Fire Recurrence and Normalized Difference Vegetation Index (NDVI) Dynamics in Brazilian Savanna


Abstract
:1. Introduction
2. Materials and Methods
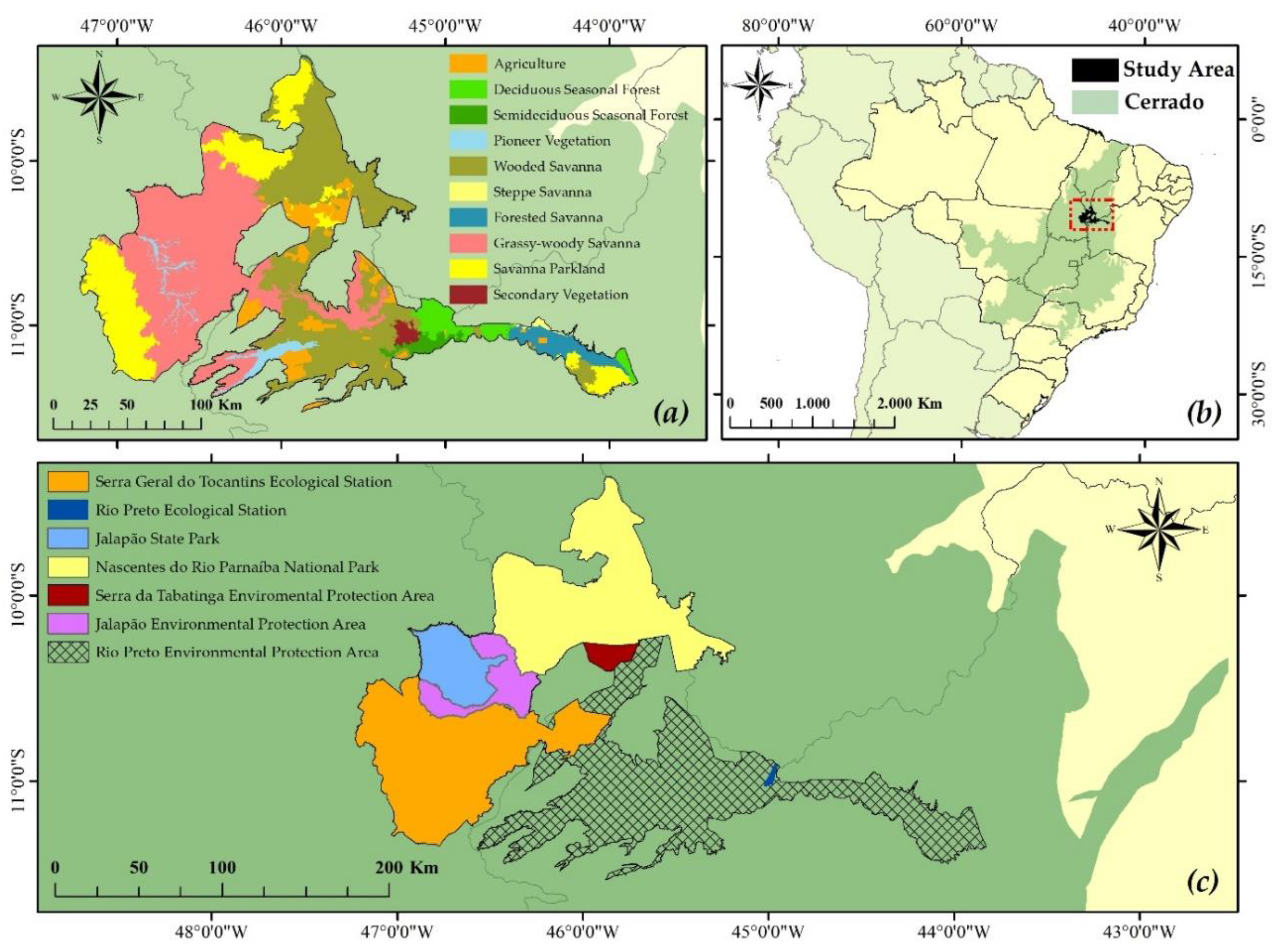
2.1. Study Area
2.2. Fire Recurrence
2.3. MODIS Time Series
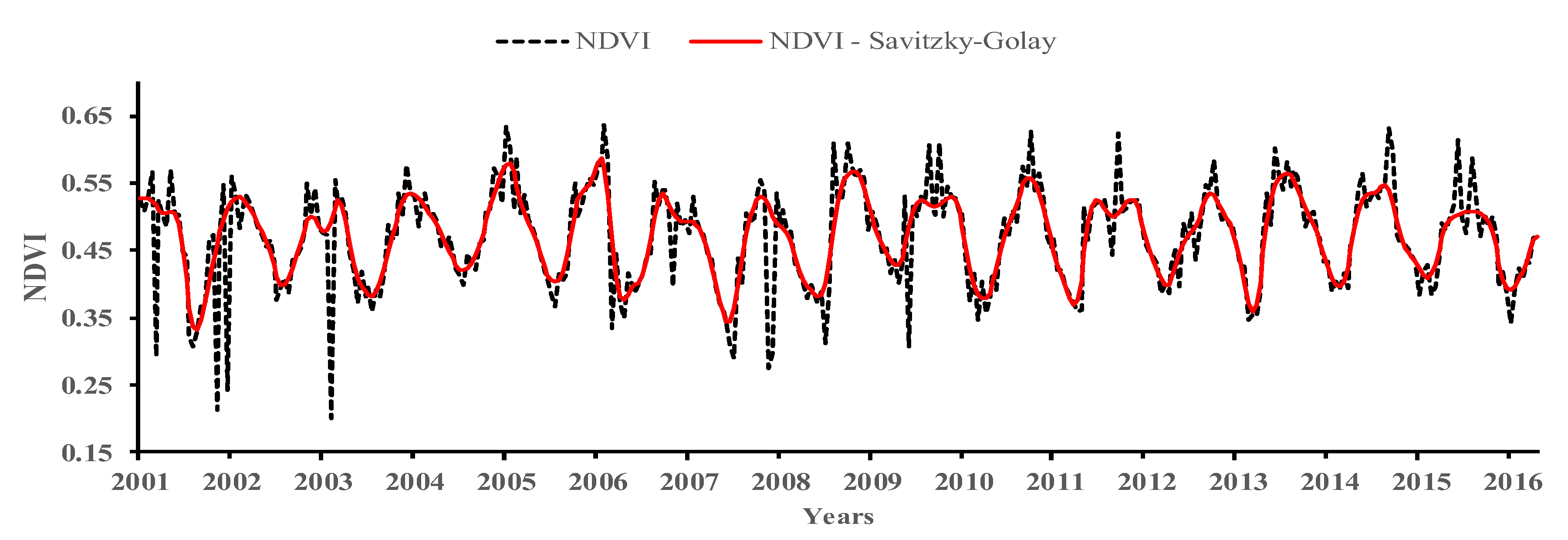
2.4. Vegetation Trend Dynamics
3. Results
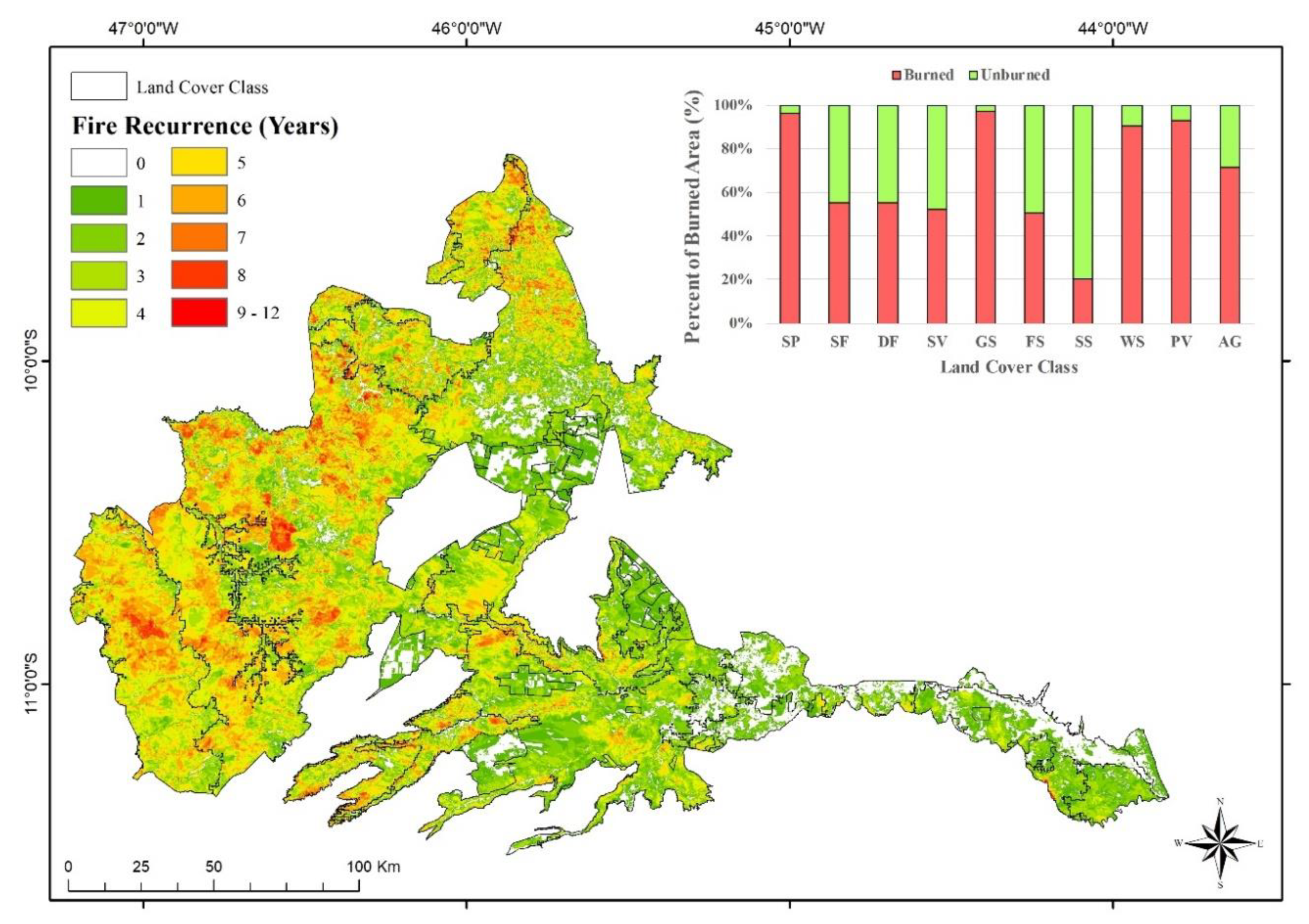
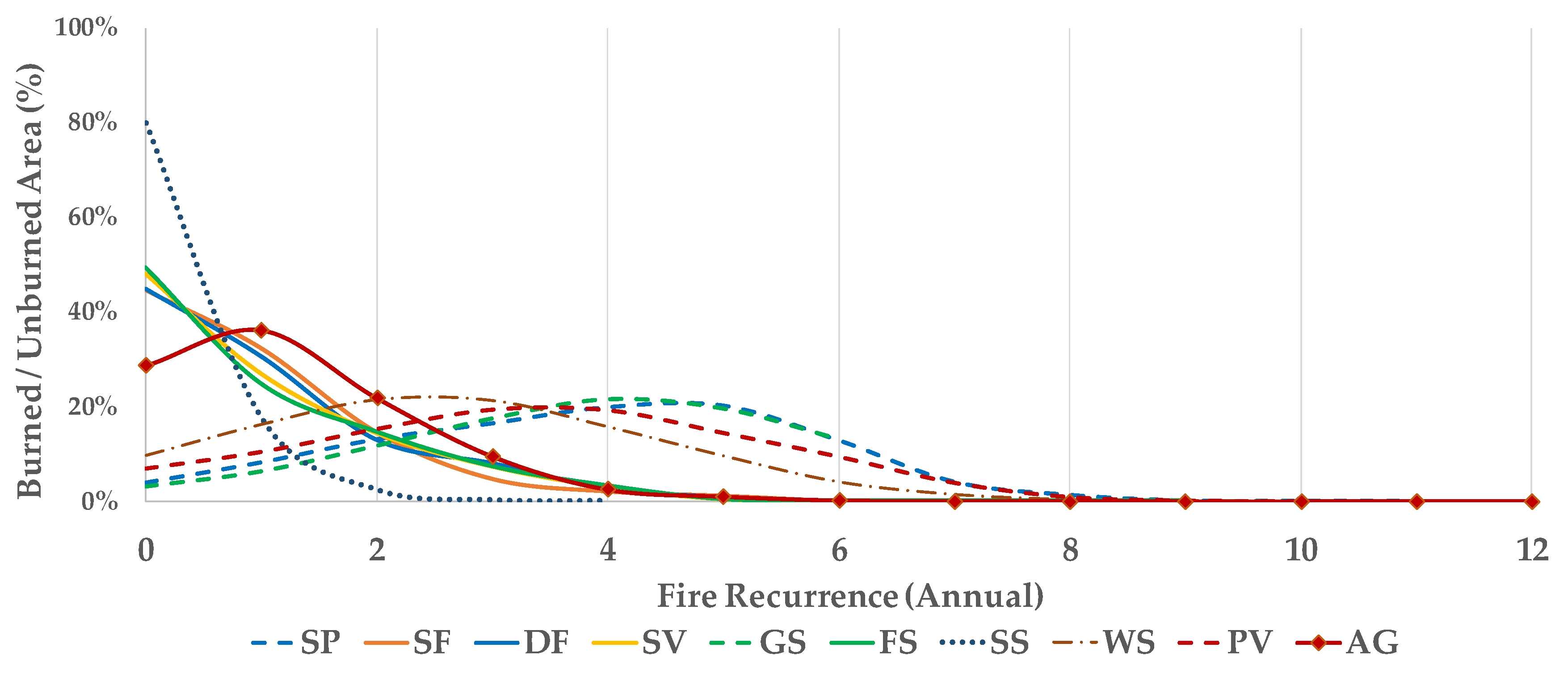
3.1. Fire Recurrence
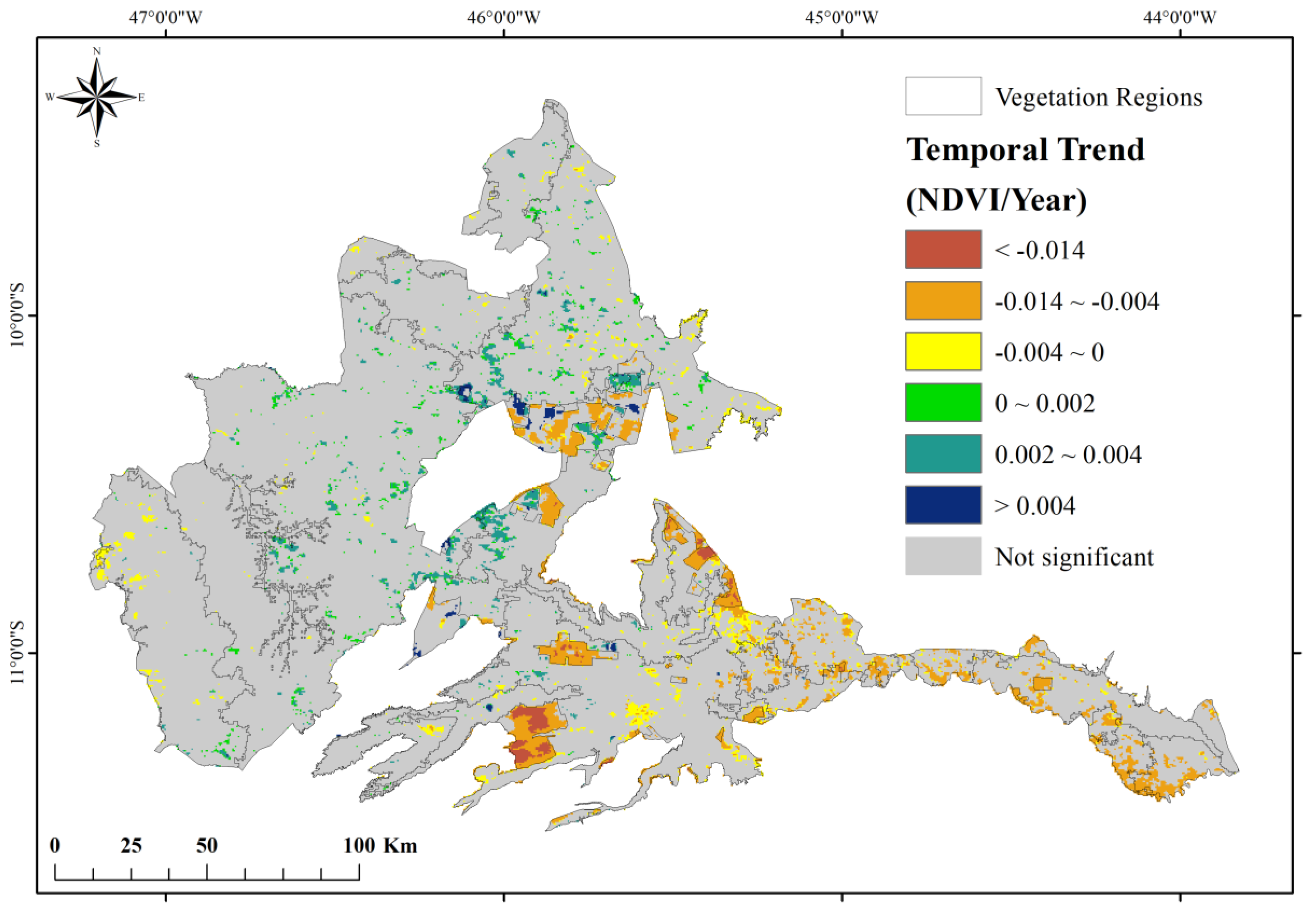
3.2. Spatiotemporal Changes in Vegetation
4. Discussion
4.1. Spatial Patterns of Fire Recurrence
4.2. Spatial Patterns of Vegetation Changes
4.3. Management Implications
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Bowman, D.M.J.S.; Balch, J.; Artaxo, P.; Bond, W.J.; Cochrane, M.A.; D’Antonio, C.M.; Defries, R.; Johnston, F.H.; Keeley, J.E.; Krawchuk, M.A.; et al. The human dimension of fire regimes on Earth. J. Biogeogr. 2011, 38, 2223–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archibald, S.; Lehmann, C.E.R.; Belcher, C.M.; Bond, W.J.; Bradstock, R.A.; Daniau, A.L.; Dexter, K.G.; Forrestel, E.J.; Greve, M.; He, T.; et al. Biological and geophysical feedbacks with fire in the Earth system. Environ. Res. Lett. 2018, 13. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bowman, D.M.J.S. What controls the distribution of tropical forest and savanna? Ecol. Lett. 2012, 15, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The Global Distribtuion of Ecosystems in a world without Fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Ribeiro, E. Fire and plant diversity at the global scale. Glob. Ecol. Biogeogr. 2017, 26, 889–897. [Google Scholar] [CrossRef]
- Oluwole, F.A.; Sambo, J.M.; Sikhalazo, D. Long-term effects of different burning frequencies on the dry savannah grassland in South Africa. African J. Agric. Res. 2008, 3, 147–153. [Google Scholar]
- Uys, R.G.; Bond, W.J.; Everson, T.M. The effect of different fire regimes on plant diversity in southern African grasslands. Biol. Conserv. 2004, 118, 489–499. [Google Scholar] [CrossRef]
- Williams, R.J.; Cook, G.D.; Gill, A.M.; Moore, P.H.R. Fire regime, fire intensity and tree survival in a tropical savanna in northern Australia. Austral Ecol. 1999, 24, 50–59. [Google Scholar] [CrossRef]
- Hoffmann, W.A. Fire and Population Dynamics of Woody Plants in a Neotropical Savanna: Matrix Model Projections. Ecology 1999, 80, 1354. [Google Scholar] [CrossRef]
- Schwieder, M.; Leitão, P.J.; da Cunha Bustamante, M.M.; Ferreira, L.G.; Rabe, A.; Hostert, P. Mapping Brazilian savanna vegetation gradients with Landsat time series. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 361–370. [Google Scholar] [CrossRef]
- De Dantas, V.L.; Batalha, M.A.; Pausas, J.G. Fire drives functional thresholds on the savanna – forest transition. Ecology 2013, 94, 2454–2463. [Google Scholar] [CrossRef]
- Coutinho, L.M. Ecological Effects of Fire in Brazilian Cerrado. In Ecology of Tropical Savannas; Huntley, B.J., Walker, B.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; pp. 273–291. [Google Scholar]
- Medeiros, M.B.; Miranda, H.S. Post-fire resprouting and mortality in Cerrado woody plant species over a three-year period. Edinburgh J. Bot. 2008, 65, 53–68. [Google Scholar] [CrossRef]
- Durigan, G.; Ratter, J.A. The need for a consistent fire policy for Cerrado conservation. J. Appl. Ecol. 2016, 53, 11–15. [Google Scholar] [CrossRef]
- Moreira, A.G. Effects of fire protection on savanna structure in central Brazil. J. Biogeogr. 2000, 27, 1021–1029. [Google Scholar] [CrossRef]
- De Oliveira, S.N.; de Carvalho Júnior, O.A.; Gomes, R.A.T.; Guimarães, R.F.; McManus, C.M. Landscape-fragmentation change due to recent agricultural expansion in the Brazilian Savanna, Western Bahia, Brazil. Reg. Environ. Chang. 2017, 17, 411–423. [Google Scholar] [CrossRef]
- Da Silva, J.M.C.; Bates, J.M. Biogeographic Patterns and Conservation in the South American Cerrado: A Tropical Savanna Hotspot. Bioscience 2002, 52, 225–233. [Google Scholar] [CrossRef]
- Klink, C.A.; Machado, R.B. Conservation of the Brazilian Cerrado. Conserv. Biol. 2005, 19, 707–713. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Schroeder, W.; Jackson, R.B. Positive feedbacks of fire, climate, and vegetation and the conversion of tropical savanna. Geophys. Res. Lett. 2002, 29, 9-1–9-4. [Google Scholar] [CrossRef]
- Vrieling, A.; De Leeuw, J.; Said, M.Y. Length of growing period over africa: Variability and trends from 30 years of NDVI time series. Remote Sens. 2013, 5, 982–1000. [Google Scholar] [CrossRef]
- Butt, B.; Turner, M.D.; Singh, A.; Brottem, L. Use of MODIS NDVI to evaluate changing latitudinal gradients of rangeland phenology in Sudano-Sahelian West Africa. Remote Sens. Environ. 2011, 115, 3367–3376. [Google Scholar] [CrossRef]
- Silva, F.B.; Shimabukuro, Y.E.; Aragão, L.E.O.C.; Anderson, L.O.; Pereira, G.; da Cardozo, F.S.; Arai, E. Large-scale heterogeneity of Amazonian phenology revealed from 26-year long AVHRR/NDVI time-series. Environ. Res. Lett. 2013, 8, 12. [Google Scholar] [CrossRef]
- Anderson, L.O.; Malhi, Y.; Aragão, L.E.O.C.; Ladle, R.; Arai, E.; Barbier, N.; Phillips, O. Remote sensing detection of droughts in Amazonian forest canopies. New Phytol. 2010, 187, 733–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koltunov, A.; Ustin, S.L.; Asner, G.P.; Fung, I. Selective logging changes forest phenology in the Brazilian Amazon: Evidence from MODIS image time series analysis. Remote Sens. Environ. 2009, 113, 2431–2440. [Google Scholar] [CrossRef]
- Zewdie, W.; Csaplovics, E.; Inostroza, L. Monitoring ecosystem dynamics in northwestern Ethiopia using NDVI and climate variables to assess long term trends in dryland vegetation variability. Appl. Geogr. 2017, 79, 167–178. [Google Scholar] [CrossRef]
- Fernandez-Manso, A.; Quintano, C.; Roberts, D.A. Burn severity influence on post-fire vegetation cover resilience from Landsat MESMA fraction images time series in Mediterranean forest ecosystems. Remote Sens. Environ. 2016, 184, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, G.; Bradstock, R.A.; Collins, L.; Penman, T.D.; Watson, P. Using MODIS data to analyse post-fire vegetation recovery in Australian eucalypt forests. J. Spat. Sci. 2014, 8596, 1–12. [Google Scholar] [CrossRef]
- Van Leeuwen, W.J.D.; Casady, G.M.; Neary, D.G.; Bautista, S.; Alloza, J.A.; Carmel, Y.; Wittenberg, L.; Malkinson, D.; Orr, B.J. Monitoring post-wildfire vegetation response with remotely sensed time-series data in Spain, USA and Israel. Int. J. Wildl. Fire 2010, 19, 75–93. [Google Scholar] [CrossRef]
- Corner, B.R.; Narayanan, R.M.; Reichenbach, S.E. Noise estimation in remote sensing imagery using data masking. Int. J. Remote Sens. 2003, 24, 689–702. [Google Scholar] [CrossRef]
- Forkel, M.; Carvalhais, N.; Verbesselt, J.; Mahecha, M.D.; Neigh, C.S.R.; Reichstein, M. Trend Change detection in NDVI time series: Effects of inter-annual variability and methodology. Remote Sens. 2013, 5, 2113–2144. [Google Scholar] [CrossRef]
- Katagis, T.; Gitas, I.Z.; Toukiloglou, P.; Veraverbeke, S.; Goossens, R. Trend analysis of medium- and coarse-resolution time series image data for burned area mapping in a Mediterranean ecosystem. Int. J. Wildl. Fire 2014, 23, 668–677. [Google Scholar] [CrossRef]
- Clemente, R.H.; Navarro Cerrillo, R.M.; Gitas, I.Z. Monitoring post-fire regeneration in Mediterranean ecosystems by employing multitemporal satellite imagery. Int. J. Wildl. Fire 2009, 18, 648–658. [Google Scholar] [CrossRef]
- Van Leeuwen, W.J.D. Monitoring the Effects of Forest Restoration Treatments on Post-Fire Vegetation Recovery with MODIS Multitemporal Data. Sensors 2008, 8, 2017–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polychronaki, A.; Gitas, I.Z.; Minchella, A. Monitoring post-fire vegetation recovery in the Mediterranean using SPOT and ERS imagery. Int. J. Wildl. Fire 2014, 23, 631–642. [Google Scholar] [CrossRef]
- Chen, X.; Vogelmann, J.E.; Rollins, M.; Ohlen, D.; Key, C.H.; Yang, L.; Huang, C.; Shi, H. Detecting post-fire burn severity and vegetation recovery using multitemporal remote sensing spectral indices and field-collected composite burn index data in a ponderosa pine forest. Int. J. Remote Sens. 2011, 32, 7905–7927. [Google Scholar] [CrossRef]
- Mishra, N.B.; Crews, K.A.; Neeti, N.; Meyer, T.; Young, K.R. MODIS derived vegetation greenness trends in African Savanna: Deconstructing and localizing the role of changing moisture availability, fire regime and anthropogenic impact. Remote Sens. Environ. 2015, 169, 192–204. [Google Scholar] [CrossRef]
- Jacquin, A.; Sheeren, D.; Lacombe, J.P. Vegetation cover degradation assessment in Madagascar savanna based on trend analysis of MODIS NDVI time series. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, 3–10. [Google Scholar] [CrossRef]
- Schmidt, M.; Lucas, R.; Bunting, P.; Verbesselt, J.; Armston, J. Multi-resolution time series imagery for forest disturbance and regrowth monitoring in Queensland, Australia. Remote Sens. Environ. 2015, 158, 156–168. [Google Scholar] [CrossRef] [Green Version]
- De Arruda, F.V.; De Sousa, D.G.; Teresa, F.B.; do Prado, V.H.M.; da Cunha, H.F.; Izzo, T.J. Trends and gaps of the scientific literature about the effects of fire on Brazilian Cerrado. Biota Neotrop. 2018, 18, 1–6. [Google Scholar] [CrossRef]
- Schmidt, I.S.; Fonseca, C.B.; Ferreira, M.C.; Sato, M.N. Implementação do Programa Piloto de Manejo Integrado do Fogo em três Unidades de Conservação do Cerrado. Biodiversidade Bras. 2016, 6, 55–70. [Google Scholar]
- Instituto Brasileiro de Geografia e Estatística (IBGE). Manual Técnico da Vegetação Brasileira; Instituto Brasileiro de Geografia e Estatística-IBGE: Rio de Janeiro, Brazil, 2012. [Google Scholar]
- De Santana, H.M.P.; Sano, E.E.; de Júniorde, M.P.O.; Lacerda, M.P.C.; Malaquias, J.V. Relação entre atributos físicos e químicos dos solos e a produtividade de capim dourado na Região do Jalapão, TO. Rev. Bras. Cienc. do Solo 2015, 39, 1172–1180. [Google Scholar] [CrossRef]
- Schmidt, I.B.; Figueiredo, I.B.; Borghetti, F.; Scariot, A. Produção e germinação de sementes de “capim dourado”, Syngonanthus nitens (Bong.) Ruhland (Eriocaulaceae): Implicações para o manejo. Acta Bot. Brasilica 2008, 22, 37–42. [Google Scholar] [CrossRef]
- Pereira Júnior, A.C.; Oliveira, S.L.J.; Pereira, J.M.C.; Turkman, M.A.A. Modelling fire frequency in a Cerrado savanna protected area. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.L.; Eloy, L.; Schmidt, I.B.; Barradas, A.C.S.; Santons, I.A. dos Manejo do Fogo em Veredas: Novas perspectivas a partir dos sistemas agrícolas tradicionais no Jalapão. Ambient. Soc. 2016, 19, 269–294. [Google Scholar] [CrossRef]
- Schmidt, I.B.; Sampaio, M.B.; Figueiredo, I.B.; Ticktin, T. Fogo e Artesanato de Capim-dourado no Jalapão: Usos Tradicionais e Consequências Ecológicas. Biodiversidade Bras. 2011, 1, 67–85. [Google Scholar]
- Instituto Nacional de Pesquisas Espaciais (INPE). Portal do Monitoramento de Queimadas e Incêndios. Available online: http://www.inpe.br/queimadas (accessed on 22 June 2015).
- Giglio, L.; Loboda, T.; Roy, D.P.; Quayle, B.; Justice, C.O. An active-fire based burned area mapping algorithm for the MODIS sensor. Remote Sens. Environ. 2009, 113, 408–420. [Google Scholar] [CrossRef]
- Arvidson, T.; Goward, S.N.; Gasch, J.; Williams, D. Landsat-7 long-term acquisition plan: Development and validation. Photogramm. Eng. Remote Sensing 2006, 72, 1137–1146. [Google Scholar] [CrossRef]
- Myneni, R.B.; Hall, F.G.; Sellers, P.J.; Marshak, A.L. The interpretation of spectral vegetation indexes. IEEE Trans. Geosci. Remote Sens. 1995, 33, 481–486. [Google Scholar] [CrossRef]
- Anderson, L.O. Biome-scale forest properties in Amazonia based on field and satellite observations. Remote Sens. 2012, 4, 1245–1271. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Vermote, E.F.; Masuoka, E.; Wolfe, R.E.; Saleous, N.; Roy, D.P.; Morisette, J.T. An overview of MODIS Land data processing and product status. Remote Sens. Environ. 2002, 83, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT-A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Abade, N.A.; de Carvalho Júnior, O.A.; Guimarães, R.F.; Oliveira, S.N. De Comparative Analysis of MODIS Time-Series Classification Using Support Vector Machines and Methods Based upon Distance and Similarity Measures in the Brazilian Cerrado-Caatinga Boundary. Remote Sens. 2015, 7, 12160–12191. [Google Scholar] [CrossRef]
- De Carvalho Júnior, O.A.; Guimarães, R.F.; Silva, C.; Gomes, R.A.T. Standardized Time-Series and Interannual Phenological Deviation: New Techniques for Burned-Area Detection Using Long-Term MODIS-NBR Dataset. Remote Sens. 2015, 7, 6950–6985. [Google Scholar] [CrossRef] [Green Version]
- Geng, L.; Ma, M.; Wang, X.; Yu, W.; Jia, S.; Wang, H. Comparison of Eight Techniques for Reconstructing Multi-Satellite Sensor Time-Series NDVI Data Sets in the Heihe River Basin, China. Remote Sens. 2014, 6, 2024–2049. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2014. [Google Scholar]
- Bai, J.; Perron, P. Computation and analysis of multiple structural change models. J. Appl. Econom. 2003, 18, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Mann, H.B. Nonparametric Tests Against Trend. Econometrica 1945, 13, 245. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank correlation methods, 2nd ed.; Hafner Publishing Co.: Oxford, UK, 1955. [Google Scholar]
- Ministério do Meio Ambiente (MMA). Mapeamento da Cobertura Vegetal do Bioma Cerrado, Edital Probio 02/2004, Projeto Executivo B.02.02.109, Relatório Final. Available online: http://mapas.mma.gov.br/ (accessed on 22 February 2018).
- Hoffmann, W.A.; Geiger, E.L.; Gotsch, S.G.; Rossatto, D.R.; Silva, L.C.R.; Lau, O.L.; Haridasan, M.; Franco, A.C. Ecological thresholds at the savanna-forest boundary: How plant traits, resources and fire govern the distribution of tropical biomes. Ecol. Lett. 2012, 15, 759–768. [Google Scholar] [CrossRef]
- Bradstock, R.A. A biogeographic model of fire regimes in Australia: Current and future implications. Glob. Ecol. Biogeogr. 2010, 19, 145–158. [Google Scholar] [CrossRef]
- Alves, D.B.; Pérez-Cabello, F. Multiple remote sensing data sources to assess spatio-temporal patterns of fire incidence over Campos Amazônicos Savanna Vegetation Enclave (Brazilian Amazon). Sci. Total Environ. 2017, 601–602, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.L.J.; Turkman, M.A.A.; Pereira, J.M.C. An analysis of fire frequency in tropical savannas of northern Australia, using a satellite-based fire atlas. Int. J. Wildl. Fire 2013, 22, 479–492. [Google Scholar] [CrossRef]
- Daldegan, G.A.; de Carvalho Júnior, O.A.; Guimarães, R.F.; Gomes, R.A.T.; Ribeiro, F.D.F.; McManus, C. Spatial patterns of fire recurrence using remote sensing and GIS in the Brazilian savanna: Serra do Tombador Nature Reserve, Brazil. Remote Sens. 2014, 6, 9873–9894. [Google Scholar] [CrossRef]
- Alvarado, S.T.; Silva, T.S.F.; Archibald, S. Management impacts on fire occurrence: A comparison of fire regimes of African and South American tropical savannas in different protected areas. J. Environ. Manage. 2018, 218, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Pivello, V.R. The use of fire in the cerrado and Amazonian rainforests of Brazil: Past and present. Fire Ecol. 2011, 7, 24–39. [Google Scholar] [CrossRef]
- Marlon, J.R.; Bartlein, P.J.; Daniau, A.L.; Harrison, S.P.; Maezumi, S.Y.; Power, M.J.; Tinner, W.; Vanniére, B. Global biomass burning: A synthesis and review of Holocene paleofire records and their controls. Quat. Sci. Rev. 2013, 65, 5–25. [Google Scholar] [CrossRef]
- Guiomar, N.; Godinho, S.; Fernandes, P.M.; Machado, R.; Neves, N.; Fernandes, J.P. Wildfire patterns and landscape changes in Mediterranean oak woodlands. Sci. Total Environ. 2015, 536, 338–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bragança, A.A. The Causes and Consequences of Agricultural Expansion in Matopiba. Rev. Bras. Econ. 2018, 72, 161–185. [Google Scholar] [CrossRef]
- Archibald, S. Managing the human component of fire regimes: Lessons from Africa. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150346. [Google Scholar] [CrossRef]
- Andela, N.; Morton, D.C.; Giglio, L.; Chen, Y.; van der Werf, G.R.; Kasibhatla, P.S.; DeFries, R.S.; Collatz, G.J.; Hantson, S.; Kloster, S.; et al. A human-driven decline in global burned area. Science 2017, 356, 1356–1362. [Google Scholar] [CrossRef] [Green Version]
- Rossiter, N.A.; Setterfield, S.A.; Douglas, M.M.; Hutley, L.B. Testing the grass-fire cycle: Alien grass invasion in the tropical savannas of northern Australia. Divers. Distrib. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Felderhof, L.; Gillieson, D. Comparison of fire patterns and fire frequency in two tropical savanna bioregions. Austral Ecol. 2006, 31, 736–746. [Google Scholar] [CrossRef]
- Schucknecht, A.; Erasmi, S.; Niemeyer, I.; Matschullat, J. Assessing vegetation variability and trends in north-eastern Brazil using AVHRR and MODIS NDVI time series. Eur. J. Remote Sens. 2013, 46, 40–59. [Google Scholar] [CrossRef]
- Li, S.; Potter, C. Vegetation regrowth trends in post forest fire ecosystems across North America from 2000 to 2010. Nat. Sci. 2012, 04, 755–770. [Google Scholar] [CrossRef]
- Schut, A.G.T.; Ivits, E.; Conijn, J.G.; Ten Brink, B.; Fensholt, R. Trends in global vegetation activity and climatic drivers indicate a decoupled response to climate change. PLoS ONE 2015, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Suepa, T.; Qi, J.; Lawawirojwong, S.; Messina, J.P. Understanding spatio-temporal variation of vegetation phenology and rainfall seasonality in the monsoon Southeast Asia. Environ. Res. 2016, 147, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Verbesselt, J.; Hyndman, R.; Newnham, G.; Culvenor, D. Detecting trend and seasonal changes in satellite image time series. Remote Sens. Environ. 2010, 114, 106–115. [Google Scholar] [CrossRef]
- Andersen, A.N.; Cook, G.D.; Corbett, L.K.; Douglas, M.M.; Eager, R.W.; Russell-Smith, J.; Setterfield, S.A.; Williams, R.J.; Woinarski, J.C.Z. Fire frequency and biodiversity conservation in Australian tropical savannas: Implications from the Kapalga fire experiment. Austral Ecol. 2005, 30, 155–167. [Google Scholar] [CrossRef]
- Higgins, S.I.; Bond, W.J.; February, E.C.; Bronn, A.; Euston-Brown, D.I.W.; Enslin, B.; Govender, N.; Rademan, L.; O’Regan, S.; Potgieter, A.L.F.; et al. Effects of four decades of fire manipulation on woody vegetation structure in savanna. Ecology 2007, 88, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Smit, I.P.J.; Asner, G.P.; Govender, N.; Kennedy-Bowdoin, T.; Knapp, D.E.; Jacobson, J. Effects of fire on woody vegetation structure in African savanna. Ecol. Appl. 2010, 20, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.; Chomitz, K.M. Effectiveness of strict vs. multiple use protected areas in reducing tropical forest fires: A global analysis using matching methods. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Welch, J.R.; Brondízio, E.S.; Hetrick, S.S.; Coimbra, C.E.A. Indigenous Burning as Conservation Practice: Neotropical Savanna Recovery amid Agribusiness Deforestation in Central Brazil. PLoS ONE 2013, 8, e81226. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Berardi, A.; Andrade, V.; Krahô, T.; Krahô, P.; Leonardos, O. Indigenous fire management in the cerrado of Brazil: The case of the Krahô of Tocantíns. Hum. Ecol. 2005, 33, 365–386. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Satellite | Sensor | Period Used | Total Scenes |
|---|---|---|---|
| Landsat 5 | TM | 2001–2011 | 11 |
| Landsat 7 | ETM+ | 2001–2003 | 3 |
| Landsat 7 | ETM+ (SLC-off) | 2003–2016 | 14 |
| Landsat 8 | OLI | 2013–2016 | 4 |
| Terra and Aqua | MODIS (MCD64A1 product) | 2012 | 1 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, N.C. Fire Recurrence and Normalized Difference Vegetation Index (NDVI) Dynamics in Brazilian Savanna. Fire 2019, 2, 1. https://doi.org/10.3390/fire2010001
Santana NC. Fire Recurrence and Normalized Difference Vegetation Index (NDVI) Dynamics in Brazilian Savanna. Fire. 2019; 2(1):1. https://doi.org/10.3390/fire2010001
Chicago/Turabian StyleSantana, Níckolas Castro. 2019. "Fire Recurrence and Normalized Difference Vegetation Index (NDVI) Dynamics in Brazilian Savanna" Fire 2, no. 1: 1. https://doi.org/10.3390/fire2010001
APA StyleSantana, N. C. (2019). Fire Recurrence and Normalized Difference Vegetation Index (NDVI) Dynamics in Brazilian Savanna. Fire, 2(1), 1. https://doi.org/10.3390/fire2010001