Dynamics of Fractional-Order Three-Species Food Chain Model with Vigilance Effect




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mathematical Model
3. Preliminaries
- •
- The equilibrium point is asymptotically stable if and only if every eigenvalue (for ) of the matrix satisfies the condition .
- •
- The equilibrium is stable if and only if all eigenvalues of meet the condition ; also, any eigenvalue for which must have matching geometric and algebraic multiplicities.
Uniqueness, Boundedness, and Non-Negativity of Solutions
4. Stability and Bifurcation
4.1. Equilibrium Points
4.2. Stability Analysis
4.3. Hopf Bifurcation
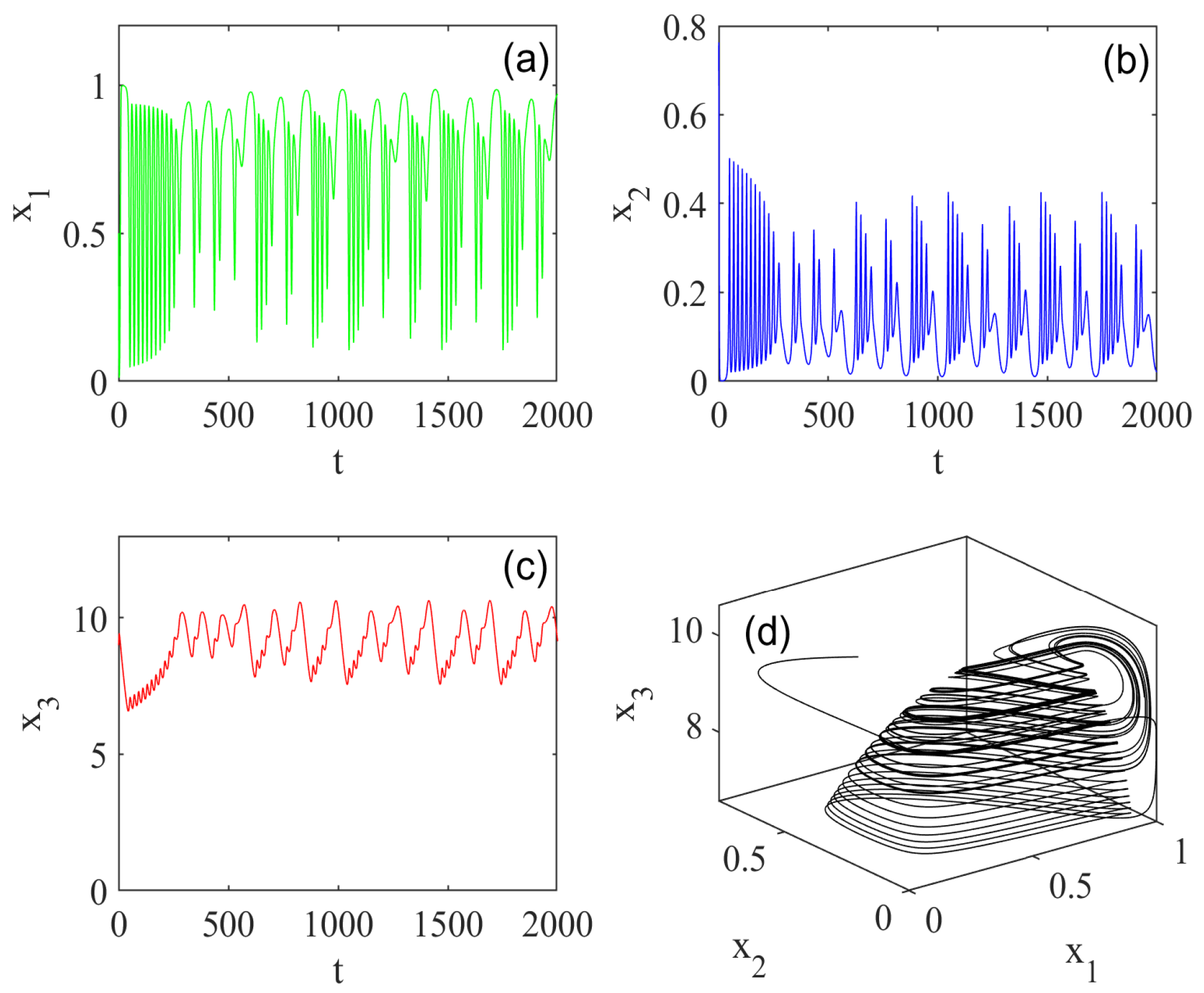
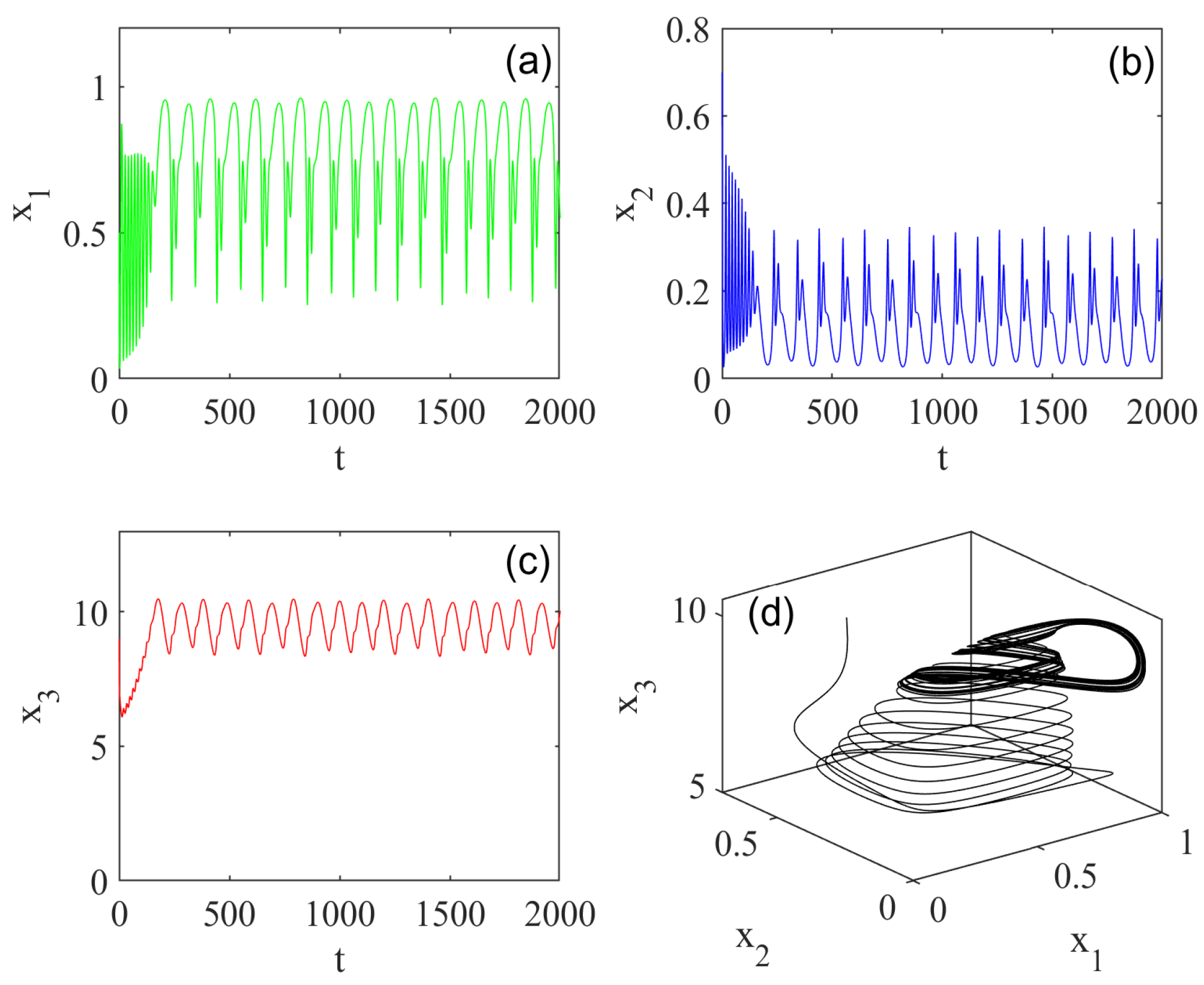
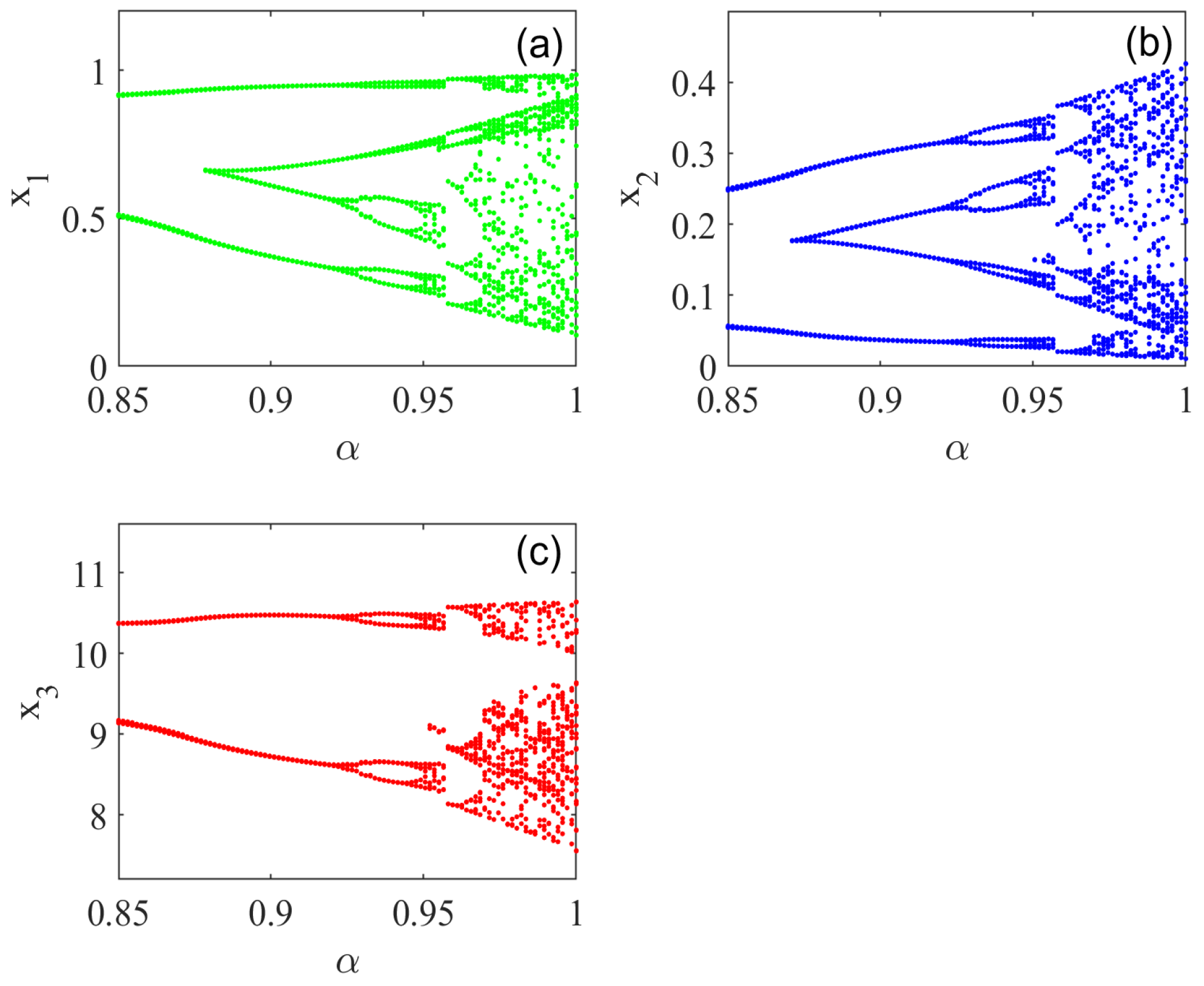
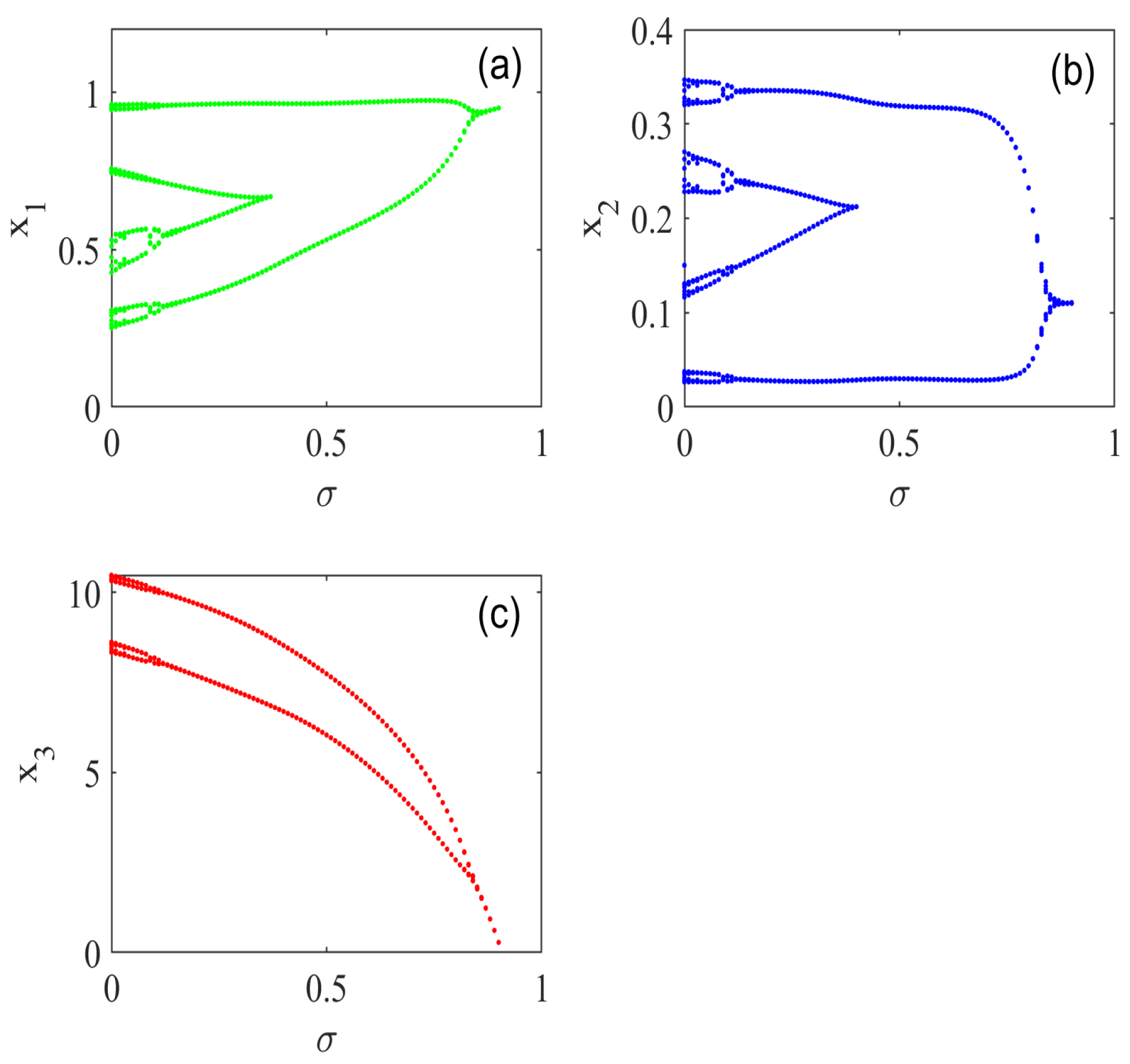
5. Numerical Simulations
5.1. Effect of Fractional Order
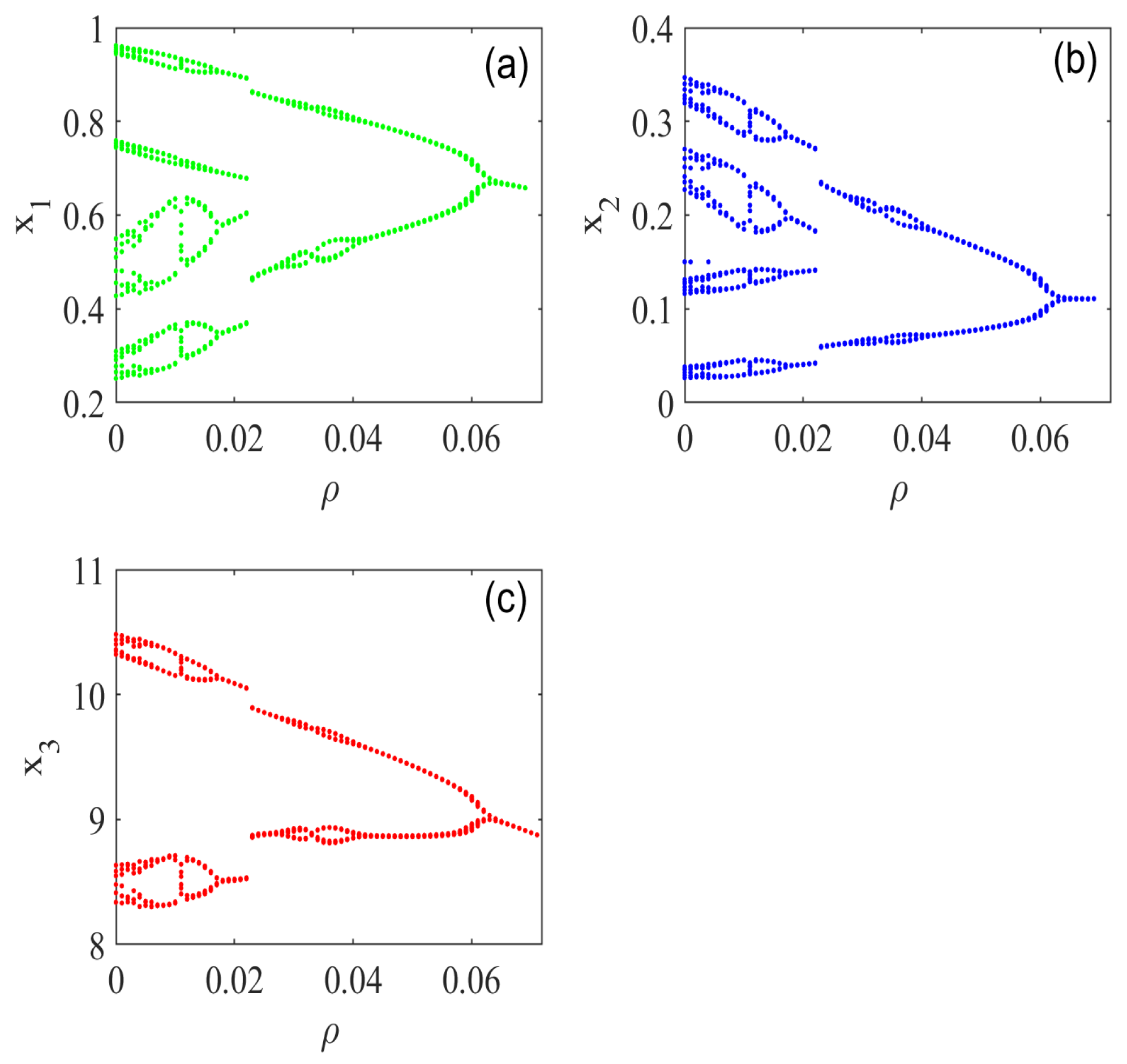
5.2. Effect of Habitat Complexity
5.3. Effect of Basal Prey Vigilance
5.4. Effect of Middle Predator Vigilance
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Symbol | Interpretation |
| T | Time |
| Basal prey density | |
| Middle predator density | |
| Top predator density | |
| B | Birth rate of basal prey |
| Natural death rate of basal prey | |
| Natural death rate of middle predator | |
| Natural death rate of top predator | |
| A | Intraspecific competition rate of basal prey |
| Maximum predation rate of middle predator | |
| Maximum predation rate of top predator | |
| Conversion efficiency of middle predator | |
| Conversion efficiency of top predator | |
| Half-saturation constant of middle predator | |
| Middle predator lethality | |
| Top predator lethality | |
| Level of vigilance of basal prey | |
| Level of vigilance of middle predator | |
| Effectiveness of basal prey’s vigilance | |
| Effectiveness of middle predator’s vigilance |
References
- Hastings, A.; Powell, T. Chaos in a three-species food chain. Ecology 1991, 72, 896–903. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Naji, R.K. Dynamics of a three species food chain model with Crowley–Martin type functional response. Chaos Solitons Fractals 2009, 42, 1337–1346. [Google Scholar] [CrossRef]
- Jana, D.; Agrawal, R.; Upadhyay, R.K. Top-predator interference and gestation delay as determinants of the dynamics of a realistic model food chain. Chaos Solitons Fractals 2014, 69, 50–63. [Google Scholar] [CrossRef]
- Vinoth, S.; Sivasamy, R.; Sathiyanathan, K.; Rajchakit, G.; Hammachukiattikul, P.; Vadivel, R.; Gunasekaran, N. Dynamical analysis of a delayed food chain model with additive Allee effect. Adv. Differ. Equ. 2021, 2021, 54. [Google Scholar] [CrossRef]
- Holling, C.S. The components of predation as revealed by a study of small-mammal predation of the European Pine Sawfly1. Can. Entomol. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism1. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Murray, J.D. Mathematical Biology: I. An Introduction; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; Volume 17. [Google Scholar]
- Canion, C.R.; Heck, K.L., Jr. Effect of habitat complexity on predation success: Re-evaluating the current paradigm in seagrass beds. Mar. Ecol. Prog. Ser. 2009, 393, 37–46. [Google Scholar] [CrossRef]
- Scharf, F.S.; Manderson, J.P.; Fabrizio, M.C. The effects of seafloor habitat complexity on survival of juvenile fishes: Species-specific interactions with structural refuge. J. Exp. Mar. Biol. Ecol. 2006, 335, 167–176. [Google Scholar] [CrossRef]
- Johnson, M.P.; Frost, N.J.; Mosley, M.W.; Roberts, M.F.; Hawkins, S.J. The area-independent effects of habitat complexity on biodiversity vary between regions. Ecol. Lett. 2003, 6, 126–132. [Google Scholar] [CrossRef]
- Hossain, M.; Kumbhakar, R.; Pal, N. Dynamics in the biparametric spaces of a three-species food chain model with vigilance. Chaos Solitons Fractals 2022, 162, 112438. [Google Scholar] [CrossRef]
- Hossain, M.; Garai, S.; Jafari, S.; Pal, N. Bifurcation, chaos, multistability, and organized structures in a predator–prey model with vigilance. Chaos Interdiscip. J. Nonlinear Sci. 2022, 32, 063139. [Google Scholar] [CrossRef]
- Creel, S. The control of risk hypothesis: Reactive vs. proactive antipredator responses and stress-mediated vs. food-mediated costs of response. Ecol. Lett. 2018, 21, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Hunter, L.T.; Skinner, J.D. Vigilance behaviour in African ungulates: The role of predation pressure. Behaviour 1998, 135, 195–211. [Google Scholar] [CrossRef]
- Hossain, M.; Garai, S.; Karmakar, S.; Pal, N.; Chattopadhyay, J. Impact of vigilance on the density variations in a food chain model. Ecol. Complex. 2022, 50, 100996. [Google Scholar] [CrossRef]
- Ramasamy, S.; Banjerdpongchai, D.; Park, P. Stability and Hopf-bifurcation analysis of diffusive Leslie–Gower prey–predator model with the Allee effect and carry-over effects. Math. Comput. Simul. 2025, 227, 19–40. [Google Scholar] [CrossRef]
- Petráš, I. Fractional-Order Nonlinear Systems: Modeling, Analysis and Simulation; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Podlubny, I. Fractional Differential Equations: An Introduction to Fractional Derivatives, Fractional Differential Equations, to Methods of Their Solution and Some of Their Applications; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Ionescu, C.; Lopes, A.; Copot, D.; Machado, J.T.; Bates, J.H. The role of fractional calculus in modeling biological phenomena: A review. Commun. Nonlinear Sci. Numer. Simul. 2017, 51, 141–159. [Google Scholar] [CrossRef]
- Du, M.; Wang, Z.; Hu, H. Measuring memory with the order of fractional derivative. Sci. Rep. 2013, 3, 3431. [Google Scholar] [CrossRef] [PubMed]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. A Rosenzweig–MacArthur model with continuous threshold harvesting in predator involving fractional derivatives with power law and mittag–leffler kernel. Axioms 2020, 9, 122. [Google Scholar] [CrossRef]
- Veeresha, P.; Prakasha, D.; Singh, J.; Khan, I.; Kumar, D. Analytical approach for fractional extended Fisher–Kolmogorov equation with Mittag-Leffler kernel. Adv. Differ. Equ. 2020, 2020, 174. [Google Scholar] [CrossRef]
- Muhammad Altaf, K.; Atangana, A. Dynamics of Ebola disease in the framework of different fractional derivatives. Entropy 2019, 21, 303. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.; Bastos, N.R.; Monteiro, M.T.T. Modeling some real phenomena by fractional differential equations. Math. Methods Appl. Sci. 2016, 39, 4846–4855. [Google Scholar] [CrossRef]
- Padder, A.; Almutairi, L.; Qureshi, S.; Soomro, A.; Afroz, A.; Hincal, E.; Tassaddiq, A. Dynamical analysis of generalized tumor model with Caputo fractional-order derivative. Fractal Fract. 2023, 7, 258. [Google Scholar] [CrossRef]
- Vadivel, R.; Hammachukiattikul, P.; Vinoth, S.; Chaisena, K.; Gunasekaran, N. An extended dissipative analysis of fractional-order fuzzy networked control systems. Fractal Fract. 2022, 6, 591. [Google Scholar] [CrossRef]
- Olayiwola, M.O.; Alaje, A.I.; Olarewaju, A.Y.; Adedokun, K.A. A Caputo fractional order epidemic model for evaluating the effectiveness of high-risk quarantine and vaccination strategies on the spread of COVID-19. Healthc. Anal. 2023, 3, 100179. [Google Scholar] [CrossRef]
- Mondal, S.; Biswas, M.; Bairagi, N. Local and global dynamics of a fractional-order predator–prey system with habitat complexity and the corresponding discretized fractional-order system. J. Appl. Math. Comput. 2020, 63, 311–340. [Google Scholar] [CrossRef]
- Panja, P. Stability and dynamics of a fractional-order three-species predator–prey model. Theory Biosci. 2019, 138, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Panja, P. Dynamics of a fractional order predator-prey model with intraguild predation. Int. J. Model. Simul. 2019, 39, 256–268. [Google Scholar] [CrossRef]
- Garrappa, R. Short Tutorial: Solving Fractional Differential Equations by Matlab Codes; Department of Mathematics, University of Bari: Bari, Italy, 2014. [Google Scholar]
- Caputo, M. Linear models of dissipation whose Q is almost frequency independent—II. Geophys. J. Int. 1967, 13, 529–539. [Google Scholar] [CrossRef]
- Diethelm, K.; Ford, N. The Analysis of Fractional Differential Equations; Lecture Notes in Mathematics; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Matignon, D. Stability results for fractional differential equations with applications to control processing. In Proceedings of the Computational Engineering in Systems Applications, Lille, France, 9–12 July 1996; Volume 2, pp. 963–968. [Google Scholar]
- Li, H.L.; Zhang, L.; Hu, C.; Jiang, Y.L.; Teng, Z. Dynamical analysis of a fractional-order predator-prey model incorporating a prey refuge. J. Appl. Math. Comput. 2017, 54, 435–449. [Google Scholar] [CrossRef]
- Choi, S.; Kang, B.; Koo, N. Stability for fractional differential equations. Proc. Jangjeon Math. Soc. 2013, 16, 165–174. [Google Scholar]
- Choi, S.K.; Kang, B.; Koo, N. Stability for Caputo fractional differential systems. Abstr. Appl. Anal. 2014, 2014, 631419. [Google Scholar] [CrossRef]
- Li, C.; Ma, Y. Fractional dynamical system and its linearization theorem. Nonlinear Dyn. 2013, 71, 621–633. [Google Scholar] [CrossRef]
- Tavazoei, M.S.; Haeri, M.; Attari, M.; Bolouki, S.; Siami, M. More details on analysis of fractional-order Van der Pol oscillator. J. Vib. Control 2009, 15, 803–819. [Google Scholar] [CrossRef]
- Tavazoei, M.S.; Haeri, M. A proof for non existence of periodic solutions in time invariant fractional order systems. Automatica 2009, 45, 1886–1890. [Google Scholar] [CrossRef]
- Tavazoei, M.S. A note on fractional-order derivatives of periodic functions. Automatica 2010, 46, 945–948. [Google Scholar] [CrossRef]
- Abdelouahab, M.S.; Hamri, N.E.; Wang, J. Hopf bifurcation and chaos in fractional-order modified hybrid optical system. Nonlinear Dyn. 2012, 69, 275–284. [Google Scholar] [CrossRef]
- Yazdani, M.; Salarieh, H. On the existence of periodic solutions in time-invariant fractional order systems. Automatica 2011, 47, 1834–1837. [Google Scholar] [CrossRef]
- Danca, M.F. Matlab code for Lyapunov exponents of fractional-order systems, part ii: The noncommensurate case. Int. J. Bifurc. Chaos 2021, 31, 2150187. [Google Scholar] [CrossRef]
- E. Alsubaie, N.; EL Guma, F.; Boulehmi, K.; Al-kuleab, N.; A. Abdoon, M. Improving influenza epidemiological models under Caputo fractional-order calculus. Symmetry 2024, 16, 929. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seralan, V.; Vadivel, R.; Gunasekaran, N.; Radwan, T. Dynamics of Fractional-Order Three-Species Food Chain Model with Vigilance Effect. Fractal Fract. 2025, 9, 45. https://doi.org/10.3390/fractalfract9010045
Seralan V, Vadivel R, Gunasekaran N, Radwan T. Dynamics of Fractional-Order Three-Species Food Chain Model with Vigilance Effect. Fractal and Fractional. 2025; 9(1):45. https://doi.org/10.3390/fractalfract9010045
Chicago/Turabian StyleSeralan, Vinoth, Rajarathinam Vadivel, Nallappan Gunasekaran, and Taha Radwan. 2025. "Dynamics of Fractional-Order Three-Species Food Chain Model with Vigilance Effect" Fractal and Fractional 9, no. 1: 45. https://doi.org/10.3390/fractalfract9010045
APA StyleSeralan, V., Vadivel, R., Gunasekaran, N., & Radwan, T. (2025). Dynamics of Fractional-Order Three-Species Food Chain Model with Vigilance Effect. Fractal and Fractional, 9(1), 45. https://doi.org/10.3390/fractalfract9010045