
Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau


Abstract
:1. Introduction
2. Materials and Methods
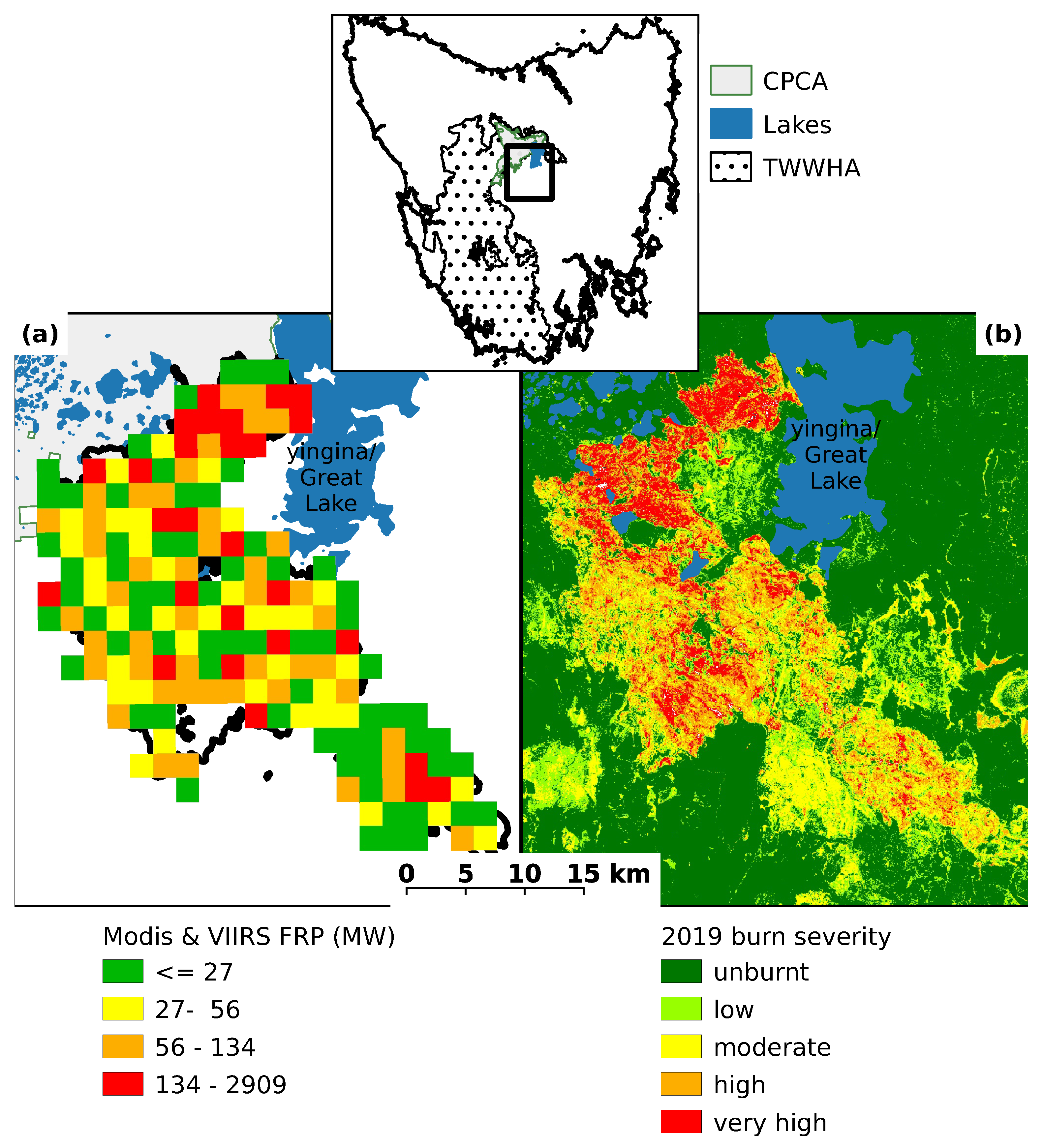
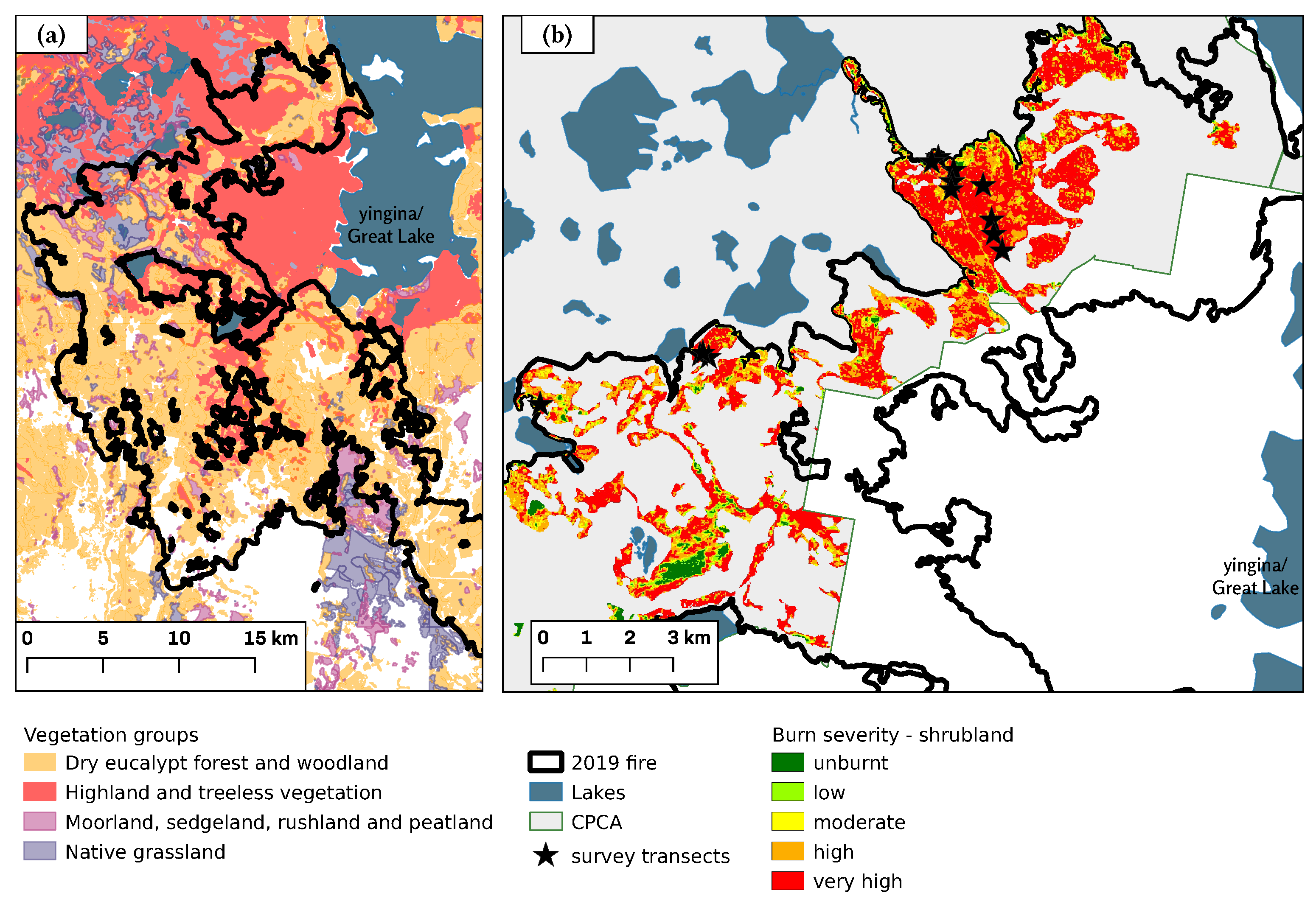
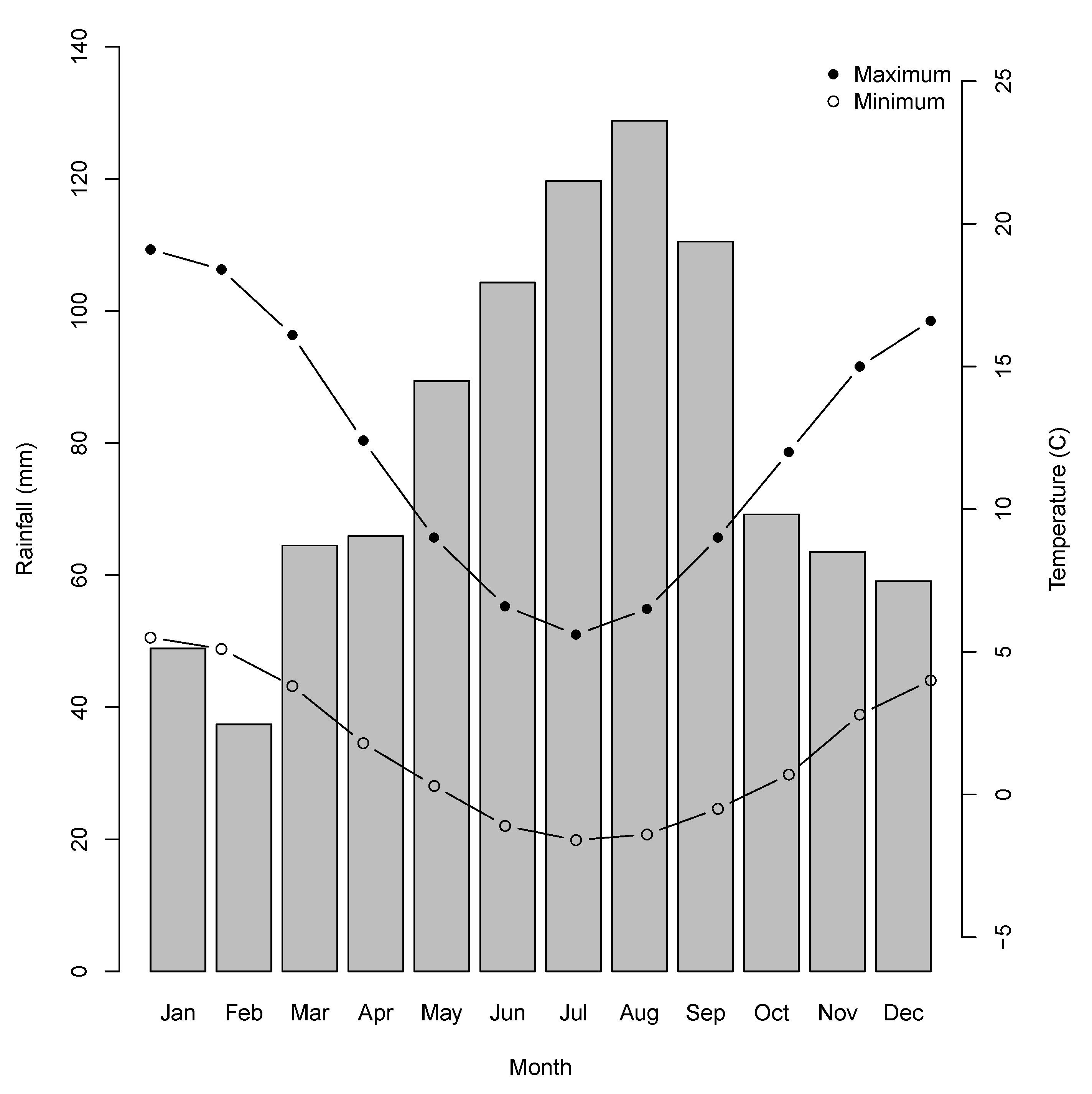
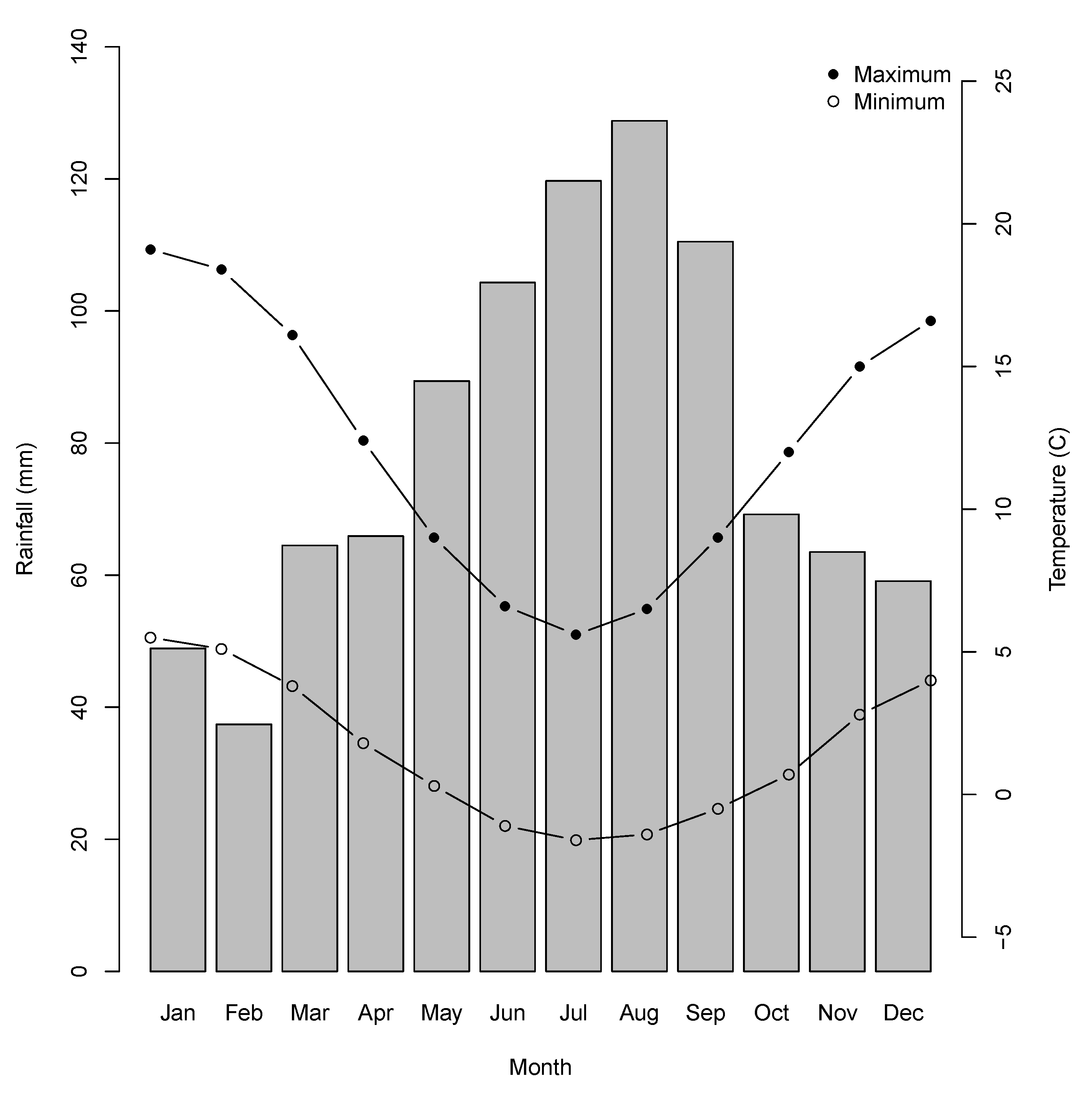
2.1. Study Region
2.2. Field Survey
2.3. Data Analysis
3. Results
3.1. Site Condition
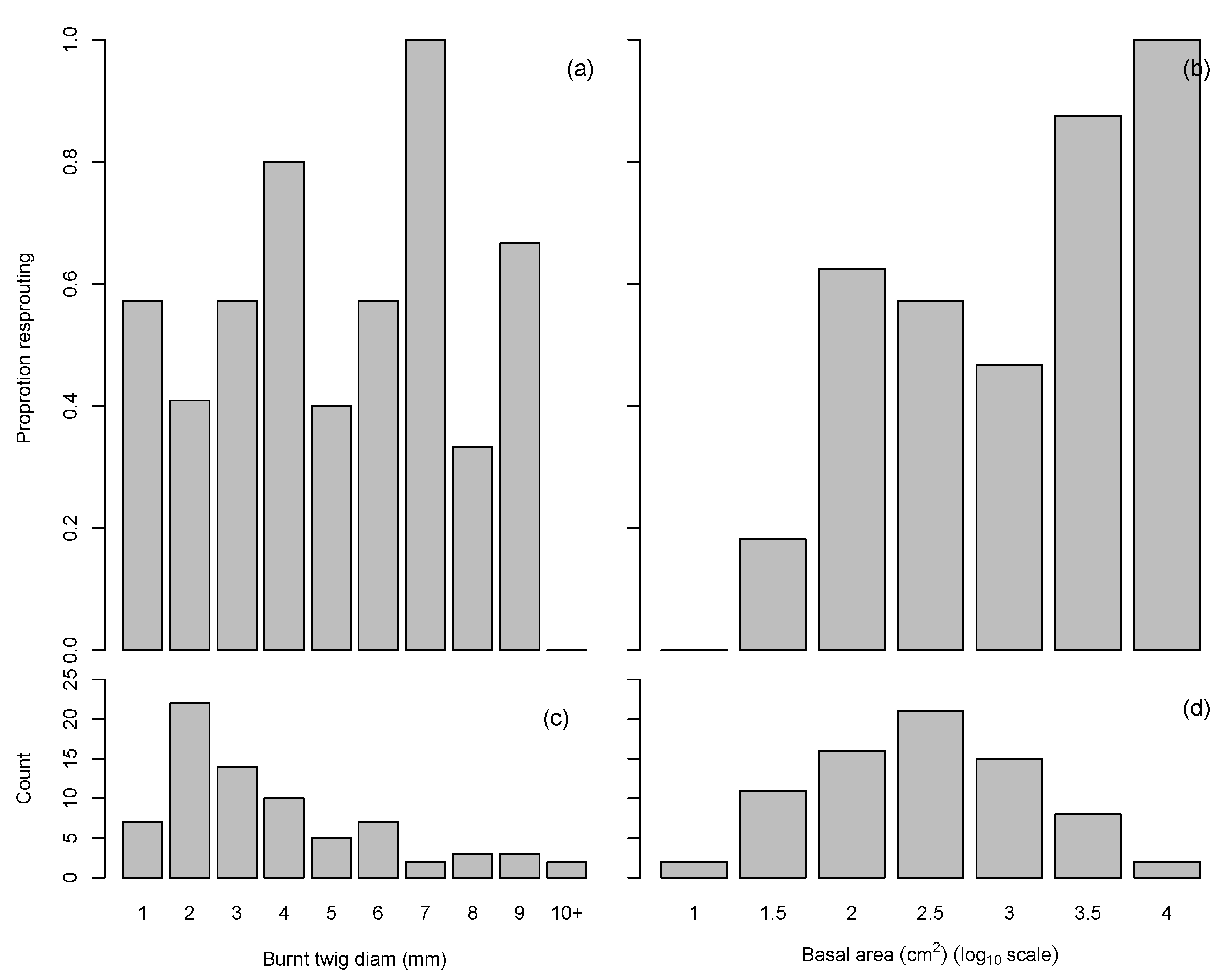
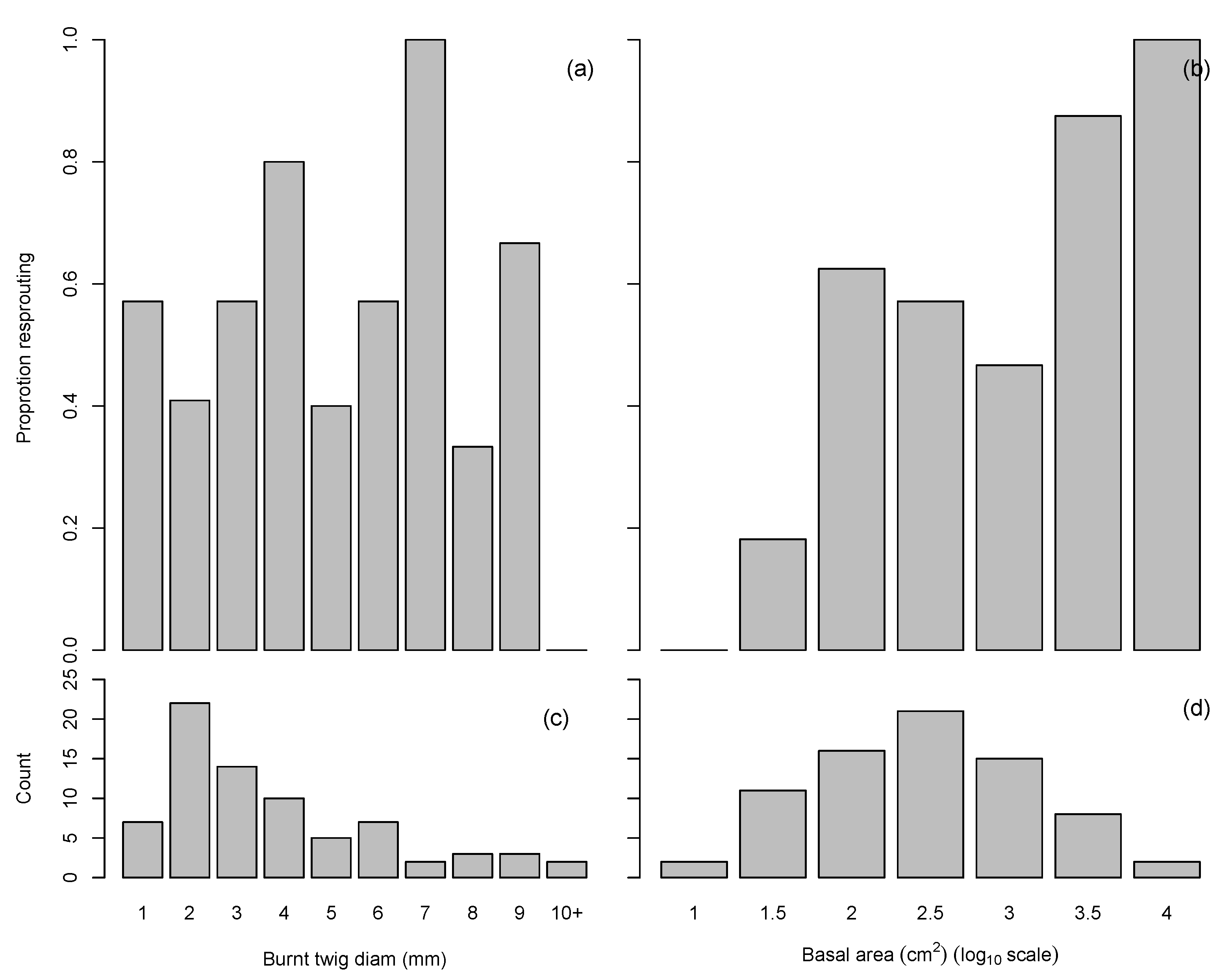
3.2. Plant Response
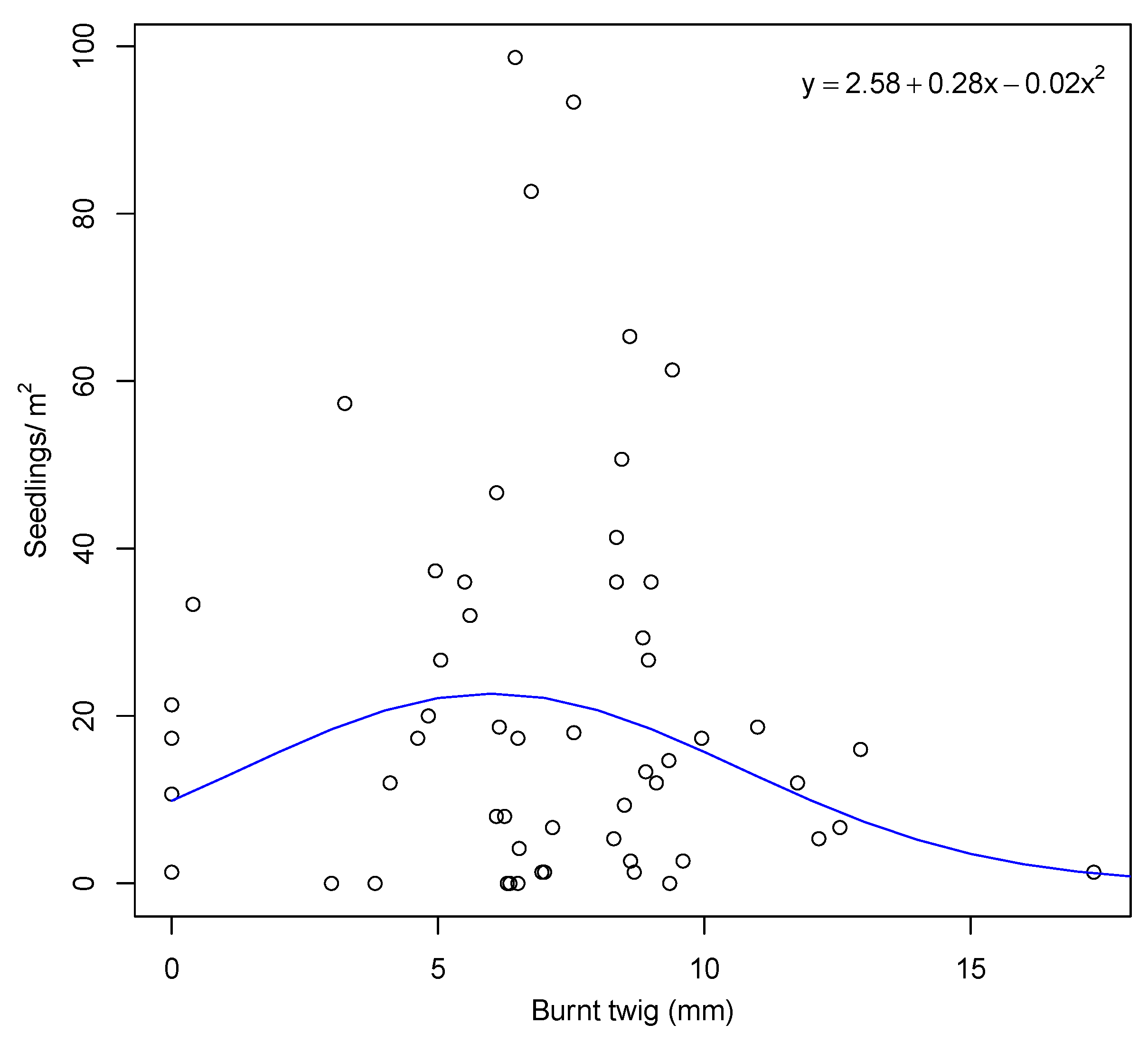
3.3. Seedling Recruitment
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal | Number of Plots | Number of Scats | ||
|---|---|---|---|---|
| Burnt | Unburnt | Burnt | Unburnt | |
| Macropod | 39 | 5 | 217 | 105 |
| Wombat | 3 | 0 | 16 | 0 |
| Deer | 9 | 0 | 18 | 0 |
| Rabbit | 1 | 0 | 21 | 0 |
| Total | 272 | 105 | ||

References
- McArthur, E.D.; Kitchen, S.G. Shrubland ecosystems: Importance, distinguishing characteristics, and dynamics. In Proceedings: Shrubland Dynamics–Fire and Water, Lubbock, TX, USA, 10–12 August 2004; Sosebee, R.E., Wester, D.B., Britton, C.M., McArthur, E.D., Kitchen, S.G., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2007; RMRS-P-47; pp. 3–10. [Google Scholar]
- McKell, C.M. Shrubs—A neglected resource of arid lands. Science 1975, 187, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Van Auken, O.W. Causes and consequences of woody plant encroachment into western North American grasslands. J. Environ. Manag. 2009, 90, 2931–2942. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Soliveres, S. Are shrubs really a sign of declining ecosystem function? Disentangling the myths and truths of woody encroachment in Australia. Aust. J. Bot. 2014, 62, 594–608. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Kröel-Dulay, G.; Ransijn, J.; Schmidt, I.K.; Beier, C.; De Angelis, P.; De Dato, G.; Dukes, J.S.; Emmett, B.; Estiarte, M.; Garadnai, J.; et al. Increased sensitivity to climate change in disturbed ecosystems. Nat. Commun. 2015, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, K.C. Shrub recruitment response to intensity and season of fire in a semi-arid woodland. J. Appl. Ecol. 1991, 28, 60–70. [Google Scholar] [CrossRef]
- Hodgkinson, K.C.; Harrington, G.N. The case for prescribed burning to control shrubs in eastern semi-arid woodlands. Aust. Rangel. J. 1985, 7, 64–74. [Google Scholar] [CrossRef]
- Joubert, D.F.; Smit, G.N.; Hoffman, M.T. The role of fire in preventing transitions from a grass dominated state to a bush thickened state in arid savannas. J. Arid Environ. 2012, 87, 1–7. [Google Scholar] [CrossRef]
- Lohmann, D.; Tietjen, B.; Blaum, N.; Joubert, D.F.; Jeltsch, F. Prescribed fire as a tool for managing shrub encroachment in semi-arid savanna rangelands. J. Arid Environ. 2014, 107, 49–56. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Camac, J.S.; Williams, R.J.; Wahren, C.H.; Hoffmann, A.A.; Vesk, P.A. Climatic warming strengthens a positive feedback between alpine shrubs and fire. Glob. Chang. Biol. 2017, 23, 3249–3258. [Google Scholar] [CrossRef] [Green Version]
- Holz, A.; Wood, S.W.; Veblen, T.T.; Bowman, D.M.J.S. Effects of high-severity fire drove the population collapse of the subalpine Tasmanian endemic conifer Athrotaxis cupressoides. Glob. Chang. Biol. 2015, 21, 445–458. [Google Scholar] [CrossRef]
- Flannigan, M.; Cantin, A.S.; De Groot, W.J.; Wotton, M.; Newbery, A.; Gowman, L.M. Global wildland fire season severity in the 21st century. For. Ecol. Manag. 2013, 294, 54–61. [Google Scholar] [CrossRef]
- Fox-Hughes, P.; Harris, R.; Lee, G.; Grose, M.; Bindoff, N. Future fire danger climatology for Tasmania, Australia, using a dynamically downscaled regional climate model. Int. J. Wildland Fire 2014, 23, 309–321. [Google Scholar] [CrossRef]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J. Resprouting as a key functional trait in woody plants—challenges to developing new organizing principles. New Phytol. 2010, 188, 651–654. [Google Scholar] [CrossRef]
- Noble, I.R.; Slatyer, R.O. The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 1980, 43, 5–21. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant functional traits in relation to fire in crown-fire ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef] [Green Version]
- Prior, L.D.; Bowman, D.M.J.S. Classification of post-fire responses of woody plants to include pyrophobic communities. Fire 2020, 3, 15. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Enright, N.J.; Fontaine, J.B.; Bowman, D.M.J.S.; Bradstock, R.A.; Williams, R.J. Interval squeeze: Altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 2015, 13, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Westerling, A.L.; Turner, M.G.; Smithwick, E.A.H.; Romme, W.H.; Ryan, M.G. Continued warming could transform Greater Yellowstone fire regimes by mid-21st century. Proc. Natl. Acad. Sci. USA 2011, 108, 13165–13170. [Google Scholar] [CrossRef] [Green Version]
- Bowman, D.M.J.S.; Bliss, A.; Bowman, C.J.W.; Prior, L.D. Fire caused demographic attrition of the Tasmanian palaeoendemic conifer Athrotaxis cupressoides. Austral Ecol. 2019, 44, 1322–1339. [Google Scholar] [CrossRef]
- Jackson, W.D. Vegetation of the Central Plateau. In The Lake Country; Banks, M.R., Ed.; Royal Society of Tasmania: Hobart, Australia, 1973; pp. 61–86. [Google Scholar]
- Johnson, K.; Marsden-Smedley, J.B. Fire history of the northern part of the Tasmanian Wilderness World Heritage Area and its associated regions. Pap. Proc. R. Soc. Tasman. 2002, 136, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, R.R. Land use on the Central Plateau with Special Reference to the Grazing Industry. In The Lake Country; Banks, M.R., Ed.; Royal Society of Tasmania: Hobart, Australia, 1973; pp. 161–176. [Google Scholar]
- Cullen, P. Land Degradation on the Central Plateau, Tasmania: The Legacy of 170 Years of Exploitation; Earth Science Section, Parks and Wildlife Service, Dept. of Environment and Land Management: Hobart, Australia, 1995. [Google Scholar]
- Misiak, W.M. Limits to Plant Regeneration in Alpine Vegetation on Tasmania’s Central Plateau. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2007. [Google Scholar]
- Costin, A.B.; Wimbush, D.J.; Kerr, D.; Gay, L.W. Studies in Catchment Hydrology in the Australian Alps. I. Trends in Soils and Vegetation; Technical Paper no.13; CSIRO: Canberra, Australia, 1959. [CrossRef]
- Williams, R.J.; Ashton, D.H. Effects of disturbance and grazing by cattle on the dynamics of heathland and grassland communities on the Bogong High Plains, Victoria. Aust. J. Bot. 1987, 35, 413–431. [Google Scholar] [CrossRef]
- Williamson, G.J.; Murphy, B.P.; Bowman, D.M.J.S. Cattle grazing does not reduce fire severity in eucalypt forests and woodlands of the Australian Alps. Austral Ecol. 2014, 39, 462–468. [Google Scholar] [CrossRef]
- AFAC. A Review of the Management of the Tasmanian Fires of December 2018 to March 2019; Technical Report; Australasian Fire and Emergency Service Authorities Council Limited: East Melbourne, VIC, Australia, 2019. [Google Scholar]
- Kirkpatrick, J.B.; Dickinson, K.J.M. The impact of fire on Tasmanian alpine vegetation and soils. Aust. J. Bot. 1984, 32, 613–629. [Google Scholar] [CrossRef]
- Balmer, J.; Whinam, J.; Kelman, J.; Kirkpatrick, J.B.; Lazarus, E. A Review of the Floristic Values of the Tasmanian Wilderness World Heritage Area. Nature Conservation Report 2004/3; Department of Primary Industries Water and Environment: Tasmania, Australia, 2004.
- Tasmanian Vegetation Mapping Program (TVMP). TASVEG Version 4.0, Tasmanian Vegetation Map 1:25,000; Department of Primary Industries, Water and Environment: Hobart, Australia, 2020.
- Banks, M.R. (Ed.) General Geology. In The Lake Country; Royal Society of Tasmania: Hobart, Australia, 1973; pp. 25–33. [Google Scholar]
- Gentilli, J. Australian Climate Patterns; Thomas Nelson (Australia) Limited: Melbourne, Australia, 1972. [Google Scholar]
- Kirkpatrick, J.B. Treeless plant communities of the Tasmanian high country. Proc. Ecol. Soc. Aust. 1983, 12, 61–77. [Google Scholar]
- Bureau of Meteorology. Climate Data Online. Climate Services. Bureau of Meteorology. Available online: http://www.bom.gov.au/ (accessed on 17 September 2020).
- Giglio, L.; Descloitres, J.; Justice, C.O.; Kaufman, Y.J. An enhanced contextual fire detection algorithm for MODIS. Remote Sens. Environ. 2003, 87, 273–282. [Google Scholar] [CrossRef]
- Williamson, G.J.; Price, O.F.; Henderson, S.B.; Bowman, D.M.J.S. Satellite-based comparison of fire intensity and smoke plumes from prescribed fires and wildfires in south-eastern Australia. Int. J. Wildland Fire 2013, 22, 121–129. [Google Scholar] [CrossRef]
- French, B.J.; Prior, L.D.; Williamson, G.J.; Bowman, D.M.J.S. Cause and effects of a megafire in sedge-heathland in the Tasmanian temperate wilderness. Aust. J. Bot. 2016, 64, 513–525. [Google Scholar] [CrossRef]
- Murphy, B.P.; Prior, L.D.; Cochrane, M.A.; Williamson, G.J.; Bowman, D.M.J.S. Biomass consumption by surface fires across Earth’s most fire prone continent. Glob. Chang. Biol. 2018, 25, 254–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, J.M.; Oechel, W.C. A simple method for estimating fire intensity after a burn in California chaparral. Acta Oecol./Oecol. Plant 1989, 10, 57–68. [Google Scholar]
- Whight, S.; Bradstock, R. Indices of fire characteristics in sandstone heath near Sydney, Australia. Int. J. Wildland Fire 1999, 9, 145–153. [Google Scholar] [CrossRef]
- Bliss, A.; Prior, L.D.; Bowman, D.M.J.S. Lack of reliable post-fire recovery mechanisms makes the iconic Tasmanian conifer Athrotaxis Cupressoides susceptible to population decline. Aust. J. Bot. 2021, 69, 162–173. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Ishiguro, M.; Kitagawa, G. Akaike Information Criterion Statistics; D. Reidel: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Vuong, Q.H. Likelihood ratio tests for model selection and non-nested hypotheses. Econom. J. Econom. Soc. 1989, 57, 307–333. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Nicholson, A.; Prior, L.D.; Perry, G.L.W.; Bowman, D.M.J.S. High post-fire mortality of resprouting woody plants in Tasmanian Mediterranean-type vegetation. Int. J. Wildland Fire 2017, 26, 532–537. [Google Scholar] [CrossRef]
- Walsh, N.G.; McDougall, K.L. Progress in the recovery of the flora of treeless subalpine vegetation in Kosciuszko National Park after the 2003 fires. Cunninghamia 2004, 8, 439–452. [Google Scholar]
- Clarke, P.J.; Knox, K.J.E.; Campbell, M.L.; Copeland, L.M. Post-fire recovery of woody plants in the New England Tableland Bioregion. Cunninghamia 2009, 11, 221–239. [Google Scholar]
- Bell, D.T. Ecological response syndromes in the flora of southwestern Western Australia: Fire resprouters versus reseeders. Bot. Rev. 2001, 67, 417–440. [Google Scholar] [CrossRef]
- Vesk, P.A.; Westoby, M. Sprouting ability across diverse disturbances and vegetation types worldwide. J. Ecol. 2004, 92, 310–320. [Google Scholar] [CrossRef]
- Vesk, P.A.; Warton, D.I.; Westoby, M. Sprouting by semi-arid plants: Testing a dichotomy and predictive traits. Oikos 2004, 107, 72–89. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Bridle, K.L.; Wild, A.S. Succession after fire in alpine vegetation on Mount Wellington, Tasmania. Aust. J. Bot. 2002, 50, 145–154. [Google Scholar] [CrossRef]
- Twidwell, D.; Rogers, W.E.; Wonkka, C.L.; Taylor, C.A., Jr.; Kreuter, U.P. Extreme prescribed fire during drought reduces survival and density of woody resprouters. J. Appl. Ecol. 2016, 53, 1585–1596. [Google Scholar] [CrossRef] [Green Version]
- Calder, J.A.; Kirkpatrick, J.B. Climate change and other factors influencing the decline of the Tasmanian cider gum (Eucalyptus gunnii). Aust. J. Bot. 2008, 56, 684–692. [Google Scholar] [CrossRef]
- Piper, F.I.; Paula, S. The role of nonstructural carbohydrates storage in forest resilience under climate change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bridle, K.L.; Kirkpatrick, J.B.; Cullen, P.; Shepherd, R.R. Recovery in alpine heath and grassland following burning and grazing, Eastern Central Plateau, Tasmania, Australia. Arctic Antarct. Alp. Res. 2001, 33, 348–356. [Google Scholar] [CrossRef]
- McPhail, D.C.; Kirkpatrick, J.B. Patch mosaic cyclic succession associated with the growth and senescence of an alpine shrub. Aust. J. Bot. 2016, 64, 111–119. [Google Scholar] [CrossRef]
- Williams, R.J. Gap dynamics in subalpine heathland and grassland vegetation in south-eastern Australia. J. Ecol. 1992, 80, 343–352. [Google Scholar] [CrossRef]
- Bridle, K.L.; Kirkpatrick, J.B. Comparative effects of stock and wild vertebrate herbivore grazing on treeless subalpine vegetation, Eastern Central Plateau, Tasmania. Aust. J. Bot. 1999, 47, 817–834. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Bridle, K.L. Grazing and the absence of fire promote the dominance of an unpalatable shrub in a patch mosaic cyclic successional system. Aust. J. Bot. 2016, 64, 45–50. [Google Scholar] [CrossRef]
- Kirkpatrick, J.B.; Marsden-Smedley, J.B.; Folco, M.B.D.; Leonard, S.W.J. Influence of grazing and vegetation type on post-fire floristic and lifeform composition in Tasmania, Australia. Plant Ecol. 2016, 217, 57–69. [Google Scholar] [CrossRef]
- Styger, J.K.; Kirkpatrick, J.B.; Marsden-Smedley, J.O.N.; Leonard, S.W.J. Fire incidence, but not fire size, affects macropod densities. Austral Ecol. 2011, 36, 679–686. [Google Scholar] [CrossRef]
- Locke, S. The Distribution and Abundance of Fallow Deer in the Central Plateau Conservation Area and Adjacent Areas in Tasmania: A Baseline Monitoring Program; Nature Conservation Report 07/02; Department of Primary Industries and Water: Hobart, Australia, 2007.
- Kirkpatrick, J.B.; Bridle, K.L.; Dickinson, K.J.M. Decades-scale vegetation change in burned and unburned alpine coniferous heath. Aust. J. Bot. 2010, 58, 453–462. [Google Scholar] [CrossRef]
- Williams, R.J.; Wahren, C.H.; Shannon, J.M.; Papst, W.A.; Heinze, D.A.; Camac, J.S. Fire regimes and biodiversity in Victoria’s alpine ecosystems. Proc. R. Soc. Vic. 2012, 124, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Fairman, T.A.; Nitschke, C.R.; Bennett, L.T. Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. Int. J. Wildland Fire 2016, 25, 831–848. [Google Scholar] [CrossRef]
- Batllori, E.; Ackerly, D.D.; Moritz, M.A. A minimal model of fire-vegetation feedbacks and disturbance stochasticity generates alternative stable states in grassland–shrubland–woodland systems. Environ. Res. Lett. 2015, 10, 034018. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Briggs, J.M.; Blair, J.M. Fire dynamics distinguish grasslands, shrublands and woodlands as alternative attractors in the Central Great Plains of North America. J. Ecol. 2014, 102, 1374–1385. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, Z.; Nippert, J.B.; Ocheltree, T.W. Abrupt transition of mesic grassland to shrubland: Evidence for thresholds, alternative attractors, and regime shifts. Ecology 2014, 95, 2633–2645. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, A.T. Grazing systems, ecosystem responses, and global change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Noble, J.C.; Hik, D.S.; Sinclair, A.R.E. Landscape ecology of the burrowing bettong: Fire and marsupial biocontrol of shrubs in semi-arid Australia. Rangel. J. 2007, 29, 107–119. [Google Scholar] [CrossRef]
- Van Auken, O.W. Shrub invasions of North American semiarid grasslands. Annu. Rev. Ecol. Syst. 2000, 31, 197–215. [Google Scholar] [CrossRef] [Green Version]
- Bowman, D.M.J.S.; Wood, S.W.; Neyland, D.; Sanders, G.J.; Prior, L.D. Contracting Tasmanian montane grasslands within a forest matrix is consistent with cessation of Aboriginal fire management. Austral Ecol. 2013, 38, 627–638. [Google Scholar] [CrossRef]
- Romanin, L.M.; Hopf, F.; Haberle, S.G.; Bowman, D.M.J.S. Fire regime and vegetation change in the transition from Aboriginal to European land management in a Tasmanian eucalypt savanna. Aust. J. Bot. 2016, 64, 427–440. [Google Scholar] [CrossRef] [Green Version]
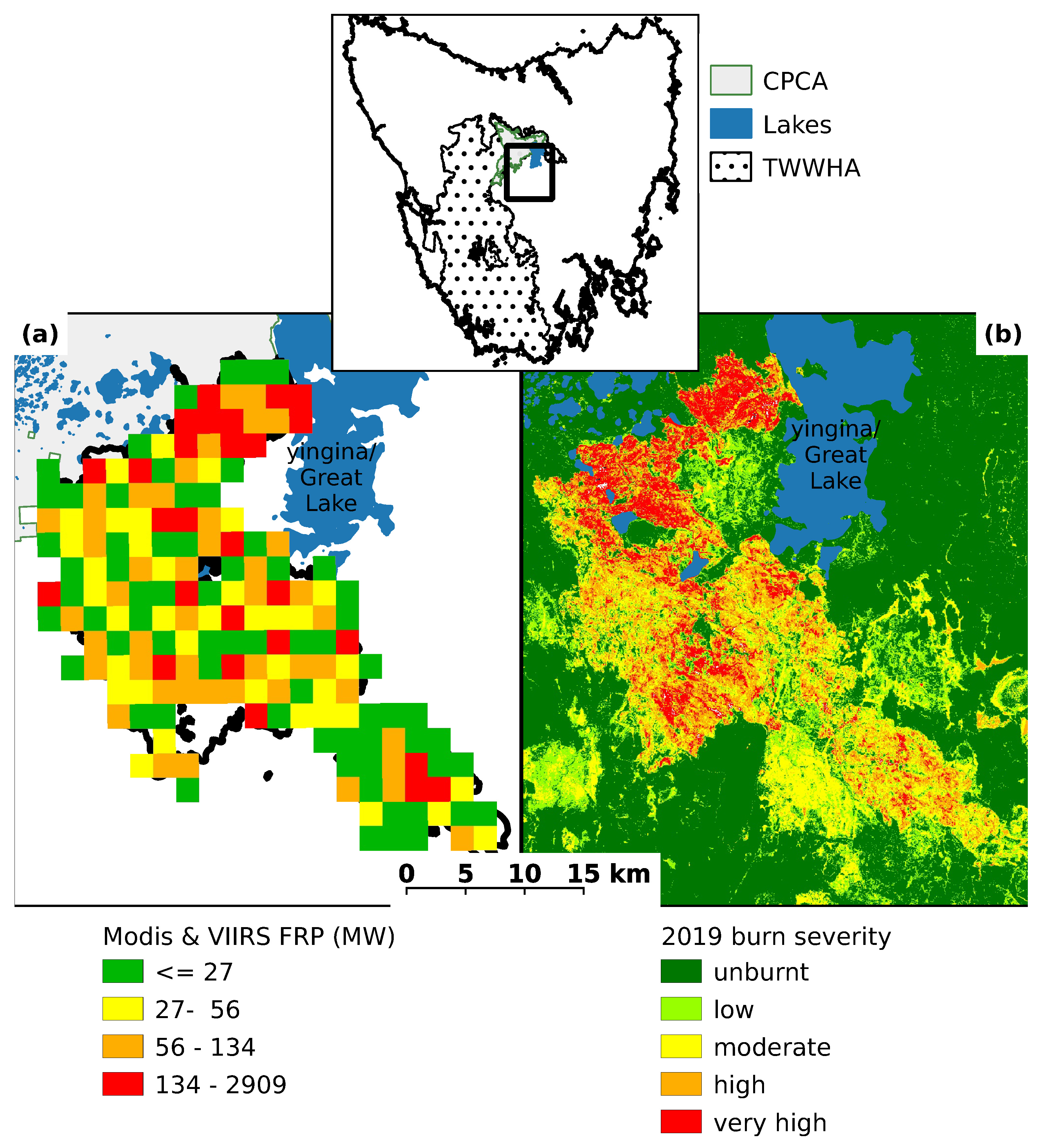



| Fire Severity | CPCA Fire Area | Survey Plots | ||
|---|---|---|---|---|
| % Total Area | % Shrubland | % Total Area | % Shrubland | |
| Unburnt | 5.2 | 4.4 | 5 | 8.3 |
| Low | 4.7 | 3.2 | 1 | 1.7 |
| Medium | 14.3 | 8.2 | 2 | 3.3 |
| High | 29.2 | 26.1 | 8 | 13.3 |
| Very high | 46.6 | 58.0 | 44 | 73.3 |
| Species | Unburnt | Burnt | Condition of Shrubs in the Burnt Plots | Shrub Basal Area (cm) ± s.d. | |||
|---|---|---|---|---|---|---|---|
| Alive | Dead | Resp. | Unburnt | Burnt | |||
| Acrothamnus montanus (R.Br.) Quinn | 0 | 1 | 1 | 0 | 0 | n.a. | 250 |
| Bauera rubioides Andrews | 0 | 1 | 0 | 0 | 1 | n.a | 135 |
| Leptospermum rupestre Hook.f. | 0 | 15 | 1 | 12 | 2 | n.a | 169 ± 256 |
| Orites acicularis (R.Br.) Roem. & Schult. | 14 | 40 | 0 | 40 | 0 | 303 ± 394 | 225 ± 216 |
| Orites revoluta R.Br. | 9 | 77 | 2 | 35 | 40 | 223 ± 294 | 528 ± 1130 |
| Ozothamnus hookeri Sond. | 30 | 13 | 0 | 13 | 0 | 141 ± 223 | 108 ± 101 |
| Richea acerosa (Lindl.) F.Muell. | 24 | 54 | 9 | 45 | 0 | 224 ± 306 | 178 ± 228 |
| Richea sprengeloides (R.Br.) F.Muell. | 0 | 43 | 0 | 43 | 0 | n.a | 65 ± 72 |
| Unknown | 0 | 715 | 0 | 715 | 0 | n.a | 72 ± 97 |
| Total | 77 | 959 | 8 | 901 | 43 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foulkes, J.A.; Prior, L.D.; Leonard, S.W.J.; Bowman, D.M.J.S. Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau. Fire 2021, 4, 32. https://doi.org/10.3390/fire4030032
Foulkes JA, Prior LD, Leonard SWJ, Bowman DMJS. Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau. Fire. 2021; 4(3):32. https://doi.org/10.3390/fire4030032
Chicago/Turabian StyleFoulkes, Judy A., Lynda D. Prior, Steven W. J. Leonard, and David M. J. S. Bowman. 2021. "Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau" Fire 4, no. 3: 32. https://doi.org/10.3390/fire4030032
APA StyleFoulkes, J. A., Prior, L. D., Leonard, S. W. J., & Bowman, D. M. J. S. (2021). Demographic Effects of Severe Fire in Montane Shrubland on Tasmania’s Central Plateau. Fire, 4(3), 32. https://doi.org/10.3390/fire4030032