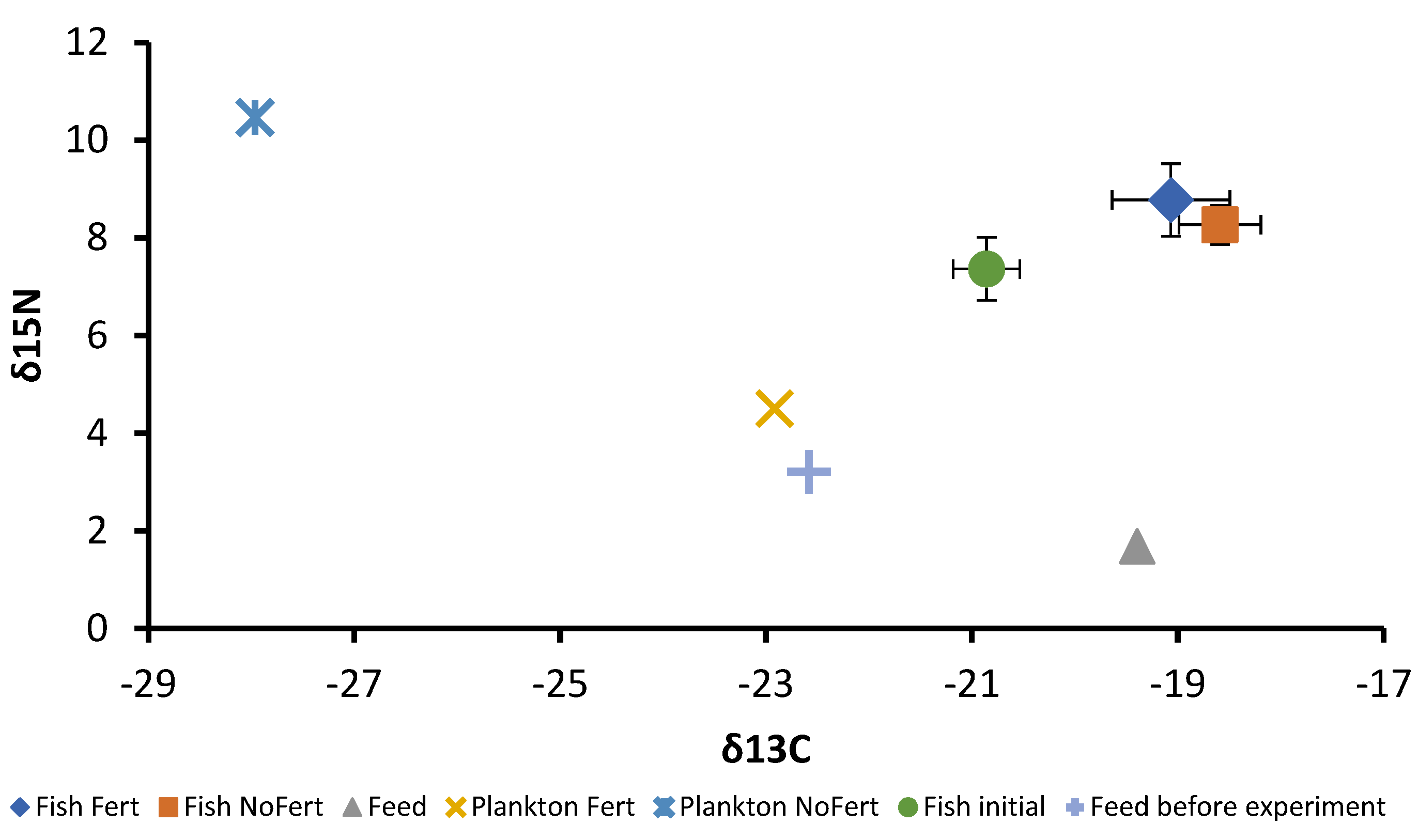
Tambaqui consumed zooplankton, fragments of vegetables, and formulated feed during the entire grow-out phase in both fertilized and unfertilized ponds. Results indicated that pond fertilization increased zooplankton density in the pond water, through phytoplankton, thereby enhancing natural food availability. This allowed a greater exploration of natural biota as food, characterized by a selective increase in zooplankton ingestion by the tambaqui. Consequently, zooplankton accounted for 39.4% of all assimilated food, whereas it constituted only 10.7% in unfertilized ponds. On the other hand, this greater assimilation of natural food did not lead to higher growth or productivity. This suggests that this species can interchange between natural food and a commercial diet without compromising its development.
The limited availability of natural food may have constrained the contribution of this food to muscle tissue growth, as evidenced by zooplankton density in unfertilized ponds. The lack of natural food was compensated by the increase in formulated feed ingestion, showed by a higher feed conversion rate (computed only using the feed supplied) of tambaqui in unfertilized ponds. A high contribution of natural food to fish growth was also observed in juvenile channel catfish (
Ictalurus punctatus) reared in nursery ponds receiving eutrophic water [
53]. In this culture, natural food accounted for 57–58% of muscle tissue growth when fish were fed commercial feeds at varying rates, suggesting that the availability of formulated feed did not determine the consumption of natural food, and rather, it was primarily influenced by its own abundance [
53].
The stable isotope analysis has been utilized in food assimilation studies to quantify the contribution of natural food to fish growth in farm ponds [
27,
28,
29,
30,
54]. However, this is the first study of tambaqui in ponds, despite the importance of this species for aquaculture. As previously observed for other species, alterations in the isotopic signal were detected in both treatments due to intake and nutrient assimilation from natural food and commercial feed [
53,
54,
55]. Fish reared in unfertilized ponds exhibited higher δ
13C enrichment, indicating greater formulated feed assimilation as confirmed by food-type assimilation analysis and lower cladocerans ingestion resulting in a higher FCR. On the other hand, zooplankton impacts more intensively in δ
14N enrichment [
56], which explains the higher δ
14N enrichment in tambaqui produced in fertilized ponds, corroborating the higher contribution of natural food in this pond management. The high assimilation of plankton by tambaqui demonstrates that these low-trophic fish may play an important role in recovering energy and nutrients in pond systems even when receiving allochthonous feed. This characteristic places the species as a promising candidate for restorative aquaculture in fed or unfed cultures in which aquaculture provides ecological benefits to the environment leading to improved environmental sustainability and ecosystem services, in addition to the supply of food [
1].
Interestingly, differences in plankton isotopic signals from fertilized and unfertilized ponds were observed, which can be related to fertilization management. Narimbi et al. [
30] observed that an inorganic nitrogen source was the main isotopic signal responsible for phytoplankton in ponds of tilapia culture even when organic fertilizer was available. Similarly, in the present study, lower δ
14N values were observed in plankton from fertilized ponds. This observation can be explained by the utilization of urea as a nitrogen source, which presents a δ
14N signal close to zero [
57].
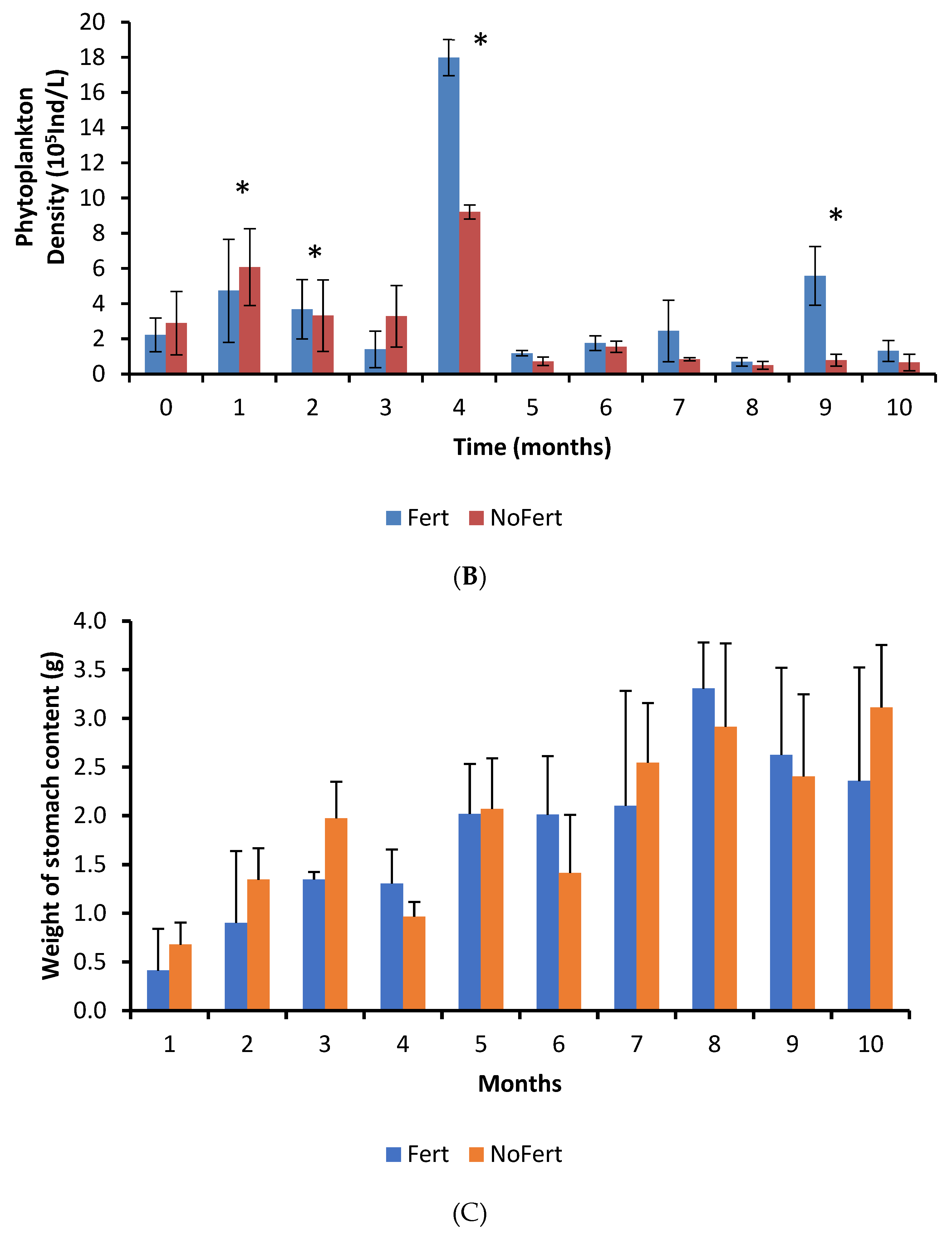
The density of phytoplankton and concentration of TAN in the pond water did not differ between fertilized and unfertilized ponds during most of the culture period. Probably, in fertilized ponds, phytoplankton rapidly assimilated the added nitrogen and phosphorous, and the populations were regulated by the zooplankton, which increased with the cascade effect. Zooplankton supports the nutrient flow between phytoplankton and fish, contributing to restoring nutrients and energy in the pond system [
58]. However, Brucet et al. [
59] and Vakkilainen et al. [
60] highlighted a more significant influence of fish predation than nutrient enrichment (via phytoplankton) in regulating zooplankton populations.
We observed a high presence of insects and terrestrial plants in tambaqui stomachs. The vegetables were seeds and leaf fragments, possibly from the vegetation that surrounded ponds. Conversely, insects and terrestrial plants have low importance in tambaqui food in Amazonian rivers [
33,
56]. The differences may be related to the availability of these food items in the water or a change in the food preference of tambaqui in culture ponds. In the present study, isotopic analyses neglected vegetables and insects because we chose the principal items observed in the natural field. Although insects were present in the zooplankton sample, it may not have covered all the diversity and abundance of insects present in the ponds. Thus, the contribution of these natural food items to tambaqui muscle tissue growth was subestimated.
The density of cladocerans, copepods, and rotifers was quite variable during the culture; however, on average, it was approximately double in fertilized ponds. Nevertheless, only cladocerans were more abundant in the stomachs of fish raised in fertilized ponds, indicating a feed preference for this group. These results may be due to the capacity to catch prey or selectivity pre- or post-capture. Larvae of tambaqui showed substantial ingestion of rotifers [
61], while juveniles [
62] and adults (present results) showed low consumption of it. Thus, it is likely that a gill raker limitation to capture this small prey is developed with the growth of tambaqui. On the other hand, copepods have similar or higher sizes than cladocerans, and thus lower consumption of them indicates an avoidance and a real preference for cladorerans. Therefore, further research should be conducted on fertilization management to boost cladocerans and insect populations. An additional benefit of increased zooplankton density in fishponds is the action of this community to improve the effectiveness of the biological carbon pump that regulates the atmospheric carbon dioxide levels, which is an ecosystem service of aquaculture [
58,
63]. Techniques promoting the intensification of natural food such as fertilization, the addition of substrates for growing periphyton, and bottom nutrients suspension may contribute to reducing tambaqui’s reliance on formulated feed and enable ecological intensification of production through ecological processes and functions [
64].
The natural biota plays a crucial role in nutrient cycling within aquaculture ponds, contributing to biogeochemical cycles and solar energy fixation. These fundamental processes form the basis for a restorative aquaculture practice [
1,
58]. The observed 8% decrease in the feed conversion ratio (FCR) in fertilized ponds indicates that the natural biota can partially substitute commercial diet in tambaqui production without compromising productivity, thereby reducing reliance on formulated feed. Therefore, an increase in natural food is desirable in this production system. Isotopic analyses showed that about 40% of tambaqui biomass comes from the aquatic biota. Therefore, tambaqui may be a candidate for restorative aquaculture practices. This characteristic may bring a new perspective to the production of this Amazon species.
The lower weight of tambaqui stomach content in fish produced in fertilized ponds could be because of the fastest movement of natural food in the gut [
65] and its greater digestibility [
65,
66]. On the other hand, higher ingestion of formulated feed and its high participation in muscle building in unfertilized ponds justify the greater weight of stomach content in this treatment because it is less digestible. The stomachs were fixed in formalin solution until the analysis, and natural food may have had higher dehydration than formulated feed due to its humidity (~90%), increasing the difference observed [
67]. Formulated feed was observed inside tambaqui stomachs despite the feeding stopped a day before gut samples. This presence is due to the long time elapsed from ingestion to gastric evacuation. In tambaqui, the gut is empty after 24 h of the last feeding only in 50% of fish [
68]. In pacu,
Piaractus mesopotamicus, a species of the same family of tambaqui, gastric emptying can occur in 56 h, depending on water temperature [
69]. The ingestion rate of tambaqui decreased during the development, as it was demonstrated by the ratio of stomach weight and total fish weight.
In general, fertilization promotes an increase in natural food productivity in fishponds, which supplement formulated feed and result in higher fish growth and productivity [
4]. The positive effect of fertilization on fish production was described for many fish species, such as carps in monoculture and polyculture [
2,
5,
8,
70], red-breasted tilapia (
Coptodon rendalli, former
Tilapia rendalli) [
9], Nile tilapia (
Oreochromis niloticus), North African catfish (
Clarias gariepinus) [
49], and pirarucu (
Arapaima gigas) [
71]. However, in the present study, fertilization did not improve tambaqui productivity, similar to what was observed by Gomes and Silva [
22] in tambaqui and Duodu et al. [
72] in Nile tilapia (
Oreochromis niloticus) in combining feed with fertilization (N:P ratio of 1:1 and 1.6:1, respectively). In the present study, the commercial feed supplied in unfertilized ponds compensated for the lower natural food ingestion and did not affect fish growth. Thus, commercial feed available for tambaqui is probably well-balanced and allows fish to grow with a low dependence on natural food. As tambaqui is not a territorial or aggressive fish, it may be produced in intensive systems where natural food is negligible.
The water quality parameters meet those recommended for fish production by Boyd [
73] in both fertilized and unfertilized ponds. Therefore, they did not affect the growth and survival of tambaqui. The temperature was similar in ponds of both treatments and fluctuated according to weather variations over the study (
cf. INMET data [
74]). Transparency was lower, and phytoplankton density was higher in some months in fertilized ponds, corroborating the impact of fertilization procedure in ponds described by Boyd [
4]. The dissolved oxygen increased up to the fifth month, when it decreased, possibly because of an elevation in fish biomass that demands more oxygen. TAN decreased until the middle of the production cycle when it increased owing to an elevation in tambaqui biomass, as observed previously by Gomes and Silva [
22] and indicated by Boyd [
75]. Boyd [
73] reports that feed and fertilizers are the primary sources of TAN in aquaculture systems. The similarity among treatments suggests that fertilizers had a low impact on TAN levels or that the ammonia was rapidly assimilated by phytoplankton in fertilized ponds. However, phosphorous concentration was significantly higher in fertilized ponds, suggesting that the ratio N:P in fertilizer was unbalanced for fulfilling the phytoplankton requests. Alkalinity and hardness were similar in all ponds of both treatments, probably because they were equally limed. Generally, the results indicated that the fertilization of earthen ponds using 5 g of urea, 3 g of triple superphosphate, and 10 g of rice bran per square meter produces a low effect on water quality in tambaqui ponds.
Fertilization management increased production costs despite reduced feed use. The costs of fertilizer and labor to handle it surpassed the expenditure reduction in commercial feed by 8%. Duodu et al. [
72] also observed a slight increment in the production costs of Nile tilapia (
Oreochromis niloticus) in Ghana when fish were produced by combining inorganic fertilization (1.6:1, N:P ratio) with commercial feed. As inorganic fertilizers are expensive, organic fertilizers could be an alternative to decrease the costs [
76]. Further studies should be conducted to define the cost-effective composition and quantity supplied of fertilizers to lead to the best composition of zooplankton to feed tambaqui.
Generally, a large amount of nutrients is accumulated in fishpond bottoms [
77]. Thus, techniques to suspend the sediments, such as aeration, may contribute to making nutrients laid in sediment available to the phytoplankton community [
60]. The use of benthic species integrated with the culture of tambaqui may resuspend nutrients in the sediment by bioturbation with no cost [
77,
78,
79]. Therefore, the culture of tambaqui combined with benthic feeders or iliophagus species may be advantageous to recovering sequestered nutrients, saving fertilizers and commercial diet. This practice increases the circularity and restorative capacity of the production system. In addition, it increases the productivity and diversity of products, transforming pollution into biomass of high economic value. Further studies should be performed to determine the best species and stocking densities and ratios for improving productivity in integrated systems.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}