
Whole Genome Analysis of Streptomyces spp. Strains Isolated from the Rhizosphere of Vitis vinifera L. Reveals Their Role in Nitrogen and Phosphorus Metabolism
 ,
,  ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Bacterial Streptomyces spp. Strains
2.2. Genomic DNA Extraction, Library Preparation, and Sequencing
2.3. Genome De Novo Assembly and Annotation
2.4. Genomes Selection and Phylogenomic Analysis of Streptomyces spp.
2.5. Average Nucleotide Identity Analysis
2.6. Pan- and Core-Genome Analysis
3. Results and Discussion
3.1. Genome De Novo Assembly
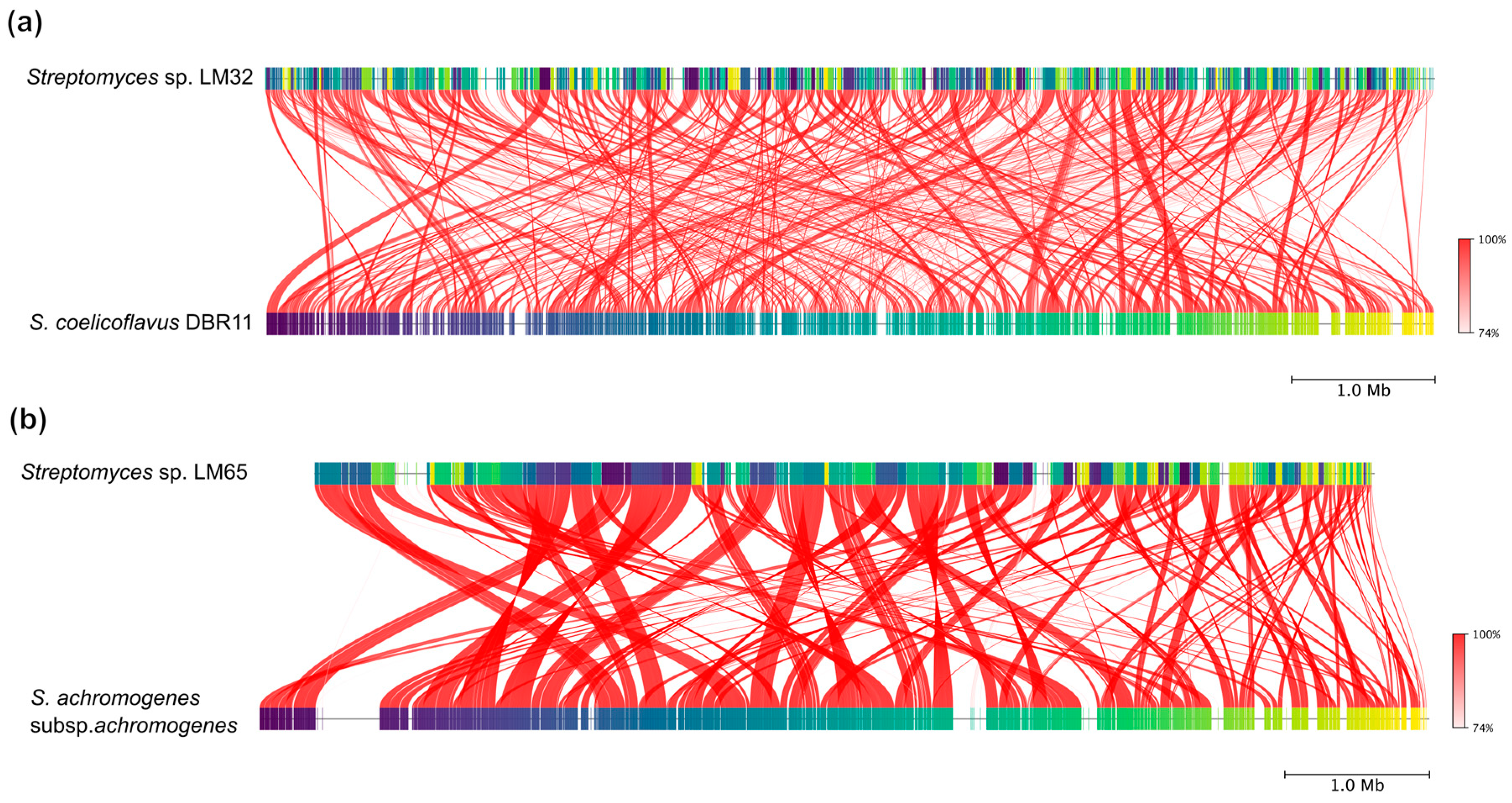
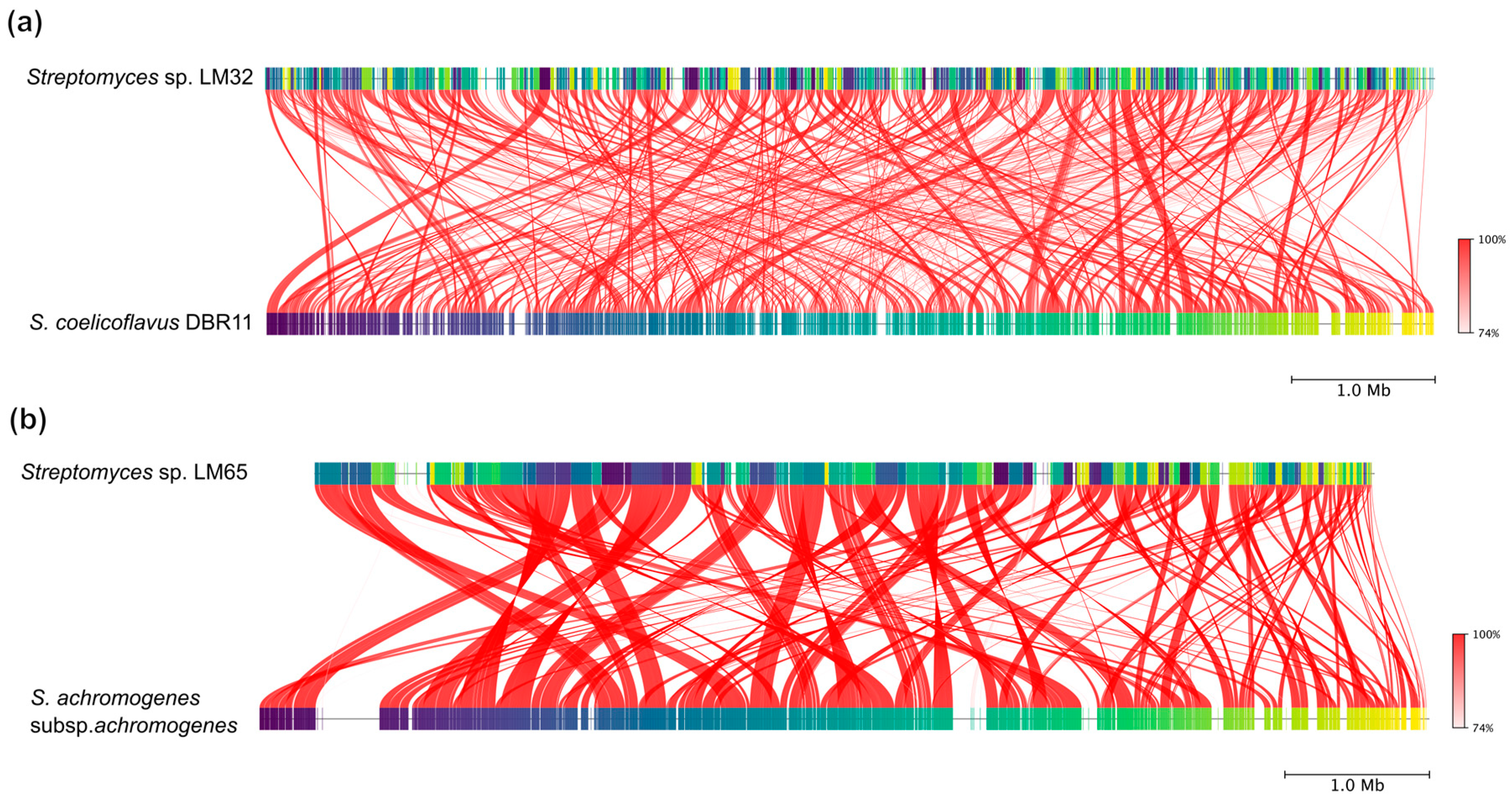
3.2. Phylogenomic Analysis of Streptomyces Genomes
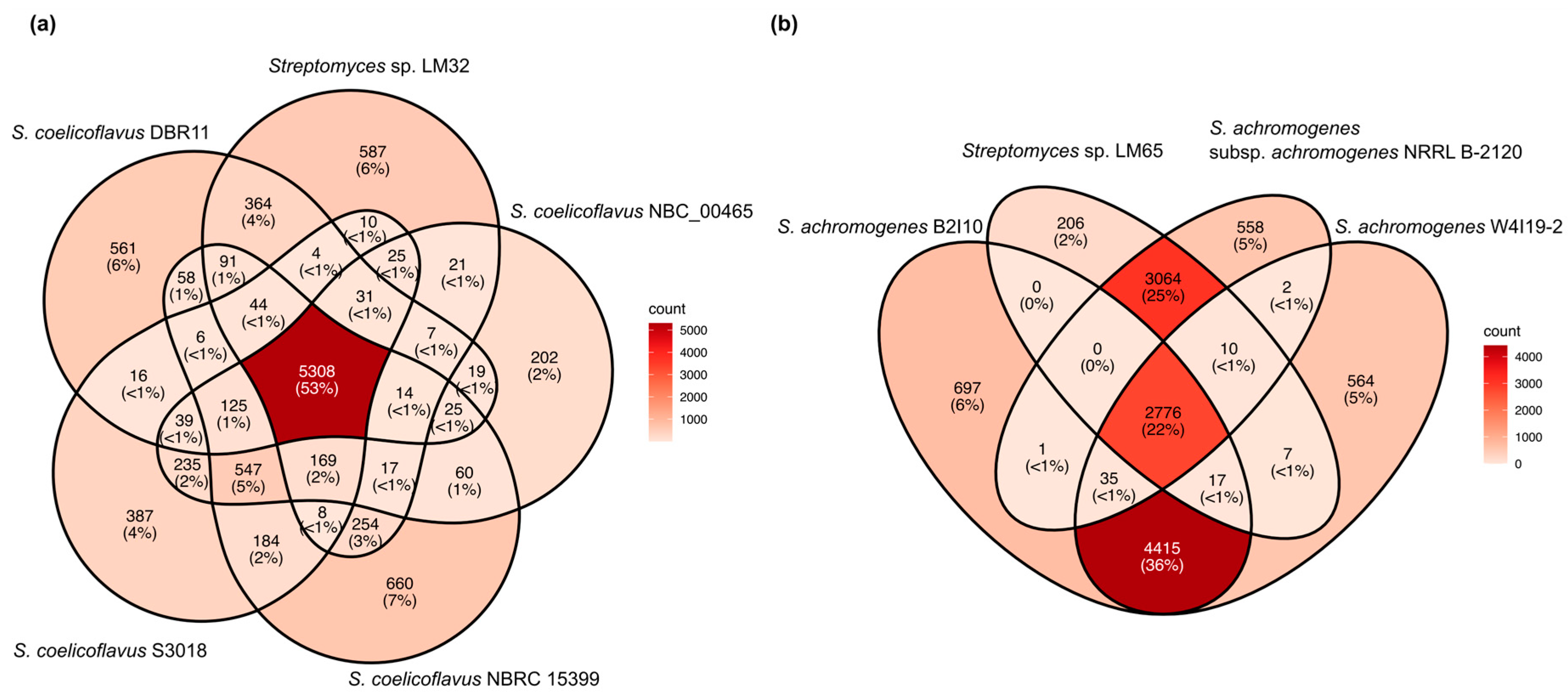
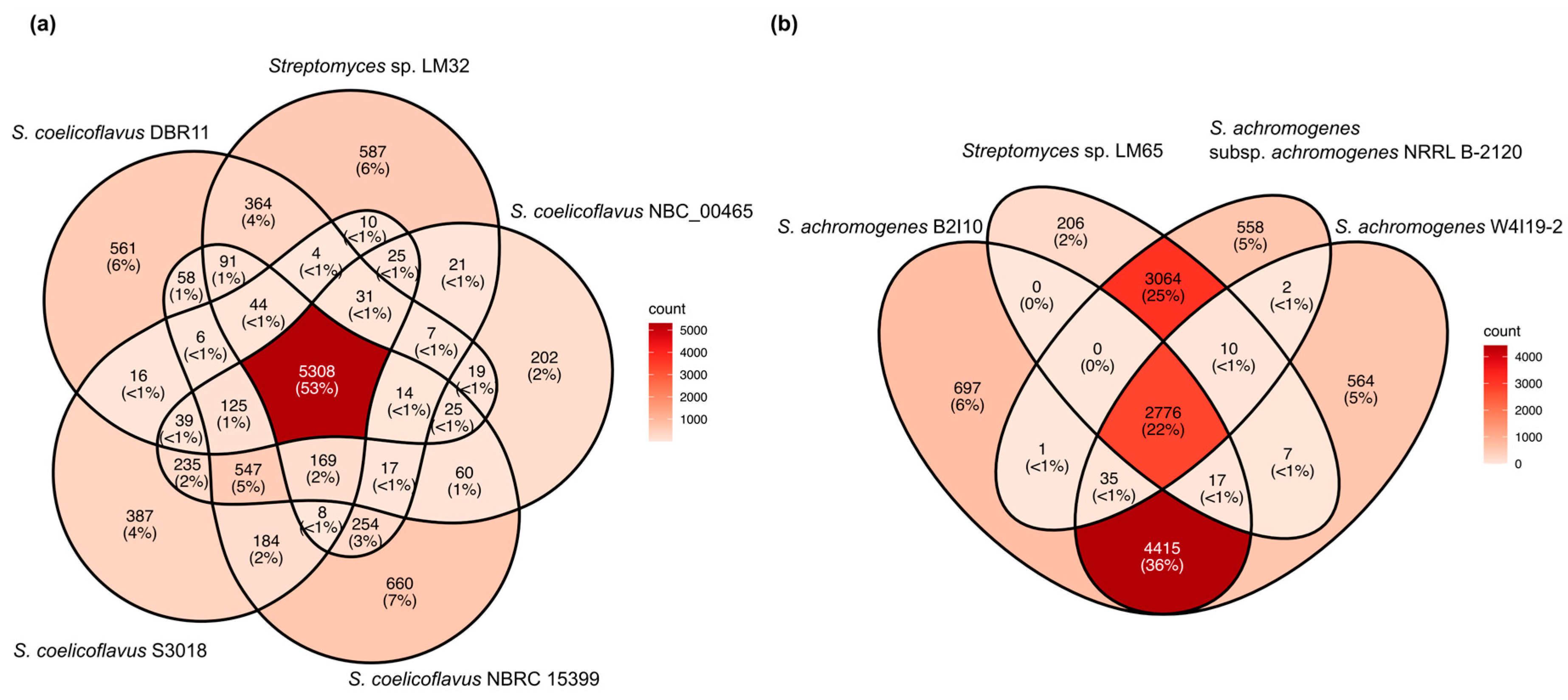
3.3. Pan- and Core-Genome
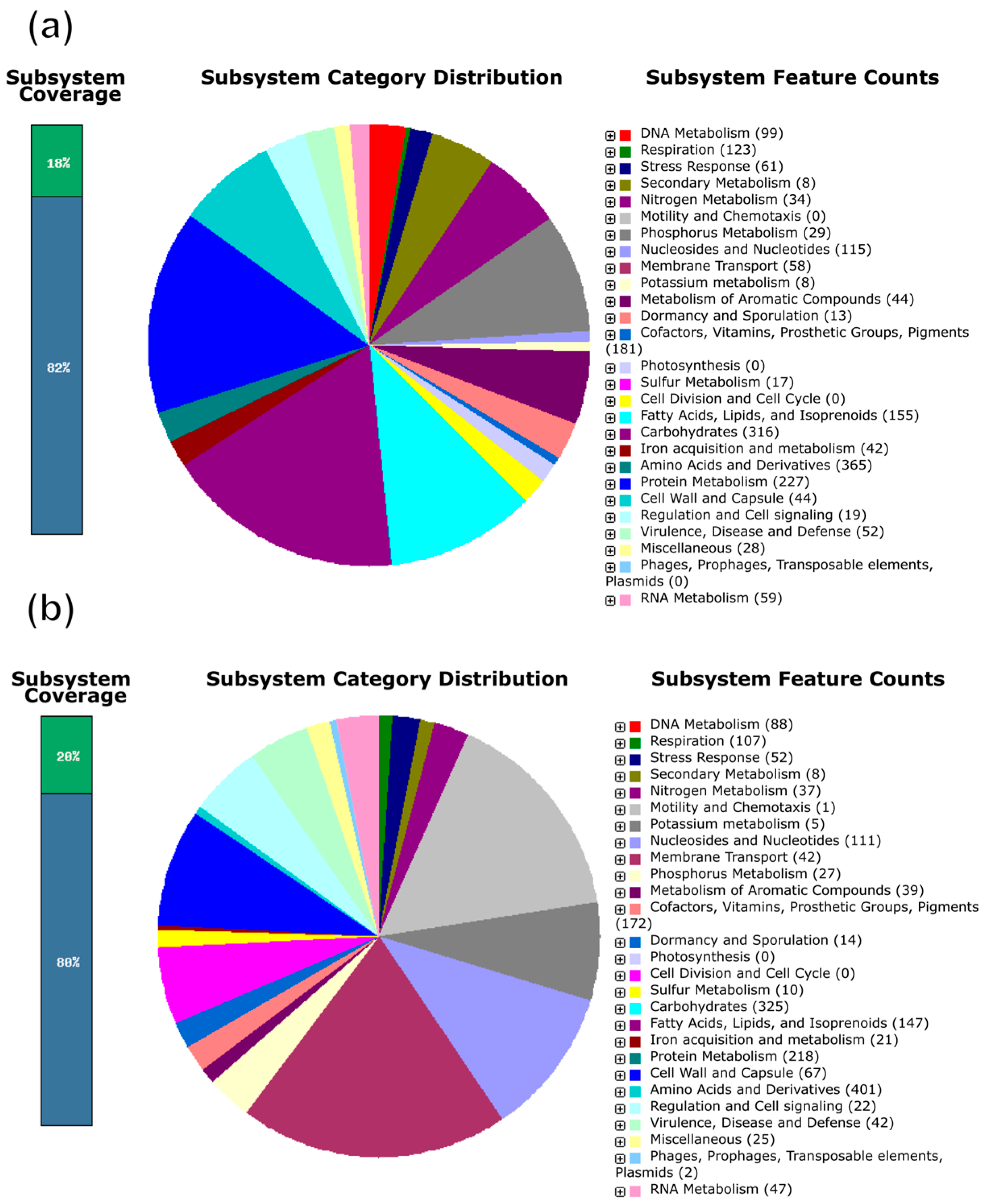
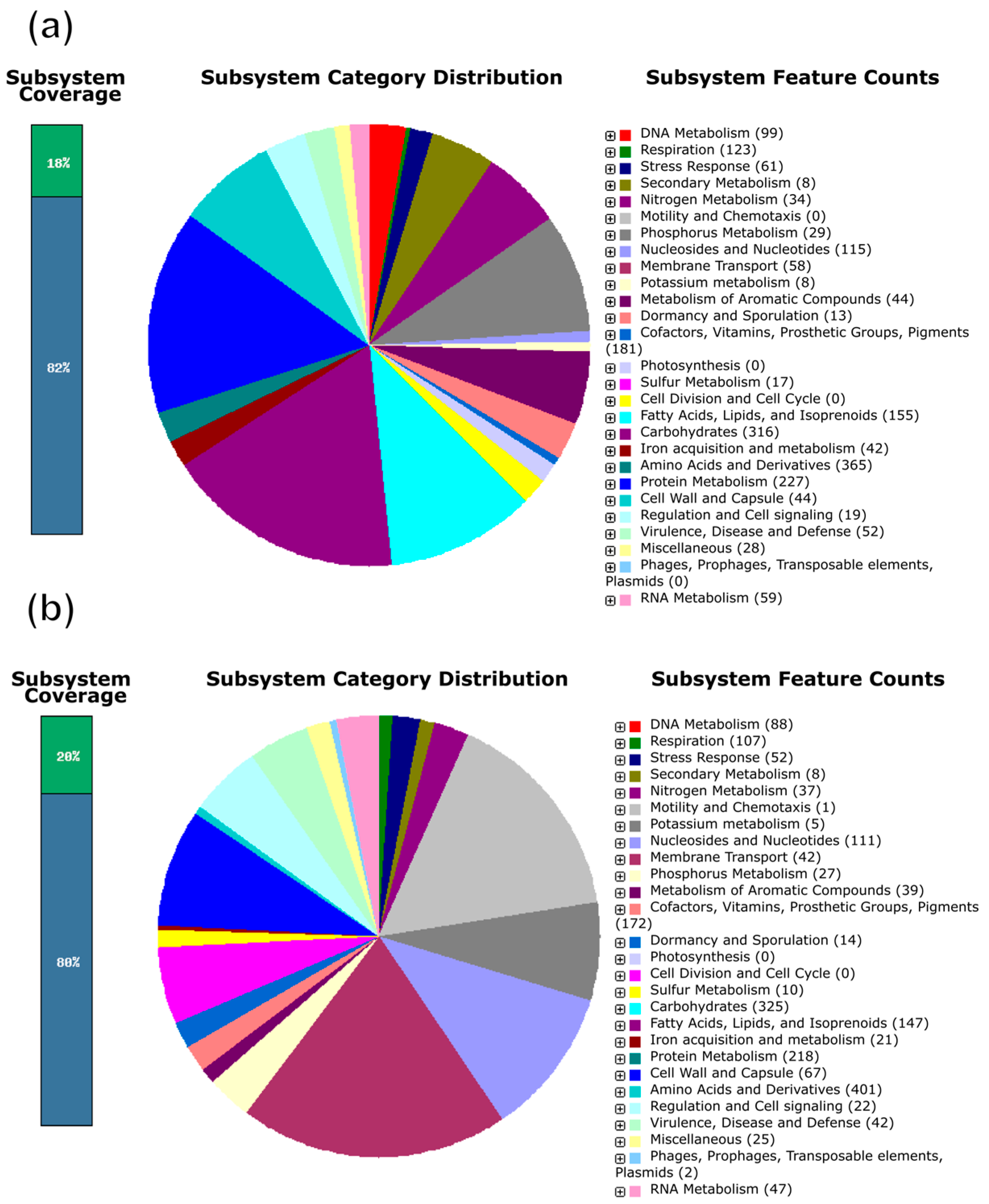
3.4. Gene Functional Annotation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumawat, K.C.; Razdan, N.; Saharan, K. Rhizospheric Microbiome: Bio-Based Emerging Strategies for Sustainable Agriculture Development and Future Perspectives. Microbiol. Res. 2022, 254, 126901. [Google Scholar] [CrossRef] [PubMed]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial Features of Plant Growth-Promoting Rhizobacteria for Improving Plant Growth and Health in Challenging Conditions: A Methodical Review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef]
- SADER Vino Mexicano Igual a Excelencia. Available online: https://www.gob.mx/agricultura/articulos/vino-mexicano-igual-a-excelencia?idiom=es (accessed on 16 February 2024).
- Bettenfeld, P.; Cadena i Canals, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.E.; Trouvelot, S. The Microbiota of the Grapevine Holobiont: A Key Component of Plant Health. J. Adv. Res. 2022, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Yadav, K.; Yadav, A.; Yadav, K. Regulation of Plant-Microbe Interactions in the Rhizosphere for Plant Growth and Metabolism: Role of Soil Phosphorus. In Phosphorus Soils Plants; IntechOpen: London, UK, 2024. [Google Scholar] [CrossRef]
- Shanthi, V. Actinomycetes: Implications and Prospects in Sustainable Agriculture. In Biofertilizers: Study and Impact; Scrivener Publishing LLC: Beverly, MA, USA, 2021; pp. 335–370. [Google Scholar] [CrossRef]
- Nwachukwu, B.C.; Ayangbenro, A.S.; Babalola, O.O. Elucidating the Rhizosphere Associated Bacteria for Environmental Sustainability. Agriculture 2021, 11, 75. [Google Scholar] [CrossRef]
- Gamalero, E.; Bona, E.; Novello, G.; Boatti, L.; Mignone, F.; Massa, N.; Cesaro, P.; Berta, G.; Lingua, G. Discovering the Bacteriome of Vitis vinifera Cv. Pinot Noir in a Conventionally Managed Vineyard. Sci. Rep. 2020, 10, 6453. [Google Scholar] [CrossRef]
- Kanjanamaneesathian, P.; Shah, A.; Ridgway, H.; Jones, E.E. Diversity and Bioactivity of Endophytic Actinobacteria Associated with Grapevines. Curr. Microbiol. 2022, 79, 390. [Google Scholar] [CrossRef]
- Santamaría, R.I.; Martínez-Carrasco, A.; Martín, J.; Tormo, J.R.; Pérez-Victoria, I.; González, I.; Genilloud, O.; Reyes, F.; Díaz, M. Grapevine Xylem Sap Is a Potent Elicitor of Antibiotic Production in Streptomyces spp. Antibiotics 2022, 11, 672. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Win, H.Y.; Islam, M.T.; Von Tiedemann, A.; Schüffler, A.; Laatsch, H. Khatmiamycin, a Motility Inhibitor and Zoosporicide against the Grapevine Downy Mildew Pathogen Plasmopara viticola from Streptomyces sp. ANK313. J. Antibiot. 2011, 64, 655–659. [Google Scholar] [CrossRef] [PubMed]
- del Pilar Martínez-Diz, M.; Díaz-Losada, E.; Andrés-Sodupe, M.; Bujanda, R.; Maldonado-González, M.M.; Ojeda, S.; Yacoub, A.; Rey, P.; Gramaje, D. Field Evaluation of Biocontrol Agents against Black-Foot and Petri Diseases of Grapevine. Pest Manag. Sci. 2021, 77, 697–708. [Google Scholar] [CrossRef]
- Buzón-Durán, L.; Sánchez-Hernández, E.; Sánchez-Báscones, M.; García-González, M.C.; Hernández-Navarro, S.; Correa-Guimarães, A.; Martín-Ramos, P. A Coating Based on Bioactive Compounds from Streptomyces spp. and Chitosan Oligomers to Control Botrytis cinerea Preserves the Quality and Improves the Shelf Life of Table Grapes. Plants 2023, 12, 577. [Google Scholar] [CrossRef]
- Rashid, M.H.; Krehenbrink, M.; Akhtar, M.S. Nitrogen-Fixing Plant-Microbe Symbioses. Sustain. Agric. Rev. 2015, 15, 193–234. [Google Scholar] [CrossRef]
- Negi, S.; Kumar, P.; Kumar, J.; Singh, A.; Dubey, R.C. Indigenous Nitrogen Fixing Microbes Engineer Rhizosphere and Enhance Nutrient Availability and Plant Growth. In Rhizosphere Engineering; Academic Press: Cambridge, MA, USA, 2022; pp. 19–43. [Google Scholar] [CrossRef]
- Pullan, S.T.; Chandra, G.; Bibb, M.J.; Merrick, M. Genome-Wide Analysis of the Role of GlnR in Streptomyces venezuelae Provides New Insights into Global Nitrogen Regulation in Actinomycetes. BMC Genom. 2011, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yan, R.; Fu, Y.; Wang, X.; Zhang, J.; Xiang, W. Antifungal, Plant Growth-Promoting, and Genomic Properties of an Endophytic Actinobacterium Streptomyces sp. NEAU-S7GS2. Front. Microbiol. 2019, 10, 2077. [Google Scholar] [CrossRef]
- Nonthakaew, N.; Panbangred, W.; Songnuan, W.; Intra, B. Plant Growth-Promoting Properties of Streptomyces spp. Isolates and Their Impact on Mung Bean Plantlets’ Rhizosphere Microbiome. Front. Microbiol. 2022, 13, 967415. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Ramírez, Z.Y.; González-Escobedo, R.; Avila-Quezada, G.D.; Ramírez-Sánchez, O.; Higareda-Alvear, V.M.; Zapata-Chávez, E.; Borrego-Loya, A.; Muñoz-Castellanos, L.N. Exploring Microbial Rhizosphere Communities in Asymptomatic and Symptomatic Apple Trees Using Amplicon Sequencing and Shotgun Metagenomics. Agronomy 2024, 14, 357. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S.; Krueger, F.; Segonds-Pichon, A.; Biggins, L.; Krueger, C.; Wingett, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. Genome Analysis QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]
- Wickham, H. Data Analysis; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 189–201. [Google Scholar] [CrossRef]
- Wilkins, D. Gggenes: Draw Gene Arrow Maps in “Ggplot2”. Available online: https://rdrr.io/cran/gggenes/ (accessed on 10 January 2024).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 6 January 2024).
- Muñoz-Ramírez, Z.Y.; Mendez-Tenorio, A.; Kato, I.; Bravo, M.M.; Rizzato, C.; Thorell, K.; Torres, R.; Aviles-Jimenez, F.; Camorlinga, M.; Canzian, F.; et al. Whole Genome Sequence and Phylogenetic Analysis Show Helicobacter pylori strains from Latin America Have Followed a Unique Evolution Pathway. Front. Cell Infect. Microbiol. 2017, 7, 245288. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Li, W.H. Mathematical Model for Studying Genetic Variation in Terms of Restriction Endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A Web Server for Prokaryotic Species Circumscription Based on Pairwise Genome Comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y. pyGenomeViz: A Genome Visualization Python Package for Comparative Genomics. Available online: https://github.com/moshi4/pyGenomeViz (accessed on 10 January 2024).
- Tonkin-Hill, G.; MacAlasdair, N.; Ruis, C.; Weimann, A.; Horesh, G.; Lees, J.A.; Gladstone, R.A.; Lo, S.; Beaudoin, C.; Floto, R.A.; et al. Producing Polished Prokaryotic Pangenomes with the Panaroo Pipeline. Genome Biol. 2020, 21, 1–21. [Google Scholar] [CrossRef]
- Gao, C.H.; Yu, G.; Cai, P. GgVennDiagram: An Intuitive, Easy-to-Use, and Highly Customizable R Package to Generate Venn Diagram. Front. Genet. 2021, 12, 706907. [Google Scholar] [CrossRef] [PubMed]
- Bury-Moné, S.; Thibessard, A.; Lioy, V.S.; Leblond, P. Dynamics of the Streptomyces chromosome: Chance and necessity. Trends in Genetics 2023, 39, 873–887. [Google Scholar] [CrossRef]
- Nikolaidis, M.; Hesketh, A.; Frangou, N.; Mossialos, D.; Van de Peer, Y.; Oliver, S.G.; Amoutzias, G.D. A Panoramic View of the Genomic Landscape of the Genus Streptomyces. Microb. Genom. 2023, 9, 001028. [Google Scholar] [CrossRef]
- Ju, K.S.; Gao, J.; Doroghazi, J.R.; Wang, K.K.A.; Thibodeaux, C.J.; Li, S.; Metzger, E.; Fudala, J.; Su, J.; Zhang, J.K.; et al. Discovery of Phosphonic Acid Natural Products by Mining the Genomes of 10,000 Actinomycetes. Proc. Natl. Acad. Sci. USA 2015, 112, 12175–12180. [Google Scholar] [CrossRef]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome Analysis of Multiple Pathogenic Isolates of Streptococcus agalactiae: Implications for the Microbial “Pan-Genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, G.; Jiménez-Hernández, N.; Peris-Bondia, F.; Moya, A.; Latorre, A. Legionella pneumophila Pangenome Reveals Strain-Specific Virulence Factors. BMC Genom. 2010, 11, 181. [Google Scholar] [CrossRef] [PubMed]
- Sawers, R.G.; Fischer, M.; Falke, D. Anaerobic Nitrate Respiration in the Aerobe Streptomyces coelicolor A3(2): Helping Maintain a Proton Gradient during Dormancy. Environ. Microbiol. Rep. 2019, 11, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Raghava Rao, K.V.; Mani, P.; Satyanarayana, B.; Raghava Rao, T. Purification and Structural Elucidation of Three Bioactive Compounds Isolated from Streptomyces coelicoflavus BC 01 and Their Biological Activity. 3 Biotech 2017, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Shaaban, M.I.; Abdel Bar, F.M.; El-Mahdy, A.M.; Shokralla, S. Quorum Sensing Inhibiting Activity of Streptomyces coelicoflavus Isolated from Soil. Front. Microbiol. 2016, 7, 188534. [Google Scholar] [CrossRef] [PubMed]
- Mon, W.P.; Jantaharn, P.; Boonlue, S.; McCloskey, S.; Kanokmedhakul, S.; Mongkolthanaruk, W. Enzymatic Degradation of Azo Bonds and Other Functional Groups on Commercial Silk Dyes by Streptomyces coelicoflavus CS-29. Environ. Nat. Resour. J. 2022, 20, 19–28. [Google Scholar] [CrossRef]
- Waheeb, M.S.; Elkhatib, W.F.; Yassien, M.A.; Hassouna, N.A. Production of Cellulase by Soil Isolated Streptomyces sp. Arch. Pharm. Sci. Ain Shams Univ. 2021, 5, 225–233. [Google Scholar] [CrossRef]
- Kaweewan, I.; Ohnishi-Kameyama, M.; Kodani, S. Isolation of a New Antibacterial Peptide Achromosin from Streptomyces achromogenes subsp. achromogenes Based on Genome Mining. J. Antibiot. 2016, 70, 208–211. [Google Scholar] [CrossRef]
- Tangjitjaroenkun, J.; Pluempanupat, W.; Tangchitcharoenkhul, R.; Yahayo, W.; Supabphol, R. Antibacterial, Antioxidant, Cytotoxic Effects and GC-MS Analysis of Mangrove-Derived Streptomyces achromogenes TCH4 Extract. Arch. Biol. Sci. 2021, 73, 223–235. [Google Scholar] [CrossRef]
- Chávez-Avila, S.; Valencia-Marin, M.F.; Guzmán-Guzmán, P.; Kumar, A.; Babalola, O.O.; Orozco-Mosqueda, M.d.C.; de los Santos-Villalobos, S.; Santoyo, G. Deciphering the Antifungal and Plant Growth-Stimulating Traits of the Stress-Tolerant Streptomyces achromogenes subsp. achromogenes strain UMAF16, a Bacterium Isolated from Soils Affected by Underground Fires. Biocatal. Agric. Biotechnol. 2023, 53, 102859. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes Benefaction Role in Soil and Plant Health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Prasanna, S.L. Streptomyces. In Beneficial Microbes in Agro-Ecology: Bacteria and Fungi; Academic Press: Cambridge, MA, USA, 2020; pp. 55–71. [Google Scholar] [CrossRef]
- Fischer, M.; Alderson, J.; Van Keulen, G.; White, J.; Sawers, R.G. The Obligate Aerobe Streptomyces coelicolor A3(2) Synthesizes Three Active Respiratory Nitrate Reductases. Microbiology 2010, 156, 3166–3179. [Google Scholar] [CrossRef]
- Stolz, J.F.; Basu, P. Evolution of Nitrate Reductase: Molecular and Structural Variations on a Common Function. ChemBioChem 2002, 3, 198–206. [Google Scholar] [CrossRef]
- Fisher, S.H.; Wray, L.V. Regulation of Glutamine Synthetase in Streptomyces coelicolor. J. Bacteriol. 1989, 171, 2378–2383. [Google Scholar] [CrossRef]
- Crack, J.C.; Munnoch, J.; Dodd, E.L.; Knowles, F.; Al Bassam, M.M.; Kamali, S.; Holland, A.A.; Cramer, S.P.; Hamilton, C.J.; Johnson, M.K.; et al. NsrR from Streptomyces coelicolor Is a Nitric Oxide-Sensing [4Fe-4S] Cluster Protein with a Specialized Regulatory Function. J. Biol. Chem. 2015, 290, 12689–12704. [Google Scholar] [CrossRef]
- Reuther, J.; Wohlleben, W. Nitrogen Metabolism in Streptomyces coelicolor: Transcriptional and Post-Translational Regulation. J. Mol. Microbiol. Biotechnol. 2007, 12, 139–146. [Google Scholar] [CrossRef]
- Mast, Y.; Wohlleben, W.; Reuther, J. Regulation of Nitrogen Assimilation in Streptomyces and Other Actinobacteria. Regulation of Nitrogen Assimilation in Streptomycetes and Other Actinobacteria; Caister Academic Press: Norfolk, UK, 2011; pp. 125–136. [Google Scholar]
- Navone, L.; Macagno, J.P.; Licona-Cassani, C.; Marcellin, E.; Nielsen, L.K.; Gramajo, H.; Rodriguez, E. AllR Controls the Expression of Streptomyces coelicolor Allantoin Pathway Genes. Appl. Environ. Microbiol. 2015, 81, 6649–6659. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Robles, M.; Santos-Beneit, F.; Rodríguez-García, A.; Martín, J.F. Analysis of the Pho Regulon in Streptomyces Tsukubaensis. Microbiol. Res. 2017, 205, 80–87. [Google Scholar] [CrossRef]
- Fernández-Martínez, L.T.; Santos-Beneit, F.; Martín, J.F. Is PhoR-PhoP Partner Fidelity Strict? PhoR Is Required for the Activation of the Pho Regulon in Streptomyces coelicolor. Mol. Genet. Genom. 2012, 287, 565–573. [Google Scholar] [CrossRef]
- Wang, L.; Yang, H.; Wu, M.; Zhang, J.; Zhang, H.; Mao, Z.; Chen, X. Integrative Transcriptome and Proteome Revealed High-Yielding Mechanisms of Epsilon-Poly-L-Lysine by Streptomyces albulus. Front. Microbiol. 2023, 14, 1123050. [Google Scholar] [CrossRef] [PubMed]
- Lindner, S.N.; Knebel, S.; Wesseling, H.; Schoberth, S.M.; Wendisch, V.F. Exopolyphosphatases PPX1 and PPX2 from Corynebacterium glutamicum. Appl. Environ. Microbiol. 2009, 75, 3161–3170. [Google Scholar] [CrossRef] [PubMed]
- Krysenko, S. Impact of Nitrogen-Containing Compounds on Secondary Metabolism in Streptomyces spp.—A Source of Metabolic Engineering Strategies. SynBio 2023, 1, 204–225. [Google Scholar] [CrossRef]
- Tucker, N.P.; Hicks, M.G.; Clarke, T.A.; Crack, J.C.; Chandra, G.; Le Brun, N.E.; Dixon, R.; Hutchings, M.I. The Transcriptional Repressor Protein NsrR Senses Nitric Oxide Directly via a [2Fe-2S] Cluster. PLoS ONE 2008, 3, e3623. [Google Scholar] [CrossRef]
- Chen, J.; Hu, L.; Chen, N.; Jia, R.; Ma, Q.; Wang, Y. The Biocontrol and Plant Growth-Promoting Properties of Streptomyces alfalfae XN-04 Revealed by Functional and Genomic Analysis. Front. Microbiol. 2021, 12, 745766. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Characteristics | LM32 | LM65 |
|---|---|---|
| Genome size (Mb) | 8.1 | 7.3 |
| Contigs > 500 bp | 112 | 92 |
| G + C content (%) | 72.14 | 71 |
| N50 | 150,500 | 164,678 |
| L50 | 18 | 16 |
| CDS | 7251 | 6440 |
| rRNA genes | 5,1,2 (5S,16S,23S) | 2,1,1 (5S,16S,23S) |
| tRNA genes | 83 | 99 |
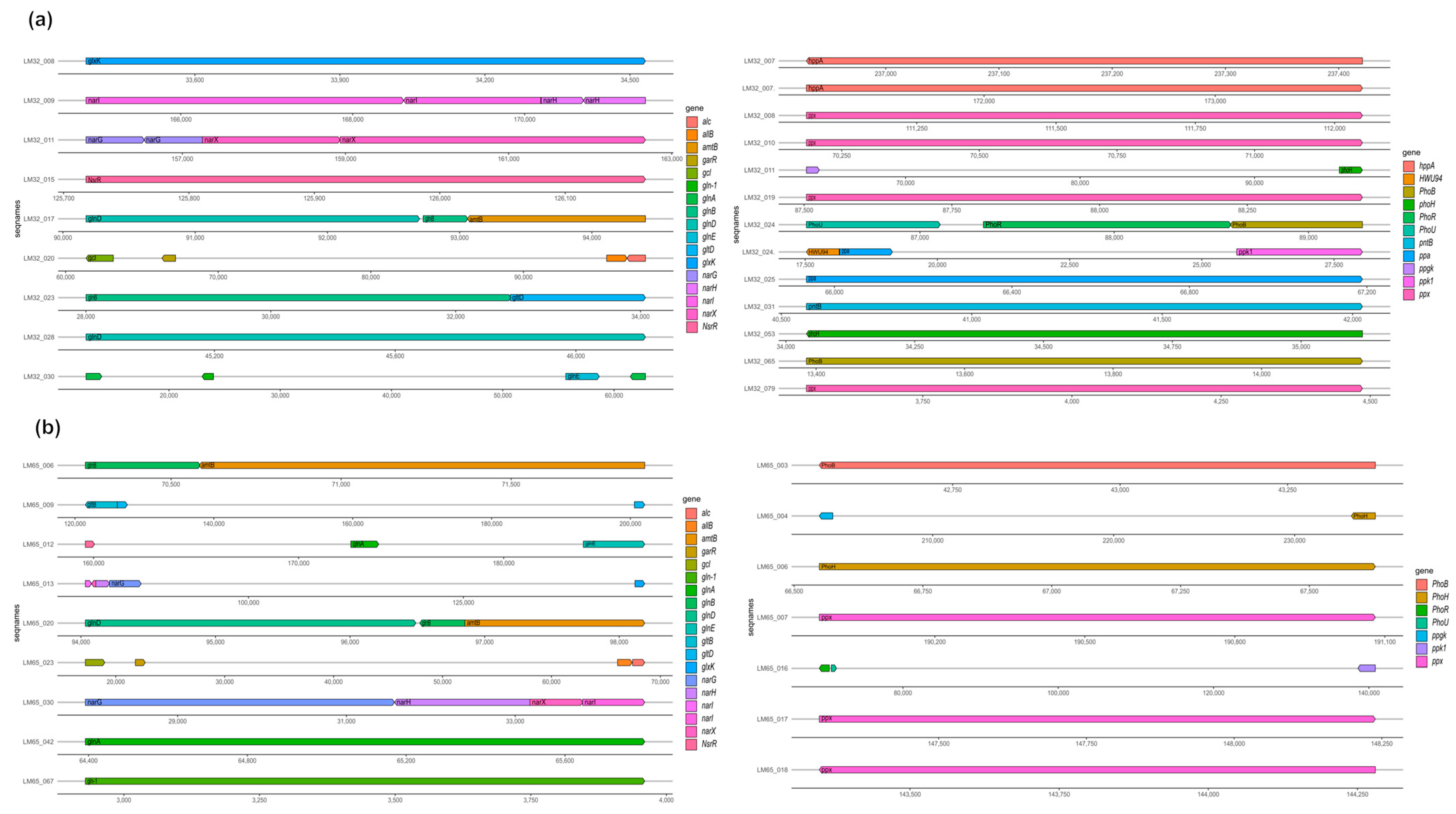
| Nitrogen Metabolism | Gene Name | Gene Annotation | Reference |
|---|---|---|---|
| Denitrifying reductase gene clusters | narG | Respiratory nitrate reductase alpha chain | [55] |
| narI | Respiratory nitrate reductase gamma chain | ||
| narX | Respiratory nitrate reductase delta chain | ||
| narH | Respiratory nitrate reductase beta chain | ||
| Nitrosative stress | NsrR | Nitrite-sensitive transcriptional repressor | [56] |
| Ammonia assimilation | gln-1 | Glutamine synthetase type II | [55,57,58] |
| gltD | Glutamate synthase [NADPH] small chain | ||
| glnB | Nitrogen regulatory protein P-II | ||
| gltB | Glutamate synthase [NADPH] large chain | ||
| glnD | [Protein-PII] uridylyltransferase | ||
| amtB | Ammonium transporter | ||
| glnE | Glutamate-ammonia-ligase adenylyltransferase | ||
| glnA | Glutamine synthetase type I | ||
| Nitrate and nitrite ammonification | narG | Respiratory nitrate reductase alpha chain | [43,54] |
| narX | Respiratory nitrate reductase delta chain | ||
| narH | Respiratory nitrate reductase beta chain | ||
| nasD | Nitrite reductase [NAD(P)H] large subunit | ||
| nasE | Nitrite reductase [NAD(P)H] small subunit | ||
| narI | Respiratory nitrate reductase gamma chain | ||
| alc | Allantoicase | [59] | |
| gcl | Glyoxylate carboligase | ||
| Allantoin utilization | glxK | Glycerate kinase | |
| allB | Allantoinase | ||
| garR | 2-hydroxy-3-oxopropionate reductase | ||
| Phosphorus metabolism | Gene name | Gene annotation | |
| High-affinity phosphate transporter and control of PHO regulon | phoU | Phosphate transport system regulatory protein | [60] |
| phoR | Phosphate regulon sensor protein | ||
| phoB | Phosphate regulon transcriptional regulatory protein | ||
| ppk1 | Polyphosphate kinase | ||
| Polyphosphate | ppgk | Polyphosphate glucokinase | [61] |
| ppx | Exopolyphosphatase | ||
| ppk1 | Polyphosphate kinase | ||
| Phosphate metabolism | phoH | Phosphate starvation-inducible protein PhoH, predicted ATPase | [61,62,63] |
| pitB | Probable low-affinity inorganic phosphate transporter | ||
| PhoU | Phosphate transport system regulatory protein | ||
| HWU94 | Phosphate transport regulator (distant homolog of PhoU) | ||
| phoL | Predicted ATPase related to phosphate starvation-inducible protein | ||
| ppx | Exopolyphosphatase | ||
| ppk1 | Polyphosphate kinase | ||
| hppA | Pyrophosphate-energized proton pump | ||
| phoB | Phosphate regulon transcriptional regulatory protein | ||
| pntB | NAD(P) transhydrogenase subunit beta | ||
| PhoR | Phosphate regulon sensor protein | ||
| ppa | Inorganic pyrophosphatase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montes-Montes, G.; González-Escobedo, R.; Muñoz-Castellanos, L.N.; Avila-Quezada, G.D.; Ramírez-Sánchez, O.; Borrego-Loya, A.; Ortiz-Aguirre, I.; Muñoz-Ramírez, Z.Y. Whole Genome Analysis of Streptomyces spp. Strains Isolated from the Rhizosphere of Vitis vinifera L. Reveals Their Role in Nitrogen and Phosphorus Metabolism. Nitrogen 2024, 5, 301-314. https://doi.org/10.3390/nitrogen5020020
Montes-Montes G, González-Escobedo R, Muñoz-Castellanos LN, Avila-Quezada GD, Ramírez-Sánchez O, Borrego-Loya A, Ortiz-Aguirre I, Muñoz-Ramírez ZY. Whole Genome Analysis of Streptomyces spp. Strains Isolated from the Rhizosphere of Vitis vinifera L. Reveals Their Role in Nitrogen and Phosphorus Metabolism. Nitrogen. 2024; 5(2):301-314. https://doi.org/10.3390/nitrogen5020020
Chicago/Turabian StyleMontes-Montes, Gustavo, Román González-Escobedo, Laila N. Muñoz-Castellanos, Graciela D. Avila-Quezada, Obed Ramírez-Sánchez, Alejandra Borrego-Loya, Ismael Ortiz-Aguirre, and Zilia Y. Muñoz-Ramírez. 2024. "Whole Genome Analysis of Streptomyces spp. Strains Isolated from the Rhizosphere of Vitis vinifera L. Reveals Their Role in Nitrogen and Phosphorus Metabolism" Nitrogen 5, no. 2: 301-314. https://doi.org/10.3390/nitrogen5020020