
Changes in Soil Water Retention and Micromorphological Properties Induced by Wetting and Drying Cycles


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Wetting and Drying (W-D) Cycles
2.3. Water Retention Measurement
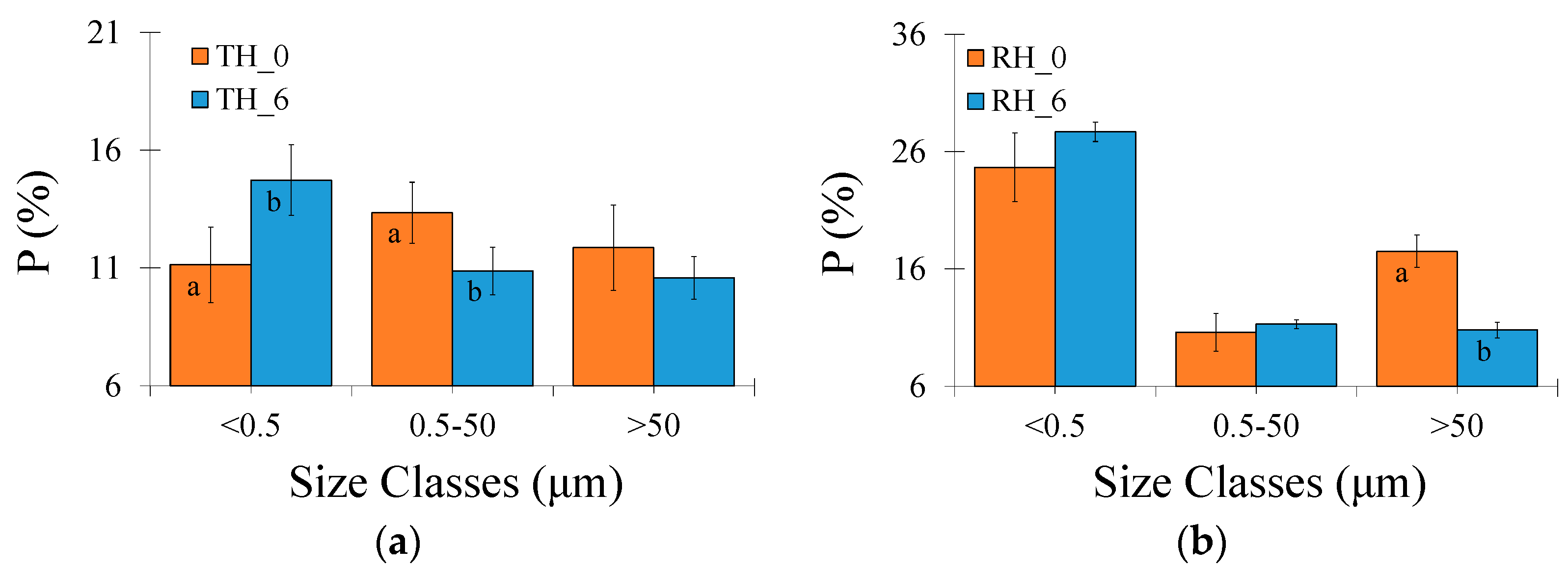
2.4. Micromorphological Analysis
2.5. Statistical Analysis
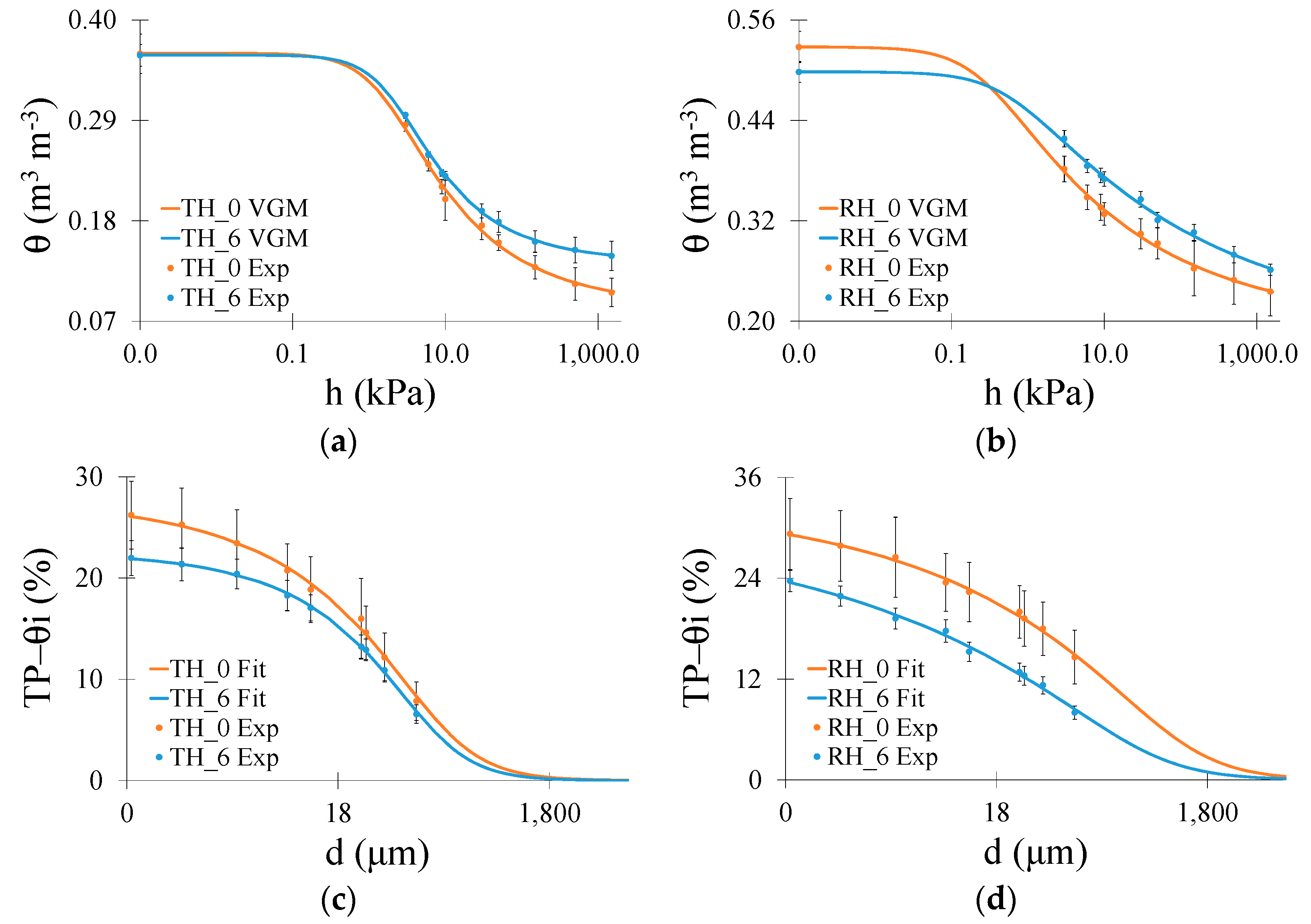
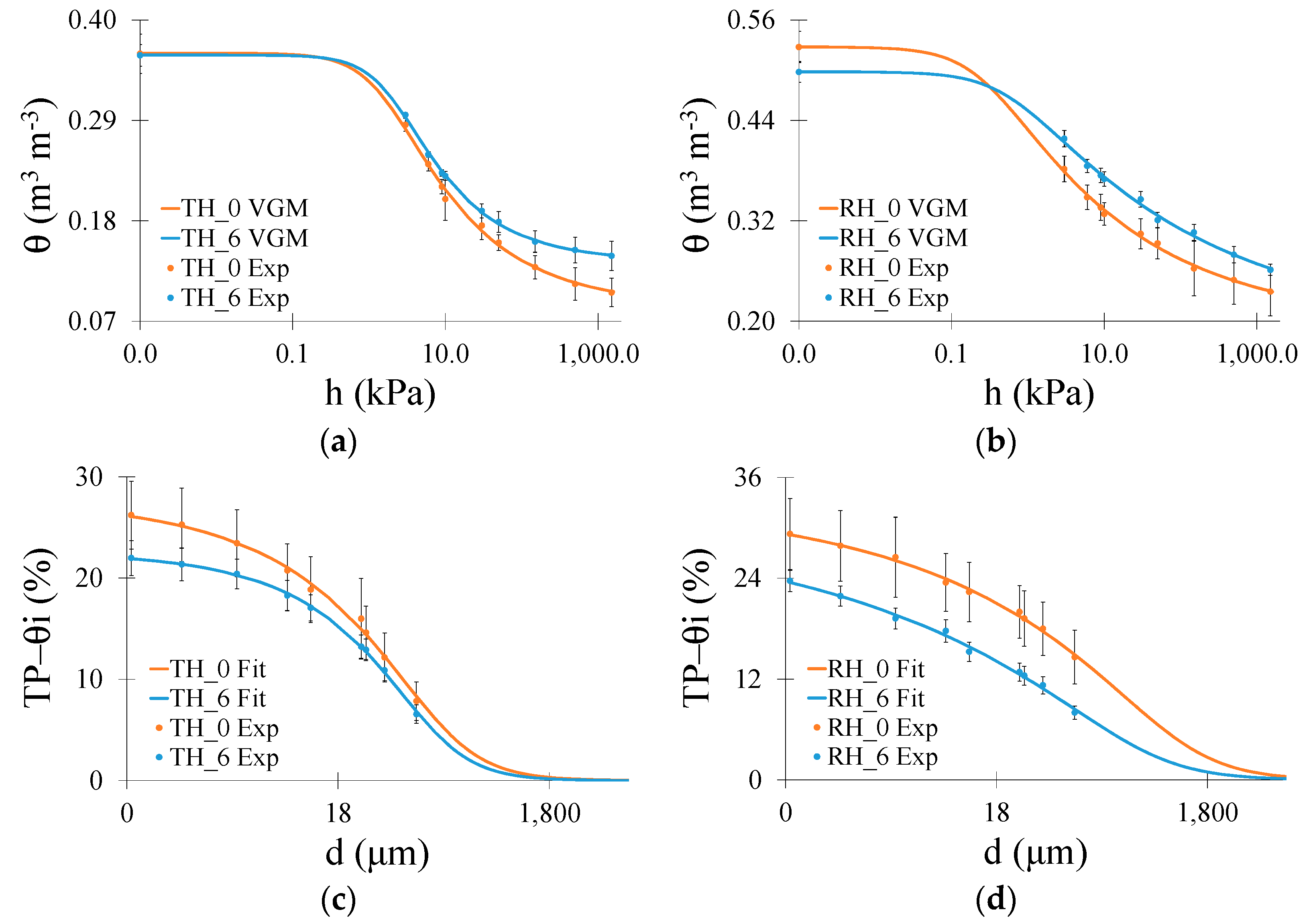
3. Results
3.1. Water Retention Measurements
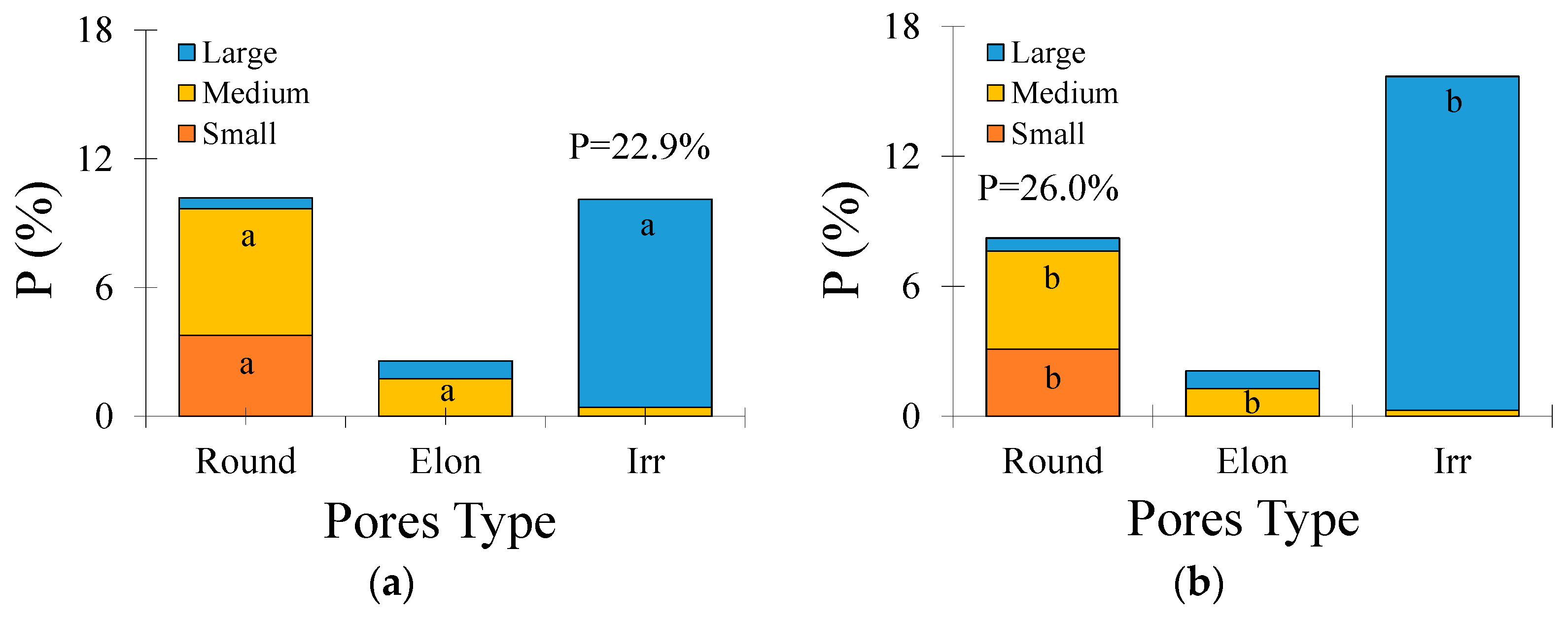
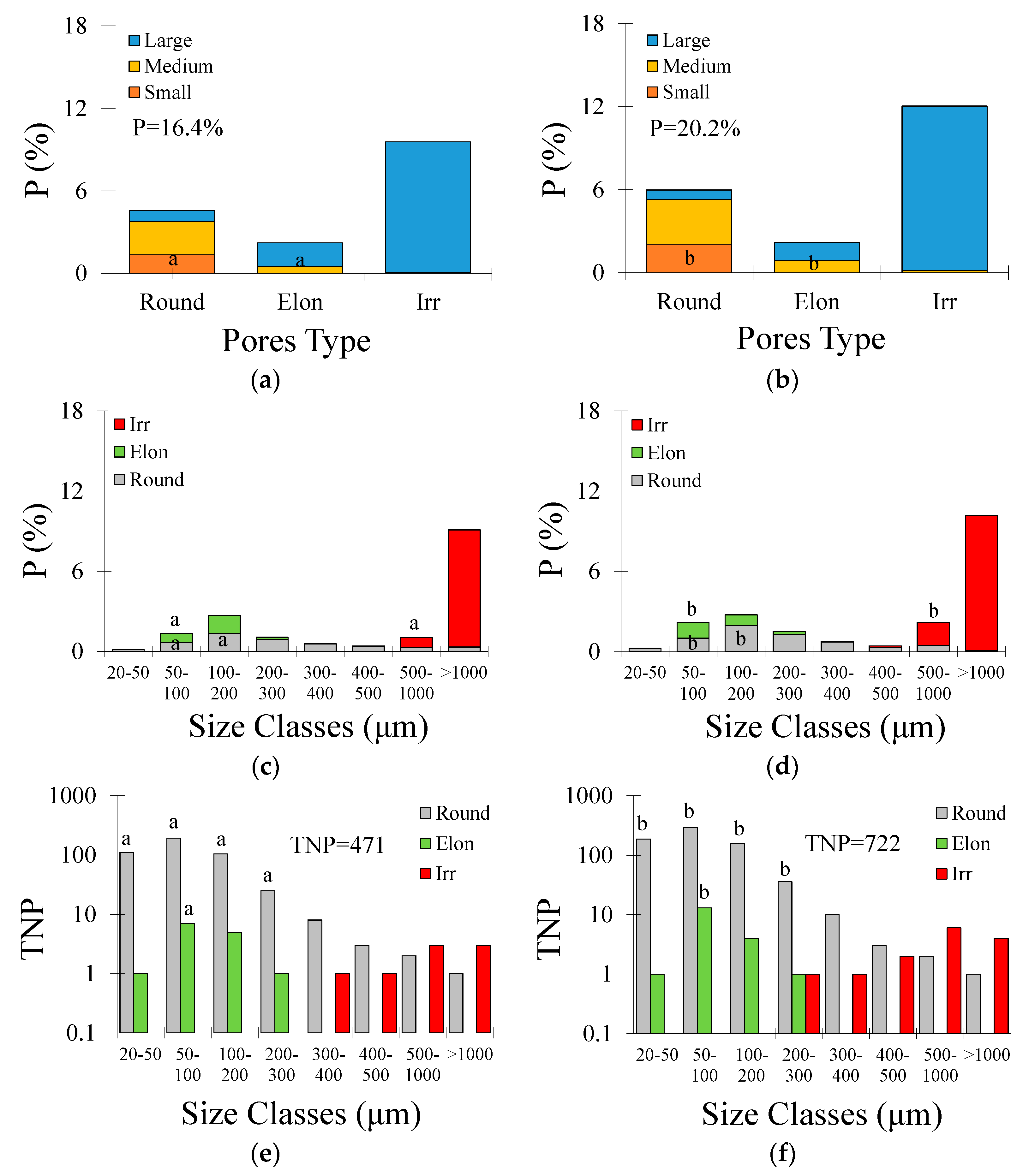
3.2. Micromorphological Soil Properties
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R.; Shukla, M.K. Principles of Soil Physics; Marcel Dekker, Inc.: New York, NY, USA, 2004. [Google Scholar]
- Reynolds, W.D.; Bowman, B.T.; Drury, C.F.; Tan, C.S.; Lu, X. Indicators of Good Soil Physical Quality: Density and Storage Parameters. Geoderma 2002, 110, 131–146. [Google Scholar] [CrossRef]
- Correchel, V.; Bacchi, O.O.S.; De Maria, I.C.; Dechen, S.C.F.; Reichardt, K. Erosion Rates Evaluated by the 137Cs Technique and Direct Measurements on Long-Term Runoff Plots under Tropical Conditions. Soil Tillage Res. 2006, 86, 199–208. [Google Scholar] [CrossRef]
- Tracy, S.R.; Black, C.R.; Roberts, J.A.; Sturrock, C.; Mairhofer, S.; Craigon, J.; Mooney, S.J. Quantifying the Impact of Soil Compaction on Root System Architecture in Tomato (Solanum lycopersicum) by X-Ray Micro-Computed Tomography. Ann. Bot. 2012, 110, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Hillel, D. Environmental Soil Physics; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Reichardt, K.; Timm, L.C. Soil, Plant and Atmosphere: Concepts, Processes and Applications; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Nimmo, J.R. Porosity and Pore Size Distribution; Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Juhász, C.E.P.; Cooper, M.; Cursi, P.R.; Ketzer, A.O.; Toma, R.S. Savanna Woodland and Soil Micromorphology Related to Water Retention. Sci. Agric. 2007, 64, 344–354. [Google Scholar] [CrossRef]
- Bouma, J.; Jongerius, A.; Boersma, O.; Jager, A.; Schoonderbeek, D. The Function of Different Types of Macropores During Saturated Flow through Four Swelling Soil Horizons. Soil Sci. Soc. Am. J. 1977, 41, 945–950. [Google Scholar] [CrossRef]
- Fox, D.M.; Bryan, R.B.; Fox, C.A. Changes in Pore Characteristics with Depth for Structural Crusts. Geoderma 2004, 120, 109–120. [Google Scholar] [CrossRef]
- De Pierri Castilho, S.C.; Cooper, M.; Simões da Silva, L.F. Micromorphometric Analysis of Porosity Changes in the Surface Crusts of Three Soil in the Piracicaba Region, São Paulo State, Brazil. Acta Sci. Agron. 2015, 37, 385–395. [Google Scholar] [CrossRef]
- Cooper, M.; Vidal-Torrado, P. Caracterização Morfológica, Micromorfológica e Físico-Hídrica de Solos com Horizonte B Nítico. Rev. Bras. Ciênc. Solo 2005, 29, 581–595. [Google Scholar] [CrossRef]
- Pagliai, M.; Marsili, A.; Servadio, P.; Vignozzi, N.; Pellegrini, S. Changes in Some Physical Properties of a Clay Soil in Central Italy Following the Passage of Rubber Tracked and Wheeled Tractors of Medium Power. Soil Tillage Res. 2003, 73, 119–129. [Google Scholar] [CrossRef]
- Hussein, J.; Adey, M.A. Changes in Microstructure, Voids and b-Fabric of Surface Samples of a Vertisol Caused by Wet/Dry Cycles. Geoderma 1998, 85, 63–82. [Google Scholar] [CrossRef]
- Tang, C.S.; Cui, Y.J.; Shi, B.; Tang, A.M.; Liu, C. Desiccation and Cracking Behaviour of Clay Layer from Slurry State under Wetting-Drying Cycles. Geoderma 2011, 166, 111–118. [Google Scholar] [CrossRef]
- An, R.; Zhang, X.; Kong, L.; Liu, X.; Chen, C. Drying-Wetting Impacts on Granite Residual Soil: A Multi-Scale Study from Macroscopic to Microscopy Investigations. Bull. Eng. Geol. 2022, 81, 447. [Google Scholar] [CrossRef]
- Pardini, G.; Vigna Guidi, G.; Pini, R.; Regüés, D.; Gallart, F. Structure and Porosity of Smectitic Mudrocks as Affected by Experimental Wetting—Drying Cycles and Freezing—Thawing Cycles. Catena 1996, 27, 149–165. [Google Scholar] [CrossRef]
- Xia, J.; Zhang, L.; Ge, P.; Lu, X.; Wei, Y.; Cai, C.; Wang, J. Structure Degradation Induced by Wetting and Drying Cycles for the Hilly Granitic Soils in Collapsing Gully Erosion Areas. Forests 2022, 13, 1426. [Google Scholar] [CrossRef]
- Louati, F.; Trabelsi, H.; Jamei, M.; Taibi, S. Impact of Wetting-Drying Cycles and Cracks on the Permeability of Compacted Clayey Soil. Eur. J. Environ. Civ. 2018, 25, 696–721. [Google Scholar] [CrossRef]
- Ng, C.W.W.; Peprah-Manu, D. Pore Structure Effects on the Water Retention Behaviour of a Compacted Silty Sand Soil Subjected to Drying-Wetting Cycles. Eng. Geol. 2023, 313, 106963. [Google Scholar] [CrossRef]
- Momoli, R.S.; Cooper, M.; de Pierri Castilho, S.C. Sediment Morphology and Distribution in a Restored Riparian Forest. Sci. Agric. 2007, 64, 486–494. [Google Scholar] [CrossRef]
- Pires, L.F.; Cooper, M.; Cássaro, F.A.M.; Reichardt, K.; Bacchi, O.O.S.; Dias, N.M.P. Micromorphological Analysis to Characterize Structure Modifications of Soil Samples Submitted to Wetting and Drying Cycles. Catena 2008, 72, 297–304. [Google Scholar] [CrossRef]
- Lima, H.V.; Silva, A.P.; Santos, M.C.; Cooper, M.; Romero, R.E. Micromorphology and Image Analysis of a Hardsetting Ultisol (Argissolo) in the State of Ceará (Brazil). Geoderma 2006, 132, 416–426. [Google Scholar] [CrossRef]
- Lipiec, J.; Walczak, R.; Witkowska-Walczak, B.; Nosalewicz, A.; Słowińska-Jurkiewicz, A.; Sławiński, C. The Effect of Aggregate Size on Water Retention and Pore Structure of Two Silt Loam Soils of Different Genesis. Soil Tillage Res. 2007, 97, 239–246. [Google Scholar] [CrossRef]
- Cooper, M.; Vidal-Torrado, P.; Chaplot, V. Origin of Microaggregates in Soils with Ferralic Horizons. Sci. Agric. 2005, 62, 256–263. [Google Scholar] [CrossRef]
- Hobson, D.; Harty, M.; Tracy, S.R.; McDonnell, K. The Effect of Tillage Depth and Traffic Management on Soil Properties and Root Development during Two Growth Stages of Winter Wheat (Triticum aestivum L.). Soil 2022, 8, 391–408. [Google Scholar] [CrossRef]
- Dal Ferro, N.; Sartori, L.; Simonetti, G.; Berti, A.; Morari, F. Soil Macro- and Microstructure as Affected by Different Tillage Systems and their Effects on Maize Root Growth. Soil Tillage Res. 2014, 140, 55–65. [Google Scholar] [CrossRef]
- Budhathoki, S.; Lamba, J.; Srivastava, P.; Williams, C.; Arriaga, F.; Karthikeyan, K.G. Impact of Land Use and Tillage Practice on Soil Macropore Characteristics Inferred from X-ray Computed Tomography. Catena 2022, 210, 105886. [Google Scholar] [CrossRef]
- Helliwell, J.R.; Sturrock, C.J.; Grayling, K.M.; Tracy, S.R.; Flavel, R.J.; Young, I.M.; Whalley, W.R.; Mooney, S.J. Applications of X-ray Computed Tomography for Examining Biophysical Interactions and Structural Development in Soil Systems: A Review. Eur. J. Soil Sci. 2013, 64, 279–297. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Negassa, W.C.; Guber, A.K.; Rivers, M.L. Protection of Soil Carbon within Macro-Aggregates depends on Intra-Aggregate Pore Characteristics. Sci. Rep. 2015, 5, 16261. [Google Scholar] [CrossRef]
- Koestel, J.; Schlüter, S. Quantification of the Structure Evolution in a Garden Soil over the Course of Two Years. Geoderma 2019, 338, 597–609. [Google Scholar] [CrossRef]
- Soil Survey Staff. Simplified Guide to Soil Taxonomy; USDA Natural Resources Conservation Service, National Soil Survey Center: Lincoln, NE, USA, 2013. [Google Scholar]
- Booman, G.; Leiker, S. Soil Sampling Guide; Document ID: RND_SSG_001; Regen Network Development, Inc.: Northfield, MA, USA, 2021. [Google Scholar]
- Klute, A. Water Retention: Laboratory Methods. In Methods of Soil Analysis. Part 1: Physical and Mineralogical Methods; Black, C.A., Ed.; Soil Science Society of America: Madison, WA, USA, 1986; pp. 635–662. [Google Scholar]
- Dane, J.H.; Hopmans, J.W. Pressure Plate Extractor. In Methods of Soil Analysis. Part 4: Physical Methods; Dane, J.H., Topp, G.C., Eds.; Soil Science Society of America: Madison, WA, USA, 2002; pp. 688–690. [Google Scholar]
- Reynolds, W.D.; Drury, C.F.; Yang, X.M.; Tan, C.S. Optimal Soil Physical Quality Inferred Through Structural Regression and Parameter Interactions. Geoderma 2008, 146, 466–474. [Google Scholar] [CrossRef]
- Van Genuchten, M.T. A Closed-Form Equation for Predicting the Hydraulic Conductivity of Unsaturated Soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- Seki, K. SWRC Fit—A Nonlinear Fitting Program with a Water Retention Curve for Soils Having Unimodal and Bimodal Pore Structure. Hydrol. Earth Syst. Sci. Discuss. 2007, 4, 407–437. [Google Scholar]
- Jury, W.A.; Horton, R. Soil Physics; Willey: New Jersey, NJ, USA, 2004. [Google Scholar]
- Reynolds, W.D.; Drury, C.F.; Tan, C.S.; Fox, C.A.; Yang, X.M. Use of Indicators and Pore Volume-Function Characteristics to Quantify Soil Physical Quality. Geoderma 2009, 152, 252–263. [Google Scholar] [CrossRef]
- Tarawally, M.A.; Medina, H.; Frómeta, M.E.; Alberto Itza, C. Field Compaction at Different Soil-Water Status: Effects on Pore Size Distribution and Soil Water Characteristics of a Rhodic Ferralsol in Western Cuba. Soil Tillage Res. 2004, 76, 95–103. [Google Scholar] [CrossRef]
- Ringrose-Voase, A.J.; Bullock, P. The Automatic Recognition and Measurement of Soil Pore Types by Image Analysis and Computer Programs. J. Soil Sci. 1984, 35, 673–684. [Google Scholar] [CrossRef]
- Huf dos Reis, A.M.; Armindo, R.A.; Pires, L.F. Physical Assessment of a Haplohumox Soil Under Integrated Crop-Livestock System. Soil Tillage Res. 2019, 194, 104294. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeont. Elect. 2001, 4, 1–9. [Google Scholar]
- Greenland, D.J. Soil Damage by Intensive Arable Cultivation: Temporary or Permanent? Philos. Trans. R. Soc. B 1977, 281, 193–208. [Google Scholar]
- Cássaro, F.A.M.; Pires, L.F.; dos Santos, R.A.; Gimenez, D.; Reichardt, K. Funil de Haines Modificado: Curvas de Retenção de Solos Próximos à Saturação. Rev. Bras. Ciênc. Solo 2008, 32, 2555–2562. [Google Scholar] [CrossRef]
- Pires, L.F.; Villanueva, F.C.A.; Dias, N.M.P.; Bacchi, O.O.S.; Reichardt, K. Chemical Migration During Soil Water Retention Curve Evaluation. Annu. Acad. Bras. Cienc. 2011, 83, 1097–1107. [Google Scholar] [CrossRef]
- Dapla, P.; Hriník, D.; Hrabovský, A.; Simkovic, I.; Zarnovican, H.; Sekucia, F.; Kollár, J. The Impact of Land-Use on the Hierarchical Pore Size Distribution and Water Retention Properties in Loamy Soils. Water 2020, 12, 339. [Google Scholar]
- Kutílek, M.; Jendele, L.; Panayiotopoulos, K.P. The Influence of Uniaxial Compression upon Pore Size Distribution in Bi-Modal Soils. Soil Tillage Res. 2006, 86, 27–37. [Google Scholar] [CrossRef]
- Bodner, G.; Scholl, P.; Kaul, H.-P. Field Quantification of Wetting–Drying Cycles to Predict Temporal Changes of Soil Pore Size Distribution. Soil Tillage Res. 2013, 133, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pires, L.F.; Bacchi, O.O.S.; Reichardt, K. Assessment of Soil Structure Repair due to Wetting and Drying Cycles through 2D Tomographic Image Analysis. Soil Tillage Res. 2007, 94, 537–545. [Google Scholar] [CrossRef]
- Testoni, S.A.; de Almeida, J.A.; da Silva, L.; Pugliese Andrade, G.R. Clay Mineralogy of Brazilian Oxisols with Shrinkage Properties. Rev. Bras. Ciênc. Solo 2017, 41, e0160487. [Google Scholar] [CrossRef]
- Lu, S.-G.; Malik, Z.; Chen, D.-P.; Wu, C.-F. Porosity and Pore Size Distribution of Ultisols and Correlations to Soil Iron Oxides. Catena 2014, 123, 79–87. [Google Scholar] [CrossRef]
- Kodešová, R.; Pavlů, L.; Kodeš, V.; Žigová, A.; Nikodem, A. Impact of Spruce Forest and Grass Vegetation Cover on Soil Micromorphology and Hydraulic Properties of Organic Matter Horizon. Biologia 2007, 62, 565–568. [Google Scholar] [CrossRef]
- Cooper, M.; Dalla Rosa, J.; Medeiros, J.C.; de Oliveira, T.C.; Toma, R.S.; Juhász, C.E.P. Hydro-Physical Characterization of Soils under Tropical Semi-Deciduous Forest. Sci. Agric. 2012, 69, 152–159. [Google Scholar] [CrossRef]
- Tang, C.-S.; Cheng, Q.; Gong, X.; Shi, B.; Inyang, H.I. Investigation on Microstructure Evolution of Clayey Soils: A Review Focusing on Wetting/Drying Process. J. Rock Mech. Geotech. Eng. 2023, 15, 269–284. [Google Scholar] [CrossRef]
- Peng, X.; Horn, R.; Smucker, A. Pore Shrinkage Dependency of Inorganic and Organic Soils on Wetting and Drying Cycles. Soil Sci. Soc. Am. J. 2007, 71, 1095–1104. [Google Scholar] [CrossRef]
- Leij, F.J.; Ghezzehei, T.A.; Or, D. Modeling the Dynamics of the Soil Pore-Size Distribution. Soil Tillage Res. 2002, 64, 61–78. [Google Scholar] [CrossRef]
- Hall, D.G.M.; Reeve, M.J.; Thomasson, A.J.; Wright, V.F. Water Retention, Porosity and Density of Field Soils; Soil Survey Technical Monograph; Rothamsted: Harpenden, UK, 1977; Volume 9. [Google Scholar]
- Pagliai, M.; La Marca, M.; Lucamante, G. Micromorphometric and Micromorphological Investigations of a Clay Loam Soil in Viticulture under Zero and Conventional Tillage. J. Soil Sci. 1983, 34, 391–403. [Google Scholar] [CrossRef]
- Ringrose-Voase, A.J. Measurements of Soil Macropore Geometry by Image Analysis of Sections Through Impregnated Soil. Plant Soil 1996, 183, 27–47. [Google Scholar] [CrossRef]
- Cooper, M.; Medeiros, J.C.; Dalla Rosa, J.; Soria, J.E.; Toma, R.S. Soil Functioning in a Toposequence under Rainforest in São Paulo, Brazil. Rev. Bras. Ciênc. Solo 2013, 37, 392–399. [Google Scholar] [CrossRef]
- Huf dos Reis, A.; Auler, A.C.; Armindo, R.A.; Cooper, M.; Pires, L.F. Micromorphological Analysis of Soil Porosity under Integrated Crop-Livestock Management Systems. Soil Tillage Res. 2021, 205, 104783. [Google Scholar] [CrossRef]
- Wen, T.; Chen, X.; Shao, L. Effect of Multiple Wetting and Drying Cycles on the Macropore Structure of Granite Residual Soil. J. Hydrol. 2022, 614, 128583. [Google Scholar] [CrossRef]
- Diel, J.; Vogel, H.J.; Schlüter, S. Impact of Wetting and Drying Cycles on Soil Structure Dynamics. Geoderma 2019, 345, 63–71. [Google Scholar] [CrossRef]
- Pagliai, M. Soil porosity aspects. Int. Agrophys. 1988, 4, 215–232. [Google Scholar]
- Sartori, G.; Ferrari, G.A.; Pagliai, M. Changes in Soil Porosity and Surface Shrinkage in a Remolded, Saline Clay Soil Treated with Compost. Soil Sci. 1985, 139, 523–530. [Google Scholar] [CrossRef]
- Pagliai, M.; La Marca, M.; Lucamante, G. Changes in Soil Porosity in Remoulded Soils Treated with Poultry Manure. Soil Sci. 1987, 144, 128–140. [Google Scholar] [CrossRef]
- Pagliai, M.; Guidi, G.; La Marca, M.; Giachetti, M.; Lucamante, G. Effect of sewage sludges and composts on soil porosity and aggregation. J. Environ. Qual. 1981, 10, 556–561. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pore Shapes | Γ1 | Γ2 |
|---|---|---|
| Rounded (Round) | ≤5 | - |
| Elongated (Elon) | 5 < Γ1 ≤ 25 | ≤2.2 |
| Irregular (Irr) | 5 < Γ1 ≤ 25 or >25 | >2.2 |
| VGM Parameters | Typic Hapludox | Rhodic Hapludox | ||
|---|---|---|---|---|
| 0 W-D | 6 W-D | 0 W-D | 6 W-D | |
| θs (m3 m−3) | 0.364 | 0.362 | 0.528 a | 0.498 b |
| θr (m3 m−3) | 0.087 a | 0.136 b | 0.195 | 0.190 |
| α (kPa−1) | 0.613 | 0.493 | 3.677 a | 1.695 b |
| n | 1.416 | 1.533 | 1.244 | 1.183 |
| m 1 | 0.294 | 0.348 | 0.196 | 0.155 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pires, L.F. Changes in Soil Water Retention and Micromorphological Properties Induced by Wetting and Drying Cycles. Soil Syst. 2023, 7, 51. https://doi.org/10.3390/soilsystems7020051
Pires LF. Changes in Soil Water Retention and Micromorphological Properties Induced by Wetting and Drying Cycles. Soil Systems. 2023; 7(2):51. https://doi.org/10.3390/soilsystems7020051
Chicago/Turabian StylePires, Luiz F. 2023. "Changes in Soil Water Retention and Micromorphological Properties Induced by Wetting and Drying Cycles" Soil Systems 7, no. 2: 51. https://doi.org/10.3390/soilsystems7020051
APA StylePires, L. F. (2023). Changes in Soil Water Retention and Micromorphological Properties Induced by Wetting and Drying Cycles. Soil Systems, 7(2), 51. https://doi.org/10.3390/soilsystems7020051