Abstract
In recent years, the Chinese hook snout carp (Opsariichthys bidens) is becoming increasingly popular due to its high nutritional value and delicious taste. However, anthropogenic influences have caused a rapid reduction in its population. Considering long-term development sustainability, it is essential to conduct research on its reproductive development. In this study, the germ-cell related genes dazl (Obdazl) and dnd (Obdnd) were cloned, and their expression patterns were revealed. RT-PCR analysis indicated that Obdazl and Obdnd were specifically expressed in gonads. In the testis, Obdazl and Obdnd RNAs have a similar expression pattern: their transcripts were abundant in spermatogonia, gradually decreased, and eventually disappeared during spermatogenesis. Early oocytes in the ovary exhibited strong expression of Obdazl and Obdnd RNAs and reduced remarkably after peak expression at III oocytes. Surprisingly, Obdazl RNA was dynamically found in perinuclear spots that finally aggregated into the Balbiani body (Bb) in the early oocytes. Moreover, Obdazl 3′ UTR enabled the reporter gene to be expressed stably in medaka PGCs, suggesting that dazl was identified as the germ cell marker in O. bidens, which may provide perspectives for further studies on the development of PGCs in the future.
Key Contribution:
(1) Obdazl and Obdnd were specifically expressed in both male and female gonads of Opsariichthys bidens. (2) Obdnd and Obdazl were enriched in spermatogonia and spermatocytes. (3) Obdazl was concentrated in the Balbiani body in ovary of Opsariichthys bidens. (4) gfp-Obdazl 3′ UTR mRNA could label medaka PGCs.
1. Introduction
Germ cells are of crucial importance in the capability of sexually reproducing organisms to produce bisexual gametes (sperm and eggs), which ensures the transmission of genetic information. Their progenitor cells, called primordial germ cells (PGCs), in Osteichthyes, become independent at the early stage of embryonic development and perform the functions associated with reproductive development. While the initial site of PGCs generation may vary slightly among species, they ultimately migrate into the gonads [1,2]. The formation, specification, and differentiation of PGCs are different across species, which are regulated by a series of genes related to reproduction. Among these, dead-end (dnd) plays a critical role in the specification and development of the PGCs, while dazl can indirectly regulate the germ cell differentiation, growth, and maturation through the interaction of target mRNAs with proteins [3,4].
Dazl belongs to the DAZ (Deleted in Azoospermia) family, which is the earliest fertility factor found in humans. Within the DAZ family, there are three members: daz, boule, and dazl, all of which are necessary regulators for sexual reproduction, especially spermatogenesis [5]. Among them, boule is expressed in meiotic cells, widely distributed from invertebrate fruit flies (Drosophilid) [6] and nematodes (Caenorhabditis Elegans) [7] to mice (Mus musculus) [5] and humans [8]. Daz, whose deletion may cause severe oligospermia or azoospermia, exists only in primates or higher mammals [7,9,10]. In vertebrates, dazl is expressed in primordial germ cells and/or in pre-meiotic and meiotic germ cells [4,11]. In medaka (Oryzias latipes), dazl is referred to as a gonadal marker and counts in the PGC formation. Knockdown of dazl cannot prevent the formation of PGCs, while the injection of dazl antibody induces a significant decrease in PGCs, even resulting in the failure of its formation [12]. In mice, it has been found that the protein DAZL can repress and activate mRNA translation during oocyte maturation [13]. Furthermore, overexpression of dazl shows testis-biased expression patterns in Hu sheep and can reprogram human Sertoli cells into male germline stem cells, suggesting that dazl plays a crucial part in male reproductive development [14,15].
Dnd was first identified in zebrafish as a vital component of germplasm. It is specifically expressed in germ cells, and when knocked down can result in abnormal migration of PGCs [16]. Its homologous genes are widely found in frogs (Xenopus laevis) [17], mice [18], chickens (Gallus gallus) [19], medaka [20], Chinese sturgeon (Acipenser sinensis) [21], Celebes medaka (Oryzias celebensis) [22], Atlantic salmon (Salmo salar) [23], Starry Flounder (Platichthys stellatus) [24], and Spinyhead croaker (Collichthys lucidus) [25]. In medaka and Celebes medaka, dnd is bisexually and differentially expressed in gonads. In the testis, dnd is abundant in spermatogonia and remarkably decreases and even disappears with spermatogenesis, but it is present throughout oogenesis. Moreover, dnd is regarded as a critical PGCs specifier, the knockdown of which by morpholino specifically abolishes PGCs, while its overexpression boosts PGCs via the increase in its precursors [20,22,26]. Attributed to the protection of dnd in the face of signals leading to somatic cell fate during PGCs migration, PGCs differentiate into gametes during embryogenesis, which ensures the fertility of the organism [27]. Recently, knockout of dnd utilizing CRISPR-Cas9 in S. salar helped to achieve sterile fish which successfully solve the genetic introgression in aquaculture, suggesting that sterile fish can also function as recipients for germ cell transplantation [3,23,28,29]. The PGCs-depleted gonads, due to the knockdown of dnd, benefit the function exploration of PGCs in hermaphroditic fish and eventually give support to the establishment of sex control techniques [30]. Furthermore, recent studies have shown that the protein Dnd1 is able to regulate the function of germ granules [31].
Chinese hook snout carp (Opsariichthys bidens) is a small, freshwater, economically farmed species with a delicious taste and high nutrition value, widely distributed in China [32]. However, due to the excessive fishing intensity and environmental destruction in recent years, the wild resources of O. bidens are very scarce. The conservation of its germplasm resources is an urgent problem to be solved. It is thereby imperative to increase the yield of O. bidens and strengthen the research on its artificial reproduction. However, studies on O. bidens focused mostly on its morphology, histology, genome assembly, and biological characteristics, suggesting that research on reproductive biology is urgently needed [33,34,35,36,37]. Our research group has recently succeeded in establishing a spermatogonial stem cell line capable of normal long-term culture in O. bidens [38]. Furthermore, a series of sex-related genes and sex-biased miRNAs have been revealed by us using RNA-Seq in O. bidens [36,39]. In this study, Obdazl and Obdnd complementary DNA were cloned, and RT-PCR results revealed that both were only expressed in gonads. For further research, to verify its ability to label PGCs, a chimeric mRNA of gfp and O. bidens dazl 3′ UTR (gfp-Obdazl 3′ UTR) was injected into medaka embryos by microinjection. The results revealed that dazl was a germline marker, which seemed to be an underlying gene affecting the process of germ cell development in O. bidens. In brief, the purpose of this study was to provide a new reference for the study of the potential role of reproductive development-related genes in the development of PGCs in fishes, as well as the exploration of genes for germ cell markers in O. bidens.
2. Materials and Methods
2.1. Fish and Sampling
The O. bidens used in this experiment were purchased from Shunchang County Zhaoxing Fish Breeding Co., Ltd. (Nanping, China) and subsequently reared in the recirculating aquaculture systems at Shanghai Ocean University (Shanghai, China). During this period, the O. bidens culture systems were maintained for stabilization (water temperature 26 ± 2 °C, dissolved oxygen > 6.2 mg/L, pH 7.4 ± 2) and the systems were cleaned regularly.
Three male and female adult O. bidens were used for adult tissue sampling. Fish are first anesthetized on the ice, and then several tissues (brain, eye, liver, kidney, gut, ovary, and testis) are dissected form them. At the same time, a pair of O. bidens with distinct secondary sex characteristics were selected for artificial insemination to obtain embryos at different developmental stages (2-cell, blastula, gastrula, neurula, 30 hpf, 48 hpf, and fry), and twenty embryos from each period were collected for subsequent experiments.
All experiments were strictly conducted in the direction of the guidelines of the Shanghai Ocean University Committee for Laboratory Animal Research.
2.2. RNA Isolation and cDNA Synthesis
Total RNA was extracted from embryos at different developmental stages and adult tissues using TRizol (Invitrogen, Carlsbad, CA, USA). RNA integrity was checked by 1% agarose gel electrophoresis, stained with ethidium bromide. The RNA quantity was examined by a NanoDrop 2000 c spectrophotometer (Thermo Scientific, Waltham, MA, USA) and the 1 μg of total RNA was used to synthesize the corresponding cDNA by reverse transcription using the M-MLV Reverse Transcription Kit (Takara, Shiga, Japan) and the oligo dT18.
2.3. Molecular Cloning and Analysis of Obdazl and Obdnd Gene
Previously, cDNA fragments corresponding to Obdazl and Obdnd have been obtained from the gonadal transcriptome that was described previously [35]. Subsequently, two pairs of specific primers were constructed from these fragments, respectively, and full-length cDNA of Obdazl and Obdnd were identified by the RACEs approach using the BD SMART RACEs kit (Clontech, Beijing, China). Following that, using the Vector NTI Advance 11.5 software, the Dazl and Dnd proteins were individually aligned across species, and the resulting alignment findings were employed in the MEGA 7.0 program and maximum likelihood method with 1000 repeat of bootstrap method analysis, respectively, to produce phylogenetic trees.
2.4. RT-PCR
To identify the expression of Obdazl and Obdnd in embryos and adult tissues, two sets of primers (Table 1) were constructed, as well as primers for β-actin as an internal reference (Table 1). Each PCR was performed in a reaction system with a volume of 25 μL for a total of 35 cycles, consisting of a denaturation step at 95 °C for 10 s, an annealing step at 58 °C for 10 s, and then an extension step at 72 °C for 1 min; these three steps are included in each cycle. The resultant products of PCR were separated using an agarose gel with a concentration of 1.5%, stained using ethidium bromide, and photographed using a bio-imaging system (Bio-Rad, Hercules, CA, USA).

Table 1.
Sequences of the relevant primers used during the experiments.
2.5. RNA Synthesis and In Situ Hybridization
The O. bidens testis and ovary were embedded in Optimal Cutting Temperature (O.C.T Sakura, Torrance, CA, USA) after fixation in 4% paraformaldehyde (PFA) and graded dehydration with RNase-free sucrose. After embedding, the tissue was sectioned at the thickness of 6 μm using a freezing microtome Leica CM1950 (Leica, Wetzlar, Germany). The subcellular localization of Obdazl and Obdnd RNA was investigated by using both whole-mount in situ hybridization (WISH) and gonadal section in situ hybridization (SISH). Furthermore, the expression of Obdazl together with Obdnd in the ovary was also distinguished by dual fluorescence in situ hybridization (FISH). WISH, SISH, and FISH were carried out as previously described [40,41]. Briefly, two plasmids (pT-Obdazl and pT-Obdnd) were obtained by inserting the open reading frames (ORF) of Obdazl and Obdnd into the pGEM-T vector species (Promega, Fitchburg, WI, USA). The SP6/T7 promoter was employed to generate the sense and antisense probes, which were tagged with the FITC labeling kit (Roche, Basel, Switzerland) and Digoxigenin (DIG) labeling kit (Roche).
2.6. Preparation of Chimeric mRNAs and Microinjection
Construction of the pCSpfObdazl 3′ UTR (gfp-Obdazl 3′ UTR) recombinant plasmid was completed after the Clvasa 3′ UTR in pCSpfClvasa 3′ UTR (gfp-Clvasa 3′ UTR) had been replaced with the Obdazl 3′ UTR (amplified by the primer Obdazl 3′ UTR F&R) [42]. Capped mRNAs for microinjection were created from the fully linearized gfp-Obdazl 3′ UTR and rfp-Drnos1 3′ UTR by using the mMESSAGE mMACHINE kit (Ambion, Austin, TX, USA). The plasmid pCSchDrnos1 3′ UTR has been previously described [22]. Co-injection of the two chimeric mRNAs into one-cell-stage medaka embryos at a dose of 100 ng/μL resulted in the observation of PGCs.
2.7. Microscopy
The Ni-E microscope with a Nikon Ds-Ri2 camera (Nikon, Tokyo, Japan) was utilized in order to carry out microscopy and photography, as previously explained [43].
3. Results
3.1. Cloning and Characterization of Obdazl and Obdnd
Employing the RACEs method, the full-length cDNA sequences of Obdazl and Obdnd were cloned. A 1548 bp Obdazl cDNA was obtained, containing a 690 bp coding sequence (CDS), which encoded 229 amino acids and 3′ UTR 858 bp. It contained a DAZ domain and a conserved RNA recognition domain (RRM) (Supplementary Figure S1). The full-length Obdnd cDNA was 1592 bp in size, including 12 bp 5′ UTR, 1107 bp CDS encoding 368 amino acids, and 473 bp 3′ UTR, which included RNA recognition motif (RRM) as well as the domains, N-terminal region (NR), and C-terminal regions (CR1–4) (Supplementary Figure S2).
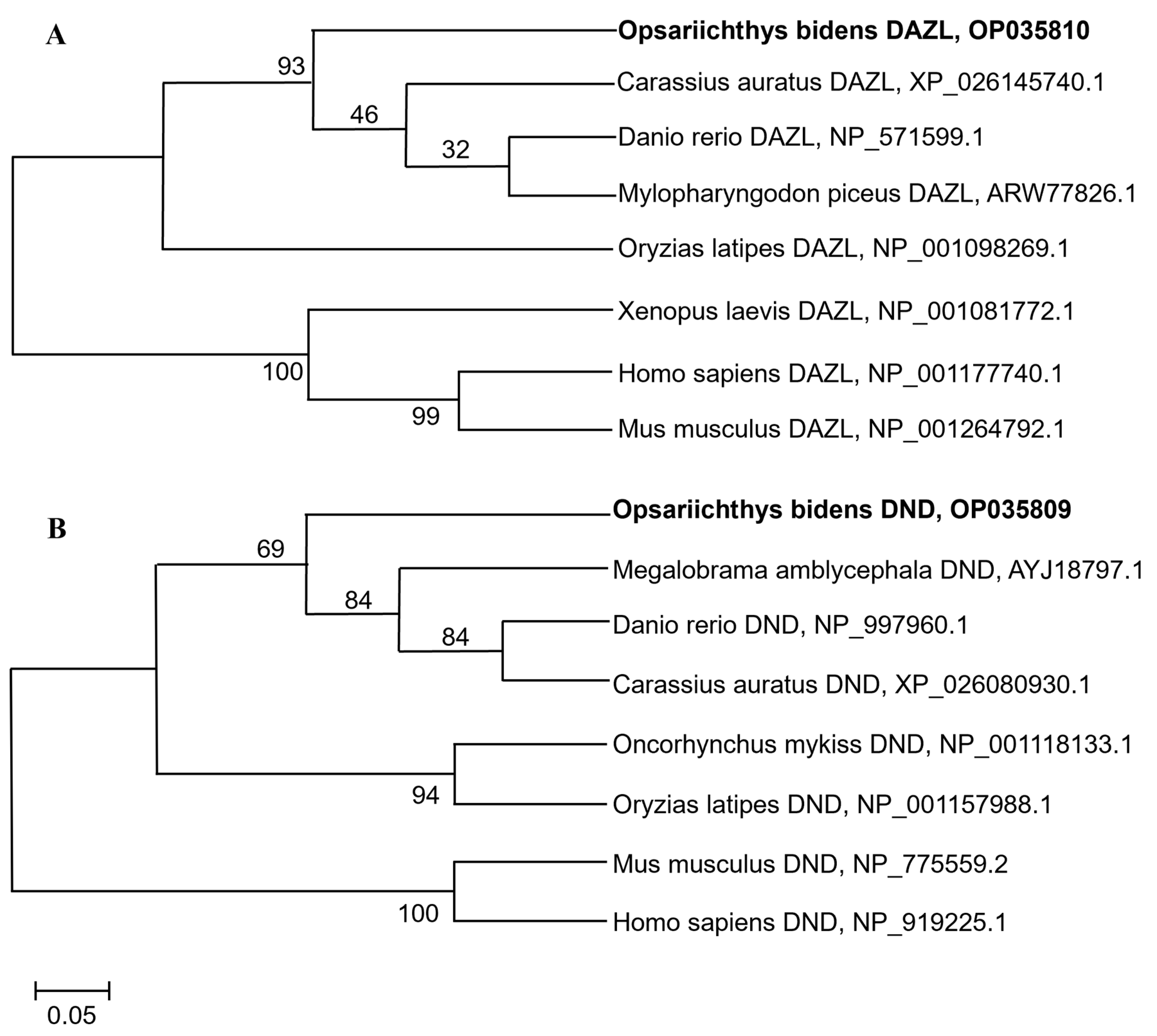
The resulting ObDazl protein was found to have the greatest degree of similarity to the Dazl ortholog in Danio rerio (90.83%) according to the results of multiple amino acid alignments, and the homology with that in humans was 28.89% (Supplementary Figure S3). ObDnd protein also had high homology with Dnd protein in Danio rerio, but its identity was only 60.83% (Supplementary Figure S4), which was significantly lower than that with ObDazl. ObDazl and ObDnd proteins were identified in two separate clades in fish and other vertebrates, according to phylogenetic tree analysis (Figure 1).
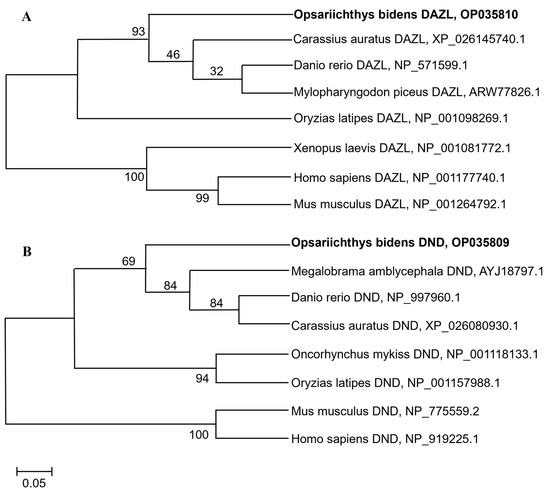
Figure 1.
Phylogenetic tree of Dazl and Dnd proteins in different species. (A) Phylogenetic tree of Dazl proteins. (B) Phylogenetic tree of Dnd proteins. The sequences of Dazl and Dnd homologs were all obtained through NCBI. The bootstrap values are presented. The species name is followed by the accession number.
3.2. Germ Cell-Specific Expression of Obdazl and Obdnd RNAs
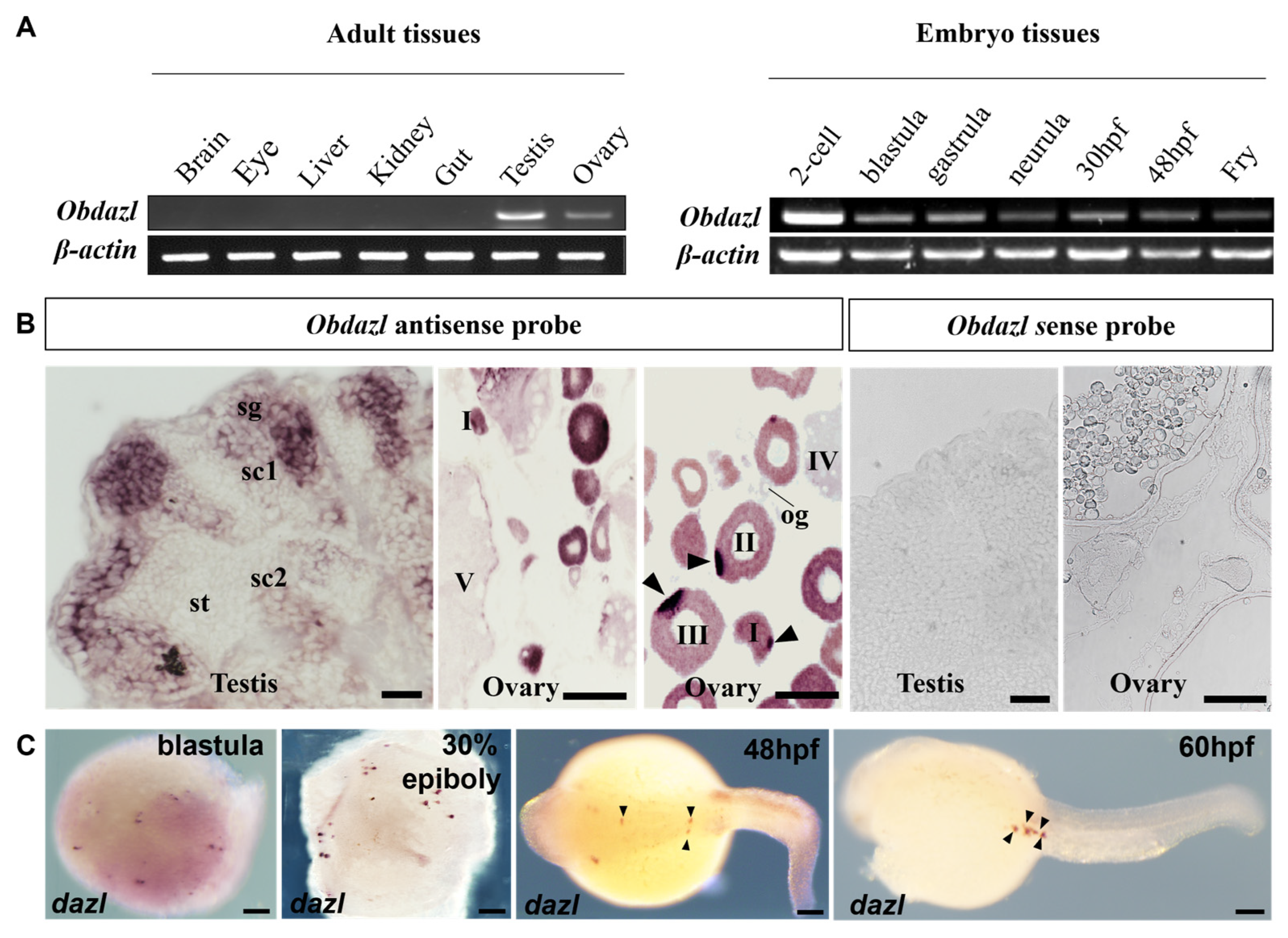
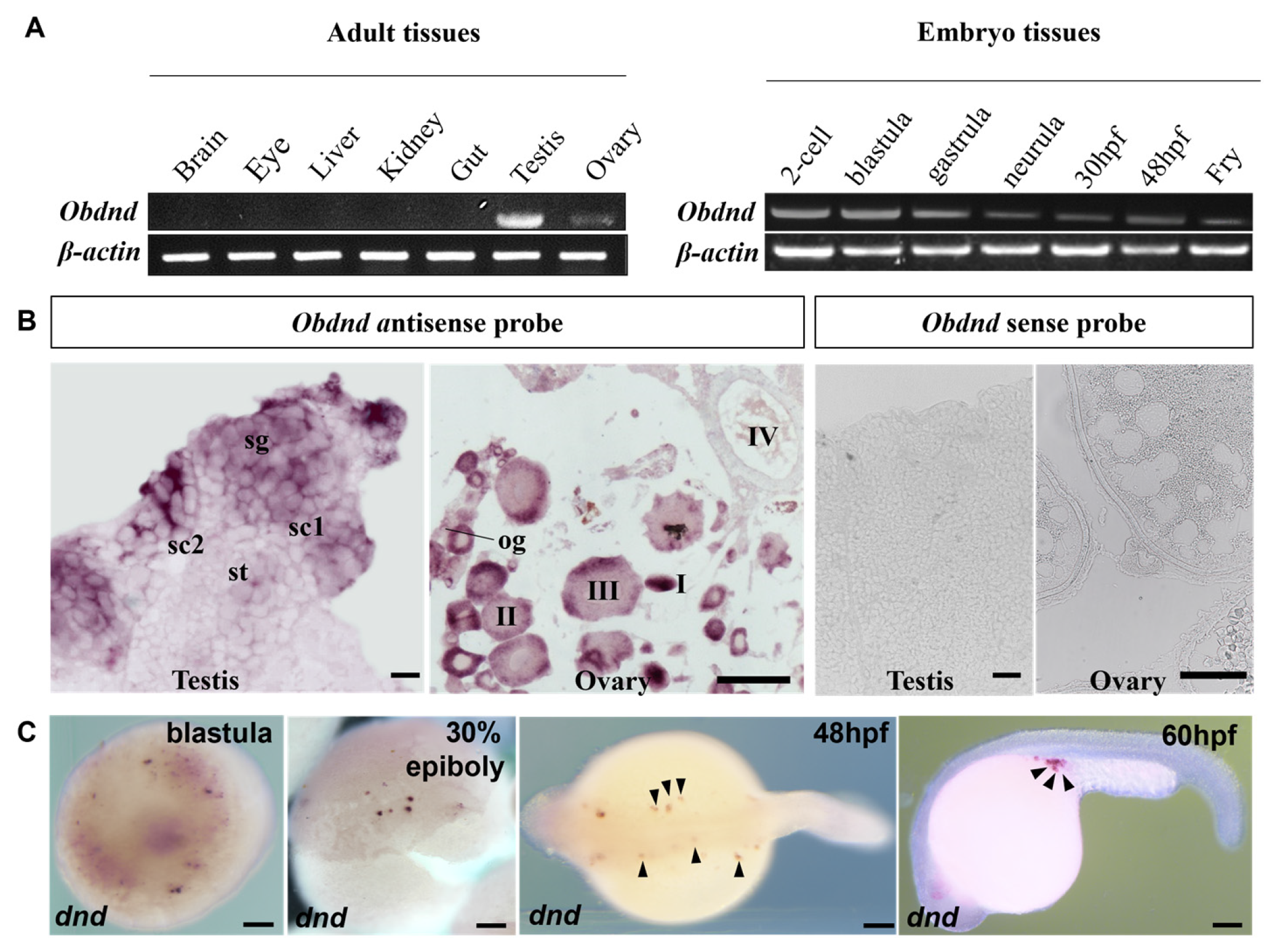
To explore the expression of Obdazl and Obdnd, RT-PCR was conducted. In adult tissues, Obdazl and Obdnd were strongly expressed in the ovary and testis, while neither was detected in body tissues including the brain, eye, liver, kidney, and gut (Figure 2A and Figure 3A). In embryos, Obdazl was strongly expressed in the 2-cell stage and expressed constantly, while the expression of Obdnd gradually weakened during embryogenesis (Figure 2B and Figure 3B).
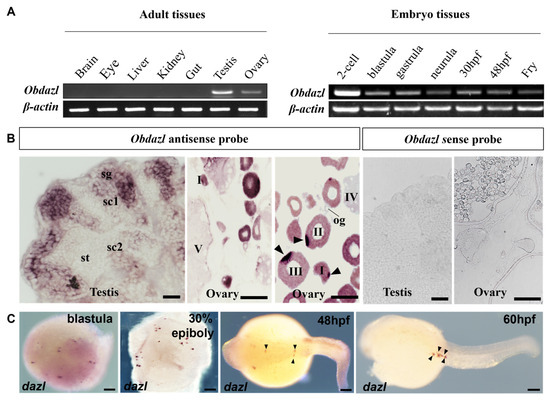
Figure 2.
Expression of Obdazl in different tissues and embryos. (A) RT-PCR analysis of Obdazl in adult tissues and developmental embryos. β-actin served as an internal control. (B) SISH on the testis and ovary by using Obdazl antisense probe and Obdazl sense probe. sg, spermatogonia; sc1 and sc2, primary and secondary spermatocytes; st, spermatids; og, oogonia; I–V, stages of oocytes. (C) WISH on developmental embryos by using Obdazl antisense probe. The arrow pointed to the PGC. Scale bar, 200 μm.
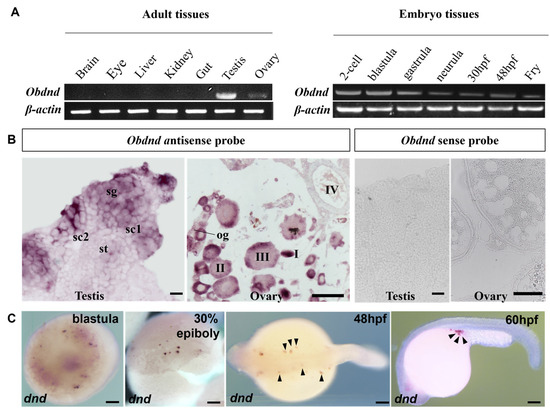
Figure 3.
Expression of Obdnd in different tissues and embryos. (A) RT-PCR analysis of Obdnd in adult tissues and developmental embryos. β-actin served as an internal control. (B) SISH on the testis and ovary by using Obdnd antisense probe and Obdnd antisense probe. sg, spermatogonia; sc1 and sc2, primary and secondary spermatocytes; st, spermatids; og, oogonia; I–IV, stages of oocytes. (C) WISH on developmental embryos by using Obdnd antisense probe. The arrow pointed to the PGC. Scale bar, 200 μm.
Furthermore, SISH and WISH were carried out to detect the temporospatial expression of Obdazl and Obdnd RNAs in gonads. SISH revealed that Obdazl and Obdnd RNAs were examined only in germ cells but were not detectable in somatic cells. (Figure 2B and Figure 3B), which was consistent with the results of RT-PCR (Figure 2A and Figure 3A). The expression of the Obdazl and Obdnd RNAs follows a similar pattern during spermatogenesis. Both Obdazl and Obdnd RNA transcripts were abundant in spermatogonia and remarkably decreased in primary and secondary spermatocytes, with no signals detectable in spermatids (Figure 2B and Figure 3B). In contrast, sense probes as a negative control did not produce any signal above background in the gonads (Figure 2B and Figure 3B).
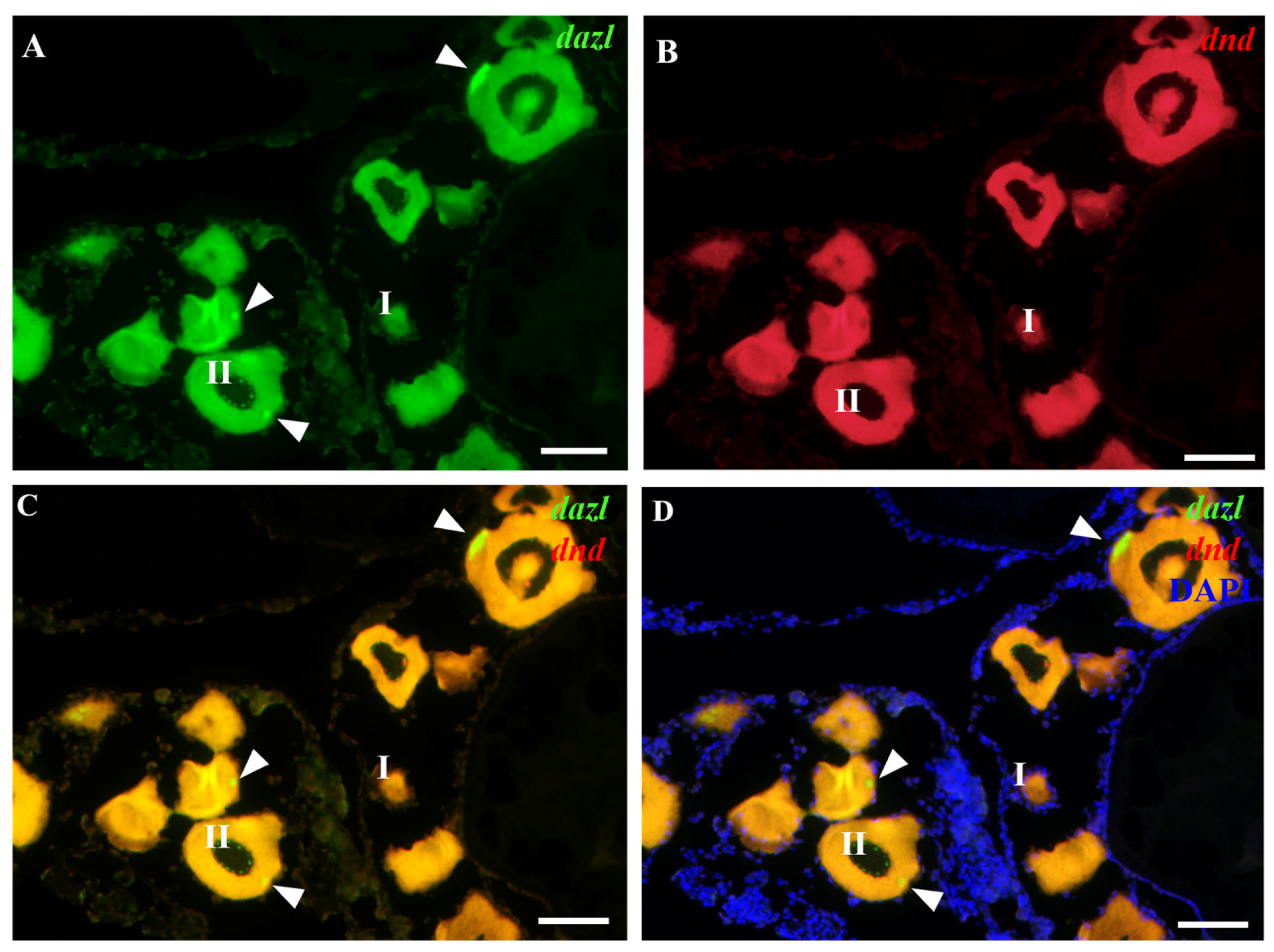
Whereas the expression patterns of Obdazl and Obdnd RNAs in the ovary are not entirely consistent, it is difficult to detect the signals of Obdazl and Obdnd RNAs in oogonia. Obdnd RNA was expressed strongly in oocytes in their early stages I-III, although this level of expression steadily decreased as oogenesis continued (Figure 3B). In an analogous manner, Obdazl RNA was able to be easily found in the cytoplasm around the nucleus of stage I-III oocytes, but its presence was noticeably diminished in stage IV-V oocytes (Figure 2B). Moreover, Obdazl RNA was dynamically found in which perinuclear spots aggregated into the Balbiani body (Bb) in the early oocytes (Figure 2B). To properly validate the distribution of Obdazl and Obdnd, FISH was carried out to evaluate their differential expression in the ovary. Intriguingly, Obdnd was not detected in Bb but was present in large quantities in the ooplasm (Figure 4B). It was in stark contrast to Obdazl, which was concentrated in Bb rather than the ooplasm (Figure 4A). The dual colocalization of FISH validated the different expression patterns between Obdazl and Obdnd in Bb subcellular localization (Figure 4C,D). Again, in contrast to the negative control, sense probes provided no specific signal in gonads (Supplementary Figure S5).
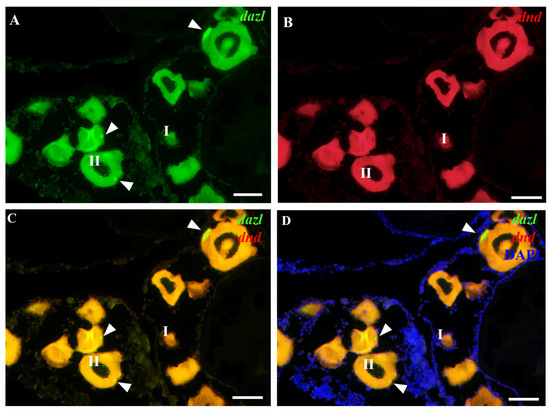
Figure 4.
Localization of Obdnd and Obdazl RNAs in adult ovary. (A,B) In FISH, Obdazl RNA was labeled by green, Obdnd RNA was red, and nuclei were labeled by the blue color of DAPI. (C) Merges of Obdazl with Obdnd. (D) Merges of Obdazl with Obdnd and DAPI. Obdazl concentrates in the Balbiani body, whereas Obdnd is diffuse in the ooplasm and essentially absent in the Balbiani body. I and II, different stages of the oocyte. The arrow pointed to the Balbiani body. Scale bar, 200 μm.
During embryogenesis, the maternal Obdazl and Obdnd RNAs were highly expressed at the cleavage stage (Figure 2A and Figure 3A). During the blastula stage, Obdazl and Obdnd RNAs concentrated in a few conspicuous particles referred to as presumptive PGCs that could differentiate into PGCs as follows (Figure 2C and Figure 3C). Obdazl and Obdnd RNAs were found to be confined in PGCs and bilaterally orientated along the axis as embryogenesis progressed, eventually located in germinal primordia.
3.3. Obdazl 3′ UTR Enables GFP to Express in the PGCs of Medaka Stably
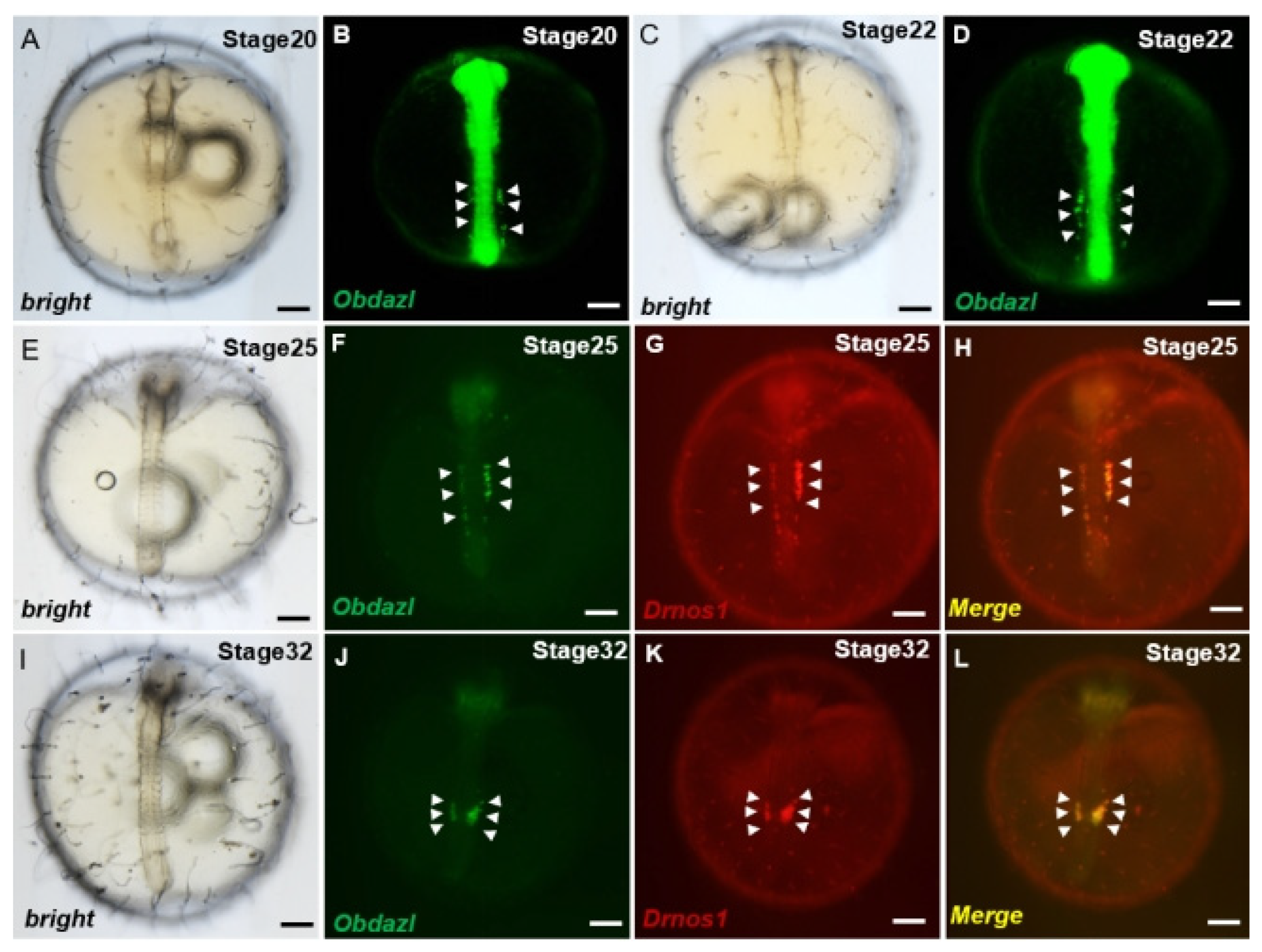
Investigations of PGCs visualization revealed that the 3′ UTR of nanos1 in Danio rerio as a protector of certain proteins was conserved across species, which ensured the stabilization of these RNAs in PGCs [44]. To make clear whether Obdazl 3′ UTR played the same role as that in Danio rerio, the mRNA of gfp-Obdazl 3′ UTR was microinjected into medaka embryos combined with rfp-Drnos1 3′ UTR mRNA, which proved to be effective on the visualization of PGCs (Figure 5). The GFP signaling granules were dispersed throughout the cell during the early stages of embryogenesis. During somitogenesis, cells displaying GFP signaling aggregate dorsally along the trunk on both sides of the embryoid body (Figure 5A–D). These cells made their way finally to the genital ridge by moving along the body of the embryo, where germ cells and mesoderm merged to produce gonads. (Figure 5F,J). Meanwhile, the RFP signal showed a comparable expression pattern to GFP (Figure 5G,K), indicating gfp-Obdazl 3′ UTR mRNA might be applied to mark medaka PGCs. (Figure 5H,L).
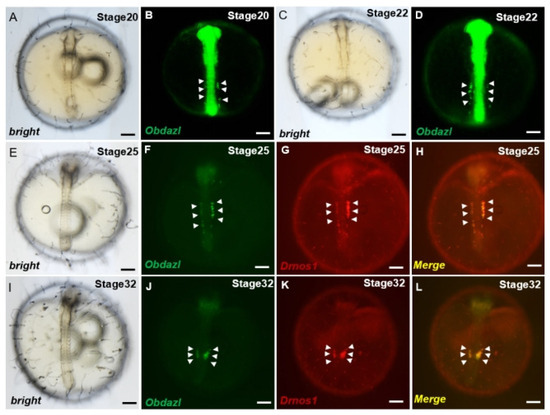
Figure 5.
Visualization of medaka PGCs by Obdazl 3′ UTR. (A–L) During development, PGCs were observed by co-injection of gfp-Obdazl 3′ UTR mRNA and rfp-Drnos1 3′ UTR mRNA. The merged images are presented on the right (H,L). The injected mRNA concentrations were all 100 ng/μL. The arrow pointed to the PGC. Scale bar, 200 μm.
4. Discussion
In this study, Obdazl and Obdnd in full-length were cloned to investigate their differential expression in adult tissues and developmental embryos at various stages. Obdazl and Obdnd RNA transcripts analyzed by ISH appeared to be abundant in the spermatogonia and the oocytes at the early stages. Obdazl RNA was concentrated in the Bb, while Obdnd RNA was found in the ooplasm. In embryogenesis, Obdazl and Obdnd RNAs gathered in presumptive PGCs and eventually migrated into germinal primordia. Furthermore, the Obdazl 3′ UTR enabled the visualization of medaka PGCs migration, confirming its role as the germline marker of O. bidens.
Multiple alignments of Dazl and Dnd proteins manifest that ObDazl and ObDnd are both RNA-binding proteins with conserved RRM domains, indicating that their functions may be similar across species, complying with what has been proven in O. latipes [45], Carassius gibelio [46], O. celebensis [22] and C. nasus [47]. Moreover, Dnd RRM functions as an indispensable indicator for subcellular localization. The RRM mutation induces the failure of Dnd translocation within the germ cell [48]. As for the function of Dazl RRM, it can bind the 3′ UTR of target mRNAs, acting as acceptors as well as activators of their translation [4]. RT-PCR revealed that Obdazl and Obdnd were both specifically expressed in gonads. During spermatogenesis, Obdazl and Obdnd were abundant in mitotic spermatogonia and decreased as meiosis progressed. The phenomenon shows a high similarity to the expression pattern in medaka [20,45], rainbow trout (Oncorhynchus mykiss) [49], and turbot (Scophthalmus maximus) [50]. On the contrary, dazl is hardly expressed in spermatogonia and increases progressively during spermatogenesis in Nile tilapia (Oreochromis niloticus) [51]. Nevertheless, the expressions of Obdazl and Obdnd existed throughout the oogenesis. Their signals could be easily detected at early oocytes and reduced gradually after peak expression at III oocytes, which was in line with dnd expression in O. celebensis [22], large yellow croaker (Larimichthys crocea) and yellow drum (Nibea albiflora) [52]. Strikingly, while Obdnd was abundant in the ooplasm, Obdazl was concentrated in Bb, which acted as a conserved oocyte aggregate in favor of polarized cell formation and was discretely distributed in the cytoplasm [53]. In gibel carp, dazl is located in Bb and facilitates the formation of PGCs [54]. Dazl of the rainbow trout is also concentrated in Bb at the early oocyte stage [49]. However, in the Asian seabass, dazl is concentrated in particles around the nucleus that develops into Bb later in oocytes [55]. Furthermore, further research on Bb function has been carried out in most vertebrates, spreading from mice and frogs to zebrafish. In frogs and zebrafish oocytes, Bb plays a crucial role in the localization of germplasm, which is of great importance for the maternally inherited specification of germ cells [53]. The differences in the expression patterns of Obdazl and Obdnd in the ovary indicated that they may make a difference as distinct factors during oogenesis.
The emergence of PGCs visualization technology has provided a support for further exploration on the functions of genes related to reproductive development. It has been reported that the synthesis of gfp with vasa or nanos1 3′ UTR can stably express in PGCs [44]. In particular, Danio rerio nanos1 3′ UTR can protect the expression of reporter gene and can even effectively mark the PGCs in most of the fish that have been studied [21,26,40,56]. In our study, Obdazl 3′ UTR was selected to combine with the reporter gene to visualize the PGCs. The fact that gfp-Obdazl 3′ UTR mRNA can label medaka PGCs gains support for dazl as a germline marker highly conserved between O. bidens and medaka. Meanwhile, this result can also provide a new angle for the further exploration of the germ cell marker genes in O. bidens, as well as the developmental process of PGCs.
5. Conclusions
In summary, Obdazl and Obdnd show differential and bisexual expression patterns during gametogenesis, suggesting that they fulfill their own part and function in diverse roles. Furthermore, dazl is identified as the germ cell marker of O. bidens, which can benefit further research on reproductive mechanisms.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/fishes9060214/s1: Figure S1: Characteristics of Opsariichthys bidens dazl sequence. Figure S2: Characteristics of Opsariichthys bidens dnd sequence. Figure S3: Multiple alignment of Dazl proteins. Figure S4: Multiple alignment of Dnd proteins. Figure S5: FISH on adult ovary by using Obdazl sense probe and Obdnd sense probe.
Author Contributions
Conceptualization, experimental designing, project administration.: M.L. and W.B.; methodology, supervision, project administration, writing—original draft: Z.Y., L.X. and C.X.; writing—review and editing: M.L., Z.Y., L.X., P.S. and M.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32373130), Fujian Provincial Department of Ocean and Fisheries Special Fund Project (FJHY-YYKJ-2022-2-2) and The Seed Industry Innovation and Industrialization Project of Fujian Province (2021 FJSCZY04).
Institutional Review Board Statement
This study was conducted in accordance with the guidelines of the Declaration of Helsinki and was approved by the Animal Care and Use Committee of the Shanghai Ocean University under approval number SHOU-2023-031.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We are grateful to all workers at the experimental sites.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wylie, C. Germ cells. Cell 1999, 96, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, T.S.; Quagio-Grassiotto, I. Presence of the matrix metalloproteinases during the migration of the primordial germ cells in zebrafish gonadal ridge. Cell Tissue Res. 2021, 383, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Baloch, A.R.; Franek, R.; Saito, T.; Psenicka, M. Dead-end (dnd) protein in fish-a review. Fish Physiol. Biochem. 2021, 47, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.F.; Cheng, S.F.; Wang, L.Q.; Yin, S.; De Felici, M.; Shen, W. DAZ Family Proteins, Key Players for Germ Cell Development. Int. J. Biol. Sci. 2015, 11, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.J.; Lee, M.; Kerr, S.; Ruggiu, M. A murine homologue of the human DAZ gene is autosomal and expressed only in male and female gonads. Hum. Mol. Genet. 1996, 5, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Eberhart, C.G.; Maines, J.Z.; Wasserman, S.A. Meiotic cell cycle requirement for a fly homologue of human Deleted in Azoospermia. Nature 1996, 381, 783–785. [Google Scholar] [CrossRef] [PubMed]
- Karashima, T.; Sugimoto, A.; Yamamoto, M. Caenorhabditis elegans homologue of the human azoospermia factor DAZ is required for oogenesis but not for spermatogenesis. Development 2000, 127, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.Y.; Moore, F.L.; Pera, R.A. A gene family required for human germ cell development evolved from an ancient meiotic gene conserved in metazoans. Proc. Natl. Acad. Sci. USA 2001, 98, 7414–7419. [Google Scholar] [CrossRef] [PubMed]
- Yen, P.H.; Chai, N.N.; Salido, E.C. The human autosomal gene DAZLA: Testis specificity and a candidate for male infertility. Hum. Mol. Genet. 1996, 5, 2013–2017. [Google Scholar] [CrossRef]
- Yen, P.H. Putative biological functions of the DAZ family. Int. J. Androl. 2004, 27, 125–129. [Google Scholar] [CrossRef]
- Wang, W.X.; Liang, S.S.; Zou, Y.X.; Li, Z.; Wu, Q.W.; Wang, L.J.; Wu, Z.H.; Peng, Z.Z.; You, F. Expression of scp3 and dazl reveals the meiotic characteristics of the olive flounder Paralichthys olivaceus(dagger). Biol. Reprod. 2023, 108, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, F.; Li, Z.; Hong, N.; Hong, Y. Dazl is a critical player for primordial germ cell formation in medaka. Sci. Rep. 2016, 6, 28317. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.R.; Rajkovic, G.; Daldello, E.M.; Luong, X.G.; Chen, J.; Conti, M. The RNA-binding protein DAZL functions as repressor and activator of mRNA translation during oocyte maturation. Nat. Commun. 2020, 11, 1399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, W.; Cui, Y.; Wen, L.; Yuan, Q.; Zhou, F.; Qiu, Q.; Sun, M.; Li, Z.; He, Z. Direct reprogramming of human Sertoli cells into male germline stem cells with the self-renewal and differentiation potentials via overexpressing DAZL/DAZ2/BOULE genes. Stem Cell Rep. 2021, 16, 2798–2812. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Deng, M.; Lv, W.; Wei, Z.; Cai, Y.; Cheng, P.; Wang, F.; Zhang, Y. Overexpression of bmp4, dazl, nanos3 and sycp2 in Hu Sheep Leydig Cells Using CRISPR/dcas9 System Promoted Male Germ Cell Related Gene Expression. Biology 2022, 11, 289. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, G.; Stebler, J.; Slanchev, K.; Dumstrei, K.; Wise, C.; Lovell-Badge, R.; Thisse, C.; Thisse, B.; Raz, E. dead end, a novel vertebrate germ plasm component, is required for zebrafish primordial germ cell migration and survival. Curr. Biol. 2003, 13, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Horvay, K.; Claussen, M.; Katzer, M.; Landgrebe, J.; Pieler, T. Xenopus Dead end mRNA is a localized maternal determinant that serves a conserved function in germ cell development. Dev. Biol. 2006, 291, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, C.; Aggarwal, S.; Zhu, R.; Kumar, M.; Zhao, M.; Meistrich, M.L.; Matin, A. The mouse dead-end gene isoform alpha is necessary for germ cell and embryonic viability. Biochem. Biophys. Res. Commun. 2007, 355, 194–199. [Google Scholar] [CrossRef]
- Aramaki, S.; Sato, F.; Kato, T.; Soh, T.; Kato, Y.; Hattori, M.A. Molecular cloning and expression of dead end homologue in chicken primordial germ cells. Cell Tissue Res. 2007, 330, 45–52. [Google Scholar] [CrossRef]
- Liu, L.; Hong, N.; Xu, H.; Li, M.; Yan, Y.; Purwanti, Y.; Yi, M.; Li, Z.; Wang, L.; Hong, Y. Medaka dead end encodes a cytoplasmic protein and identifies embryonic and adult germ cells. Gene Expr. Patterns 2009, 9, 541–548. [Google Scholar] [CrossRef]
- Yang, X.; Yue, H.; Ye, H.; Li, C.; Wei, Q. Identification of a germ cell marker gene, the dead end homologue, in Chinese sturgeon Acipenser sinensis. Gene 2015, 558, 118–125. [Google Scholar] [CrossRef]
- Zhu, T.; Gui, L.; Zhu, Y.; Li, Y.; Li, M. Dnd is required for primordial germ cell specification in Oryzias Celebensis. Gene 2018, 679, 36–43. [Google Scholar] [CrossRef]
- Guralp, H.; Skaftnesmo, K.O.; Kjaerner-Semb, E.; Straume, A.H.; Kleppe, L.; Schulz, R.W.; Edvardsen, R.B.; Wargelius, A. Rescue of germ cells in dnd crispant embryos opens the possibility to produce inherited sterility in Atlantic salmon. Sci. Rep. 2020, 10, 18042. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Cho, Y.S.; Lee, H.B.; Park, J.Y.; Lim, H.K. Dead-End (dnd) Gene Cloning and Gonad-Specific Expression Pattern in Starry Flounder (Platichthys stellatus). Animals 2021, 11, 2256. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Li, Y.; Wen, Z.; Jawad, M.; Gui, L.; Li, M. Spinyhead Croaker Germ Cells Gene dnd Visualizes Primordial Germ Cells in Medaka. Life 2022, 12, 1226. [Google Scholar] [CrossRef] [PubMed]
- Hong, N.; Li, M.; Yuan, Y.; Wang, T.; Yi, M.; Xu, H.; Zeng, H.; Song, J.; Hong, Y. Dnd Is a Critical Specifier of Primordial Germ Cells in the Medaka Fish. Stem Cell Rep. 2016, 6, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Gross-Thebing, T.; Yigit, S.; Pfeiffer, J.; Reichman-Fried, M.; Bandemer, J.; Ruckert, C.; Rathmer, C.; Goudarzi, M.; Stehling, M.; Tarbashevich, K.; et al. The Vertebrate Protein Dead End Maintains Primordial Germ Cell Fate by Inhibiting Somatic Differentiation. Dev. Cell 2017, 43, 704–715.e5. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, R.; Katayama, N.; Sadaie, S.; Miwa, M.; Sanchez Matias, G.A.; Ichida, K.; Fujii, W.; Naito, K.; Hayashi, M.; Yoshizaki, G. Production of Germ Cell-Less Rainbow Trout by dead end Gene Knockout and their Use as Recipients for Germ Cell Transplantation. Mar. Biotechnol. 2022, 24, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hong, N.; Xu, H.; Song, J.; Hong, Y. Germline replacement by blastula cell transplantation in the fish medaka. Sci. Rep. 2016, 6, 29658. [Google Scholar] [CrossRef]
- Hou, M.; Feng, K.; Luo, H.; Jiang, Y.; Xu, W.; Li, Y.; Song, Y.; Chen, J.; Tao, B.; Zhu, Z.; et al. Complete Depletion of Primordial Germ Cells Results in Masculinization of Monopterus albus, a Protogynous Hermaphroditic Fish. Mar. Biotechnol. 2022, 24, 320–334. [Google Scholar] [CrossRef]
- Westerich, K.J.; Tarbashevich, K.; Schick, J.; Gupta, A.; Zhu, M.; Hull, K.; Romo, D.; Zeuschner, D.; Goudarzi, M.; Gross-Thebing, T.; et al. Spatial organization and function of RNA molecules within phase-separated condensates in zebrafish are controlled by Dnd1. Dev. Cell 2023, 58, 1578–1592.e5. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; He, S.; Mayden, R.L. The complete mitochondrial genome of the Chinese hook snout carp Opsariichthys bidens (Actinopterygii: Cypriniformes) and an alternative pattern of mitogenomic evolution in vertebrate. Gene 2007, 399, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Schmidt, B.V.; He, S. The potential colonization histories of Opsariichthys bidens (Cyprinidae) in China using Bayesian binary MCMC analysis. Gene 2018, 676, 1–8. [Google Scholar] [CrossRef]
- Fu, S.J.; Peng, Z.; Cao, Z.D.; Peng, J.L.; He, X.K.; Xu, D.; Zhang, A.J. Habitat-specific locomotor variation among Chinese hook snout carp (Opsariichthys bidens) along a river. PLoS ONE 2012, 7, e40791. [Google Scholar] [CrossRef]
- Perdices, A.; Sayanda, D.; Coelho, M.M. Mitochondrial diversity of Opsariichthys bidens (Teleostei, Cyprinidae) in three Chinese drainages. Mol. Phylogenet. Evol. 2005, 37, 920–927. [Google Scholar] [CrossRef]
- Tang, R.; Zhu, Y.; Gan, W.; Zhang, Y.; Yao, Z.; Ren, J.; Li, M. De novo transcriptome analysis of gonads reveals the sex-associated genes in Chinese hook snout carp Opsariichthys bidens. Aquac. Rep. 2022, 23, 101068. [Google Scholar] [CrossRef]
- Xu, X.; Guan, W.; Niu, B.; Guo, D.; Xie, Q.P.; Zhan, W.; Yi, S.; Lou, B. Chromosome-Level Assembly of the Chinese Hooksnout Carp (Opsariichthys bidens) Genome Using PacBio Sequencing and Hi-C Technology. Front. Genet. 2021, 12, 788547. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kan, Y.T.; Zhong, Y.; Jawad, M.; Wei, W.B.; Gu, K.Y.; Gui, L.; Li, M.Y. Generation of a normal long-term cultured Chinese hook snout carp spermatogonial stem cell line capable of sperm production in vitro. Biology 2022, 11, 1069. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Xu, C.; Zhu, Y.; Yan, J.; Yao, Z.; Zhou, W.; Gui, L.; Li, M. Identification and expression analysis of sex biased miRNAs in Chinese hook snout carp Opsariichthys bidens. Front. Genet. 2022, 13, 990683. [Google Scholar] [CrossRef]
- Li, M.; Hong, N.; Gui, J.; Hong, Y. Medaka piwi is essential for primordial germ cell migration. Curr. Mol. Med. 2012, 12, 1040–1049. [Google Scholar] [CrossRef]
- Yuan, C.; Chen, K.; Zhu, Y.; Yuan, Y.; Li, M. Medaka igf1 identifies somatic cells and meiotic germ cells of both sexes. Gene 2018, 642, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, W.; Zhu, Y.F.; Zhu, T.Y.; Ma, L.B.; Li, M.Y. Evolutionarily conserved vasa identifies embryonic and gonadal germ cells in spinyhead croaker Collichthys lucidus. J. Fish Biol. 2019, 94, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhu, Y.; Yuan, C.; Zhao, Y.; Zhou, W.; Li, M. Differential Expression of Duplicate Insulin-like Growth Factor-1 Receptors (igf1rs) in Medaka Gonads. Life 2022, 12, 859. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Fujimoto, T.; Maegawa, S.; Inoue, K.; Tanaka, M.; Arai, K.; Yamaha, E. Visualization of primordial germ cells in vivo using GFP-nos1 3′UTR mRNA. Int. J. Dev. Biol. 2006, 50, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, M.; Gui, J.; Hong, Y. Cloning and expression of medaka dazl during embryogenesis and gametogenesis. Gene Expr. Patterns 2007, 7, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Z.; Liu, W.; Li, Z.; Wang, Y.; Zhou, L.; Yi, M.S.; Gui, J.F. Molecular characterization and expression pattern of a germ cell marker gene dnd in gibel carp (Carassius gibelio). Gene 2016, 591, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, P.; Xia, J.; Guo, J.; Shi, Y.; Zhong, Y.; Li, M. Evolutionarily conserved boule and dazl identify germ cells of Coilia nasus. Aquac. Fish. 2021, 8, 244–251. [Google Scholar] [CrossRef]
- Slanchev, K.; Stebler, J.; Goudarzi, M.; Cojocaru, V.; Weidinger, G.; Raz, E. Control of Dead end localization and activity—Implications for the function of the protein in antagonizing miRNA function. Mech. Dev. 2009, 126, 270–277. [Google Scholar] [CrossRef]
- Li, M.; Shen, Q.; Xu, H.; Wong, F.M.; Cui, J.; Li, Z.; Hong, N.; Wang, L.; Zhao, H.; Ma, B.; et al. Differential conservation and divergence of fertility genes boule and dazl in the rainbow trout. PLoS ONE 2011, 6, e15910. [Google Scholar] [CrossRef]
- Lin, F.; Zhao, C.Y.; Xu, S.H.; Ma, D.Y.; Xiao, Z.Z.; Xiao, Y.S.; Xu, C.A.; Liu, Q.H.; Li, J. Germline-specific and sexually dimorphic expression of a dead end gene homologue in turbot (Scophthalmus maximus). Theriogenology 2013, 80, 665–672. [Google Scholar] [CrossRef]
- Bhat, N.; Hong, Y. Cloning and expression of boule and dazl in the Nile tilapia (Oreochromis niloticus). Gene 2014, 540, 140–145. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, Y.; Ye, H.; Lu, L.; Li, H.; Xu, Z.; Li, W.; Yin, X.; Xu, D. Identification of germ cells in large yellow croaker (Larimichthys crocea) and yellow drum (Nibea albiflora) using RT-PCR and in situ hybridization analyses. Gene 2023, 863, 147280. [Google Scholar] [CrossRef] [PubMed]
- Jamieson-Lucy, A.; Mullins, M.C. The vertebrate Balbiani body, germ plasm, and oocyte polarity. Curr. Top. Dev. Biol. 2019, 135, 1–34. [Google Scholar] [PubMed]
- Peng, J.X.; Xie, J.L.; Zhou, L.; Hong, Y.H.; Gui, J.F. Evolutionary conservation of Dazl genomic organization and its continuous and dynamic distribution throughout germline development in gynogenetic gibel carp. J. Exp. Zool. B Mol. Dev. Evol. 2009, 312, 855–871. [Google Scholar] [CrossRef] [PubMed]
- Dwarakanath, M.; Lim, M.; Xu, H.; Hong, Y. Differential expression of boule and dazl in adult germ cells of the Asian seabass. Gene 2014, 549, 237–242. [Google Scholar] [CrossRef]
- Koprunner, M.; Thisse, C.; Thisse, B.; Raz, E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes Dev. 2001, 15, 2877–2885. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).