
Mechanism of Ligilactobacillus salivarius GX118 in Regulating the Growth of Rainbow Trout (Oncorhynchus mykiss) and Resistance to Aeromonas salmonicida Infection

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactic Acid Bacteria Strain and Cultivation Conditions
2.2. In Vitro Antibacterial Experiment
2.3. Whole-Genome Analysis of GX118
2.4. Rainbow Trout Feeding Experiment
2.5. Rainbow Trout Sampling
2.6. Growth and Survival of Rainbow Trout
2.7. 16S rRNA Sequencing Analysis
2.8. Gene Expression Pertaining to the Immune System
2.9. Freezing Section DAPI Staining Experiment
2.10. Statistical Analysis
3. Results
3.1. Bacteriostasis of GX118 In Vitro
3.2. Figures, Tables, and Schemes
3.3. Genome-Wide Analysis of L. salivarius GX118
3.4. GX118 Improved Growth Performance and Survival of Rainbow Trout
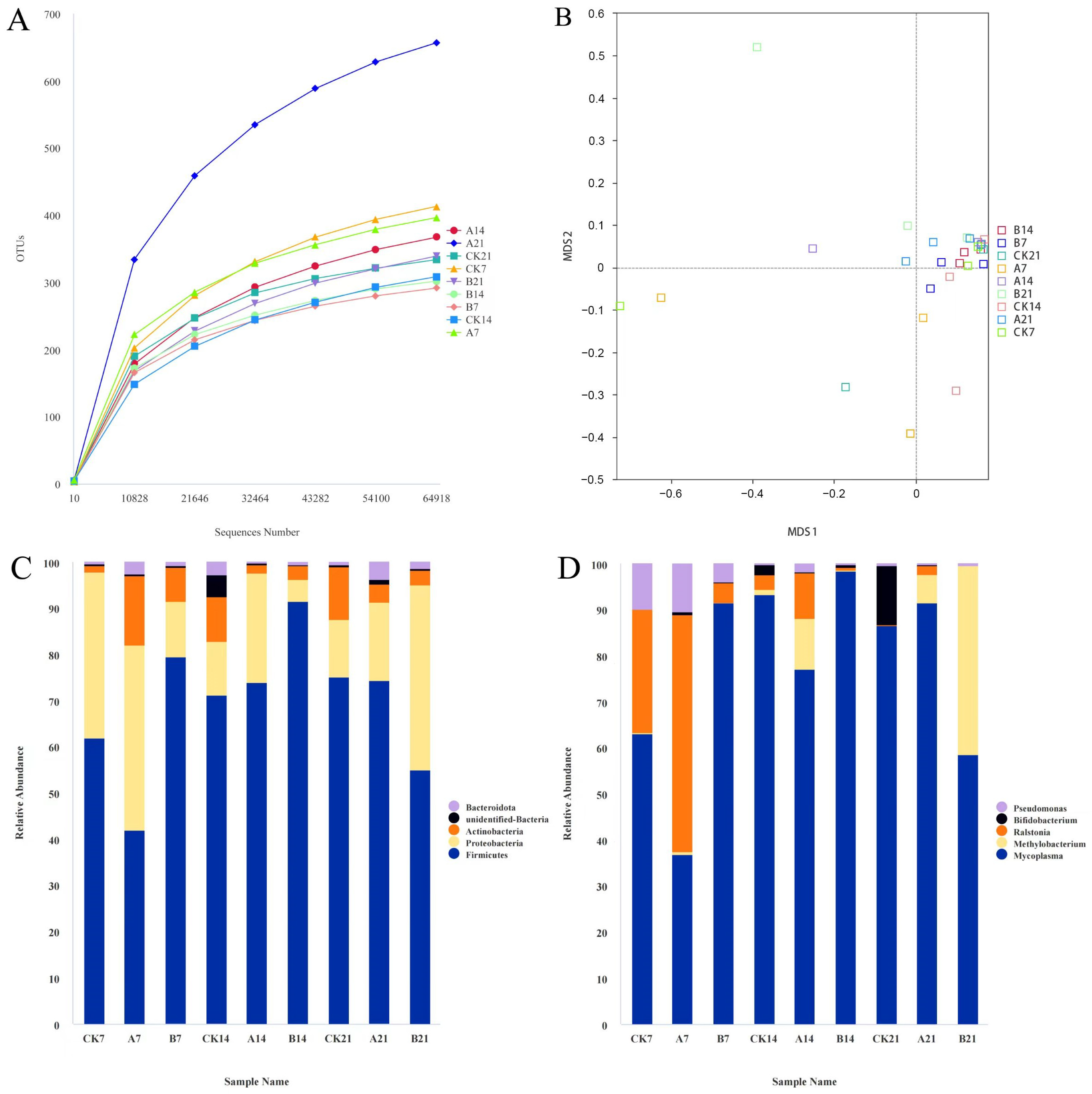
3.5. GX118 Affected Gut Microbiota Diversity and Composition
3.6. Expression of Immune-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, J.; Sun, R.; Pan, C.; Sun, Y.; Mai, B.; Li, Q.X. Antibiotics and Food Safety in Aquaculture. J. Agric. Food Chem. 2020, 68, 11908–11919. [Google Scholar] [CrossRef]
- Kakakhel, M.A.; Narwal, N.; Kataria, N.; Johari, S.A.; Zaheer Ud Din, S.; Jiang, Z.; Khoo, K.S.; Xiaotao, S. Deciphering the dysbiosis caused in the fish microbiota by emerging contaminants and its mitigation strategies—A review. Environ. Res. 2023, 237, 117002. [Google Scholar] [CrossRef]
- Zhao, C.; Men, X.; Dang, Y.; Zhou, Y.; Ren, Y. Probiotics Mediate Intestinal Microbiome and Microbiota-Derived Metabolites Regulating the Growth and Immunity of Rainbow Trout (Oncorhynchus mykiss). Microbiol. Spectr. 2023, 11, e0398022. [Google Scholar] [CrossRef]
- Amoah, K.; Tan, B.; Zhang, S.; Chi, S.; Yang, Q.; Liu, H.; Yang, Y.; Zhang, H.; Dong, X. Host gut-derived Bacillus probiotics supplementation improves growth performance, serum and liver immunity, gut health, and resistive capacity against Vibrio harveyi infection in hybrid grouper (female symbol Epinephelus fuscoguttatus × male symbol Epinephelus lanceolatus). Anim. Nutr. 2023, 14, 163–184. [Google Scholar] [CrossRef]
- Alonso, S.; Carmen Castro, M.; Berdasco, M.; de la Banda, I.G.; Moreno-Ventas, X.; de Rojas, A.H. Isolation and Partial Characterization of Lactic Acid Bacteria from the Gut Microbiota of Marine Fishes for Potential Application as Probiotics in Aquaculture. Probiotics Antimicrob. Proteins 2019, 11, 569–579. [Google Scholar] [CrossRef]
- Hashim, H. Bacteriocin: The avenues of innovation towards applied microbiology. Pure Appl. Biol. 2018, 7, 460–478. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial production of bacteriocins: Latest research development and applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef]
- Shan, C.; Wu, H.; Zhu, Y.; Zhou, J.; Yan, W.; Jianhao, Z.; Liu, X. Preservative effects of a novel bacteriocin from Lactobacillus panis C-M2 combined with dielectric barrier discharged cold plasma (DBD-CP) on acquatic foods. Food Sci. Technol. Int. 2023, 29, 406–416. [Google Scholar] [CrossRef]
- Selle, K.; Klaenhammer, T.R. Genomic and phenotypic evidence for probiotic influences of Lactobacillus gasseri on human health. FEMS Microbiol. Rev. 2013, 37, 915–935. [Google Scholar] [CrossRef]
- Daba, G.M.; Elkhateeb, W.A. Ribosomally synthesized bacteriocins of lactic acid bacteria: Simplicity yet having wide potentials—A review. Int. J. Biol. Macromol. 2024, 256, 128325. [Google Scholar] [CrossRef] [PubMed]
- Xiang, F.; Xu, G.; Gu, W.; Huang, T.; Liu, C.; Wang, B. Expression of cytochrome genes during the early gonadal development in triploid female rainbow trout Oncorhynchus mykiss. J. Fish. Sci. China 2018, 25, 949. [Google Scholar] [CrossRef]
- Duman, M.; Altun, S.; Saticioglu, I.B.; Romalde, J.L. A review of bacterial disease outbreaks in rainbow trout (Oncorhynchus mykiss) reported from 2010 to 2022. J. Fish Dis. 2023, 00, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Everson, J.L.; Jones, D.R.; Taylor, A.K.; Rutan, B.J.; Leeds, T.D.; Langwig, K.E.; Wargo, A.R.; Wiens, G.D. Aquaculture Reuse Water, Genetic Line, and Vaccination Affect Rainbow Trout (Oncorhynchus mykiss) Disease Susceptibility and Infection Dynamics. Front. Immunol. 2021, 12, 721048. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Shen, X.; Shi, X.; Sakandar, H.A.; Quan, K.; Li, Y.; Jin, H.; Kwok, L.-Y.; Zhang, H.; Sun, Z. Targeting gut microbiota and metabolism as the major probiotic mechanism—An evidence-based review. Trends Food Sci. Technol. 2023, 138, 178–198. [Google Scholar] [CrossRef]
- Darbandi, A.; Asadi, A.; Mahdizade Ari, M.; Ohadi, E.; Talebi, M.; Halaj Zadeh, M.; Darb Emamie, A.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and potential use as antimicrobials. J. Clin. Lab. Anal. 2022, 36, e24093. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, T.; Ruiz-Zarzuela, I.; de Blas, I.; Balcázar, J.L. Probiotics in aquaculture: A current assessment. Rev. Aquac. 2013, 6, 133–146. [Google Scholar] [CrossRef]
- Ramos, M.A.; Weber, B.; Goncalves, J.F.; Santos, G.A.; Rema, P.; Ozorio, R.O. Dietary probiotic supplementation modulated gut microbiota and improved growth of juvenile rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 302–307. [Google Scholar] [CrossRef]
- Khan, H.; Flint, S.H.; Yu, P.L. Determination of the mode of action of enterolysin A, produced by Enterococcus faecalis B9510. J. Appl. Microbiol. 2013, 115, 484–494. [Google Scholar] [CrossRef]
- Abramov, V.M.; Kosarev, I.V.; Machulin, A.V.; Deryusheva, E.I.; Priputnevich, T.V.; Panin, A.N.; Chikileva, I.O.; Abashina, T.N.; Manoyan, A.M.; Ahmetzyanova, A.A.; et al. Ligilactobacillus salivarius 7247 Strain: Probiotic Properties and Anti-Salmonella Effect with Prebiotics. Antibiotics 2023, 12, 1535. [Google Scholar] [CrossRef]
- O’Callaghan, J.; Butto, L.F.; MacSharry, J.; Nally, K.; O’Toole, P.W. Influence of adhesion and bacteriocin production by Lactobacillus salivarius on the intestinal epithelial cell transcriptional response. Appl. Environ. Microbiol. 2012, 78, 5196–5203. [Google Scholar] [CrossRef]
- Iyer, N.; Williams, M.A.; O‘Callaghan, A.A.; Dempsey, E.; Cabrera-Rubio, R.; Raverdeau, M.; Crispie, F.; Cotter, P.D.; Corr, S.C. Lactobacillus salivarius UCC118 Dampens Inflammation and Promotes Microbiota Recovery to Provide Therapeutic Benefit in a DSS-Induced Colitis Model. Microorganisms 2022, 10, 1383. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A. Probiotics: Nature′s medicine. Int. J. Nutr. Pharmacol. Neurol. Dis. 2013, 3, 219–228. [Google Scholar] [CrossRef]
- Bu, Y.; Liu, Y.; Zhang, T.; Liu, Y.; Zhang, Z.; Yi, H. Bacteriocin-Producing Lactiplantibacillus plantarum YRL45 Enhances Intestinal Immunity and Regulates Gut Microbiota in Mice. Nutrients 2023, 15, 3437. [Google Scholar] [CrossRef]
- Amin, M.; Adams, M.B.; Burke, C.M.; Bolch, C.J.S. Screening and activity of potential gastrointestinal probiotic lactic acid bacteria against Yersinia ruckeri O1b. J. Fish. Dis. 2023, 46, 369–379. [Google Scholar] [CrossRef]
- Wei, S.; Bahl, M.I.; Baunwall, S.M.D.; Hvas, C.L.; Licht, T.R. Determining Gut Microbial Dysbiosis: A Review of Applied Indexes for Assessment of Intestinal Microbiota Imbalances. Appl. Environ. Microbiol. 2021, 87, e00395-21. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Kong, Q.; Wang, Y.; Duan, X.; Wang, P.; Li, C.; Huan, Y. Colonization of Clostridium butyricum in Rats and Its Effect on Intestinal Microbial Composition. Microorganisms 2021, 9, 1573. [Google Scholar] [CrossRef]
- Kapoor, B.; Gulati, M.; Gupta, R.; Singla, R.K. Microbiota dysbiosis and myasthenia gravis: Do all roads lead to Rome? Autoimmun. Rev. 2023, 22, 103313. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; Xie, P.; Yang, Y.; Yan, L.; Guo, A.; Yang, J.; Zhang, J.; Liu, L.; Wang, Q.; Gao, X. Effects of Nisin, Cecropin, and Penthorum chinense Pursh on the Intestinal Microbiome of Common Carp (Cyprinus carpio). Front. Nutr. 2021, 8, 729437. [Google Scholar] [CrossRef]
- Cheng, J.; Hu, J.; Geng, F.; Nie, S. Bacteroides utilization for dietary polysaccharides and their beneficial effects on gut health. Food Sci. Human Wellness 2022, 11, 1101–1110. [Google Scholar] [CrossRef]
- Fan, Z.; Wu, D.; Li, J.; Zhang, Y.; Cui, Z.; Li, T.; Zheng, X.; Liu, H.; Wang, L.; Li, H. Assessment of Fish Protein Hydrolysates in Juvenile Largemouth Bass (Micropterus salmoides) Diets: Effect on Growth, Intestinal Antioxidant Status, Immunity, and Microflora. Front. Nutr. 2022, 9, 816341. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, N.; Lindqvist, L.L.; Wibowo, M.; Sonnenschein, E.C.; Bentzon-Tilia, M.; Gram, L. Role is in the eye of the beholder-the multiple functions of the antibacterial compound tropodithietic acid produced by marine Rhodobacteraceae. FEMS Microbiol. Rev. 2022, 46, fuac007. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Bi, D.; Zheng, W.; Xu, T. MicroRNA negatively regulates NF-kappaB-mediated immune responses by targeting NOD1 in the teleost fish Miichthys miiuy. Sci. China Life Sci. 2021, 64, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Barnabei, L.; Laplantine, E.; Mbongo, W.; Rieux-Laucat, F.; Weil, R. NF-kappaB: At the Borders of Autoimmunity and Inflammation. Front. Immunol. 2021, 12, 716469. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Sun, Z.; Liang, D.; Li, H. Lactobacillus salivarius alleviates inflammation via NF-kappaB signaling in ETEC K88-induced IPEC-J2 cells. J. Anim. Sci. Biotechnol. 2020, 11, 76. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, A.; Zeng, X.; Hou, C.; Liu, H.; Qiao, S. Lactobacillus reuteri I5007 modulates tight junction protein expression in IPEC-J2 cells with LPS stimulation and in newborn piglets under normal conditions. BMC Microbiol. 2015, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Hsu, W.F.; Chang, J.S.; Shih, C.K. Combination of Lactobacillus acidophilus and Bifidobacterium animalis subsp. lactis Shows a Stronger Anti-Inflammatory Effect than Individual Strains in HT-29 Cells. Nutrients 2019, 11, 969. [Google Scholar] [CrossRef]
- Hafsan, H.; Saleh, M.M.; Zabibah, R.S.; Obaid, R.F.; Jabbar, H.S.; Mustafa, Y.F.; Sultan, M.Q.; Gabr, G.A.; Ramirez-Coronel, A.A.; Khodadadi, M.; et al. Dietary Thymol Improved Growth, Body Composition, Digestive Enzyme Activities, Hematology, Immunity, Antioxidant Defense, and Resistance to Streptococcus iniae in the Rainbow Trout (Oncorhynchus mykiss). Aquac. Nutr. 2022, 2022, 3288139. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Sheikhzadeh, N.; Hamidian, G.; Mardani, K.; Oushani, A.K.; Firouzamandi, M.; Esteban, M.A.; Shohreh, P. Changes in rainbow trout (Oncorhynchus mykiss) growth and mucosal immune parameters after dietary administration of grape (Vitis vinifera) seed extract. Fish. Physiol. Biochem. 2021, 47, 547–563. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Direction | Primer Sequences (5′–3′) |
|---|---|---|
| IL-6 | F | ACTCCCCTCTGTCACACACC |
| R | GGCAGACAGGTCCTCCACTA | |
| IL-8 | F | AGAATGTCAGCCAGCCTTGT |
| R | TCTCAGACTCATCCCCTCAGT | |
| TNF-α- | F | GGGGACAAACTGTGGACTGA |
| R | GAAGTTCTTGCCCTGCTCTG | |
| IFN-γ | F | AAGGGCTGTGATGTGTTTCTG |
| R | TGTACTGAGCGGCATTACTCC | |
| NF-κB | F | CAGCGCAGAGAACAACGAAT |
| R | CCGAAGACAGGCTCAGGTAG | |
| ef1α | F | CATGCCTGGTGACAATGTTGG |
| R | TGGGGGCATCCTCAAGTTTC |
| Sample | BWI (g) | BWF (g) | WGR (g) | SGR (%/d) |
|---|---|---|---|---|
| CK | 7.02 ± 0.34 | 9.33 ± 0.80 | 2.26 ± 0.35 a | 1.28 ± 0.15 a |
| A | 7.53 ± 0.48 | 10.23 ± 0.87 | 2.70 ± 0.46 b | 1.45 ± 0.16 b |
| B | 7.14 ± 0.37 | 9.46 ± 0.42 | 2.32 ± 0.15 a | 1.34 ± 0.09 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Wang, B.; Ren, W.; Wang, J.; Liu, Q.; Ren, Y.; Meng, X. Mechanism of Ligilactobacillus salivarius GX118 in Regulating the Growth of Rainbow Trout (Oncorhynchus mykiss) and Resistance to Aeromonas salmonicida Infection. Fishes 2024, 9, 157. https://doi.org/10.3390/fishes9050157
Cao X, Wang B, Ren W, Wang J, Liu Q, Ren Y, Meng X. Mechanism of Ligilactobacillus salivarius GX118 in Regulating the Growth of Rainbow Trout (Oncorhynchus mykiss) and Resistance to Aeromonas salmonicida Infection. Fishes. 2024; 9(5):157. https://doi.org/10.3390/fishes9050157
Chicago/Turabian StyleCao, Xiyu, Bowen Wang, Wenhao Ren, Jiang Wang, Qi Liu, Yichao Ren, and Xianliang Meng. 2024. "Mechanism of Ligilactobacillus salivarius GX118 in Regulating the Growth of Rainbow Trout (Oncorhynchus mykiss) and Resistance to Aeromonas salmonicida Infection" Fishes 9, no. 5: 157. https://doi.org/10.3390/fishes9050157
APA StyleCao, X., Wang, B., Ren, W., Wang, J., Liu, Q., Ren, Y., & Meng, X. (2024). Mechanism of Ligilactobacillus salivarius GX118 in Regulating the Growth of Rainbow Trout (Oncorhynchus mykiss) and Resistance to Aeromonas salmonicida Infection. Fishes, 9(5), 157. https://doi.org/10.3390/fishes9050157