Abstract
This study assesses the impact of defoliation applied to three developmental stages across three cropping seasons from 2021 to 2023 on growth, yield and forage quality in maize. The experimental design included three treatments: defoliation of three expanded leaves at the 3rd–4th leaf stage (DF1), the 5th–6th expanded leaves by leaf punch (DF2) and expanding leaves with the DF2 treatment (DF3) at the 6th–7th leaf stages, compared with no defoliation (control). Over three years, the most significant decrease in dry matter (DM) yield occurred in DF1 during spring sowing, while in summer sowing, the largest reduction was in DF3, both of which were correlated with changes in the number of grains per ear. The DM yields at harvest were positively correlated with plant leaf areas at the silking stage. The digestibility of forage in in vitro DM decreased concomitantly with an increase in acid detergent fiber content, indicating a decrease in forage quality. Given the frequent severe damage observed in summer sown maize and the detrimental effects of early growth stage leaf feeding on quality and quantity of spring sown maize, the application of registered insecticides is advised to reduce pest damage to maize crops.
Keywords:
clipping; cropping season; quantity and quality; pest damage; yield component; Zea mays L. 1. Introduction
The fall armyworm (FAW, Spodoptera frugiperda), which is native to the subtropical and tropical regions of the Americas, has undergone a rapid ‘west-to-east’ range expansion, spreading from the west African countries of Nigeria and Ghana where it was first detected in 2016 [1,2], through to Kenya in East Africa [3] and onward to Indonesia [4] and the Philippines [5] in Southeast Asia. This highly polyphagous and invasive pest persists through winter in regions where temperatures do not fall below 10 °C, reflecting the larval lowest threshold temperature of 10.39 °C [6]. In North America, adult FAW migrate northwards from the winter breeding regions in southern Texas and southern Florida, transitioning through several generations before reaching regions as far north as Québec and Ontario in Canada, where it damages maize crops from spring to autumn [7]. In Japan, the FAW was first recorded in Kagoshima Prefecture in early July 2019, and the pest had invaded 21 of the 47 prefectures in Japan (i.e., from Kyushu-Okinawa to the Tohoku region) by December 2019 [8]. In the first winter of 2019–2020, the FAW was observed year-round in tropical and subtropical regions of southern China [9]. Significant migrations to the Japanese mainland began in May 2020 [8], and by November 2020, the FAW had spread to 42 Prefectures, including Hokkaido, causing extensive damage to 12 agricultural crops [10].
Kyushu employs several cropping systems for maize because of its warm climate, allowing planting from late March to early August [11]. The three main systems include spring sowing from March to April with harvesting in late July to August, late sowing from May to June with harvesting from August to October and summer sowing from July to August with harvesting in November, complemented by a double cropping systems that combines spring and summer sowing crops [12].
The damage inflicted by the FAW exhibits distinct characteristics at different growth stages. Initially, young larvae (first to second instar) scrape at the 3rd to 4th maize leaf stages, subsequently migrating to feed within the whorls on the expanding leaves and stalks as third to fourth instars, or older when the maize plants are at the early 5th to 6th leaf stages, prior to stem elongation [13]. This phase typically leaves insect bite marks along the midrib of expanded leaves [14]. High larval densities can lead to severe damage, consuming almost all expanding leaves and potentially killing the maize. Several phenotyping techniques have been developed and validated for assessing yield loss due to the FAW [15,16]. In India, across the several artificial larval load of FAW to maize genotypes in net house, it tended to increase maize leaf damage rating score with the increase in the larval load at 14 days after infection and the authors discovered the resistant maize germplasm to the larval load [15]. In Indonesia, the damages of FAW on maize crops were monitored across three altitudinal categories with weeks after planting of maize to find that the highest population density and the percentage and intensity of FAW attack were conducted at an altitude of 500–1000 m a.s.l. at 2–4 weeks after planting [16]. However, accurately estimating the impact of FAW on maize growth, yield and quality under natural infestations remains challenging [14,17]. The effects of defoliation on maize growth and yields varies depending on the growth stage of the maize [18], where young maize plants at the 1st–2nd and 3rd–4th expanded leaf stages can compensate for 33–66% defoliation without reducing grain yields in New Mexico, USA [18]. However, kernel weight is particularly affected by ‘source’ manipulation during grain filling [19].
Therefore, in the present study, we simulated pest damage by applying varying degrees of defoliation at two growth stages across the three typical cropping seasons from 2021 to 2023. The objectives of these treatments were to evaluate how defoliation affects the quantity and quality of maize over these three cropping seasons spanning three years.
2. Materials and Methods
2.1. Study Site and Climatic Conditions
The study was conducted in the experimental fields of the Faculty of Agriculture, University of Miyazaki (31°49′39″ N, 131°24′46″ E, 27 m above sea level), from April 2021 to November 2023. Based on data collected by the Japan Meteorological Agency [20], daily mean temperature and cumulative precipitation in the growing season from April to November were 21.9 °C and 2752 mm, 22.6 °C and 2549 mm, and 22.3 °C and 2493 mm in 2021, 2022 and 2023, respectively, which was higher than the 30-year average of 21.8 °C and 2227 mm (Figure 1). In 2021, Typhoon No. 9 hit the area in early-mid August, causing abundant rainfall (>320 mm), and in mid-September of the same year, an active autumn rain front was associated with more than 560 mm of rainfall. The autumn temperature tended to be higher from mid-September to mid-October in 2021 when the 10-day mean temperature was above 22 °C. Typhoon No. 14 in 2022 hit Miyazaki in mid-September, causing rainfall to exceed 370 mm. In 2023, Typhoon No. 6 approached Miyazaki in early August, causing more than 450 mm of rainfall, and this was followed by more than 450 mm in early June due to the ‘Baiu’ rainy season. The field was flooding on the day of Typhoon No. 6, 9 August 2023, while the flooding water disappeared due to highly permeable volcanic ash soils on the following day, August 10. In the summer sowing maize season, the unusually high temperature of 26 °C extended from early August to late September. No symptom of drought damage was found in maize across three sowing seasons, fitting to the minimum requirement of precipitation in all growing seasons, every year.
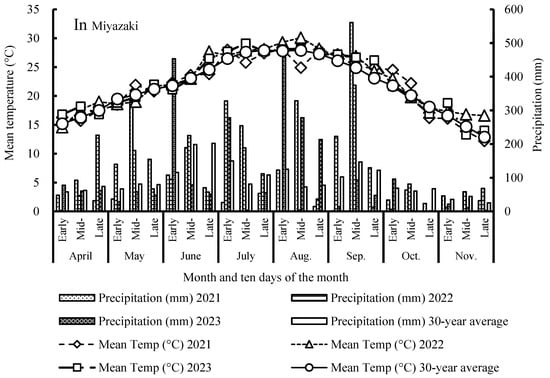
Figure 1.
Daily mean temperature and precipitation for 10-day intervals in the maize growing seasons from 2021 to 2023, together with the 30-year average from 1991 to 2020 for Miyazaki Prefecture.
2.2. Experimental Design, Treatments and Measurements
Maize crops were planted using a randomized block design comprising three replicate blocks (replicates), with each block subdivided into four plots measuring 3 × 5 m (15 m2) for each treatment. Each plot consisted of two rows of 3 × 1 m (3 m2), with a 3 × 1 m (3 m2) buffer between blocks, implemented over three cropping seasons from 2021 to 2023. The defoliation treatments were categorized into three levels: defoliation of three expanded leaves at the 3rd–4th leaf stage (DF1), defoliation of the 5th–6th expanded leaves by leaf punch to create three 15 mm × 4 mm holes on each side of the midrib (total of 6 holes, DF2) and defoliation of the expanding leaves together with the DF2 treatment at the 6th–7th leaf stage (DF3), with a control (Cont) featuring no treatment (Figure 2).

Figure 2.
Methods of defoliation treatments in (a) DF1 (defoliation of the 1st–3rd expanded leaves at the 3rd–4th leaf stage), (b) DF2 (defoliation of the 5th–6th expanded leaves by leaf punch at the 6th–7th leaf stage) and (c) DF3 (defoliation of the expanding leaves in addition to DF2 treatment at the 6th–7th leaf stage) and (d) the impact of FAW at the 6th–7th leaf stage in the field, and the experimental field in (e) DF1 plot (Center) at the 3rd–4th leaf stage, and (f) DF2 (Left) and DF3 (Right) plots at the 6th–7th leaf stage.
Figure 2d shows the severe impact of FAW on the summer sowing maize crops, monitored in Miyakonojo, Miyazaki (31.786° N, 131.053° E) on 1 September 2022, when all expanding leaves were lost by the feeding of FAW in addition to the typical insect bite marks along the midrib on the expanded leaves. Therefore, the defoliation treatment of DF3 was simulated for this worst impact case, that of DF2 was simulated for the case when the effective control of FAW damage was achieved by finding the FAW bites at the 5th–6th leaf stage (Figure 2f), and that of DF1 was tried to simulate the impact of FAW on the youngest seedling stage (Figure 2e), followed by the successful control achieved at the later growing stages.
Cropping seasons consisted of spring sowing in early to mid-April, late sowing in early June and summer sowing in early August, each maintained for three years. The maize varieties used were early-maturity LG3520, LG2008 and LG30500 for spring sowing in 2021, 2022 and 2023, respectively; late-maturity PI2008 consistently for late sowing; and late-maturity P3577, PI2008 and P3577 for summer sowing in 2021, 2022 and 2023, respectively. Maize planting employed two seeds per hole (0.2 × 0.5 m spacing) and thinned to 1 plant per hole at the 5th–6th leaf stage to maintain 80,000 plants ha−1.
The experimental site featured fine volcanic ash soils (Andosol). Before sowing the maize, the plots were rotary-plowed, and basal fertilization was performed using cattle manure at 30 Mg ha−1 and slaked lime at 1.5 Mg ha−1. Chemical fertilizers were applied in split doses, each at 100 kg ha−1 of N, P2O5 and K2O, during each growing season. Weeding was performed manually as required. No chemical insecticides were used throughout the three-year study period due to low pest incidence; however, dolomite fertilizer was applied to the whorl of maize in response to leaf damage at the vegetative stage to induce larval mortality.
Plant growth attributes, including height and the number of expanded leaves, were monitored at 2–6-week intervals at 11 plants per row (22 plants per treatment) for each treatment per replicate. At the silking stage in 2023, areas (cm2) of all live leaves were measured by portable leaf area meter (LI-3000C, LI-COR Corp., Lincoln, NE, USA) at one plant per replicate for the spring and summer sowings, except for late sowing due to lodging and leaf drops by Typhoon No. 6. Leaf area index (LAI) was calculated by summing up the areas of all live leaves, multiplied by planting density and divided by 10,000. At the milk-ripe or yellow-ripe stage of each crop, the fresh weights (FWs) of maize plants were measured for 5 plants per row (10 plants per treatment) per replicate. In addition, the average one plant per row (2 plants per treatment) per replicate was fractionated into constituent plant parts, such as leaves, stem inclusive leaf sheathes, husks and ears, and cut into 2 cm piece so that dry matter (DM) content could be determined. Material for DM measurements was dried in a forced-air oven at 70 °C for 3–4 days. Unfortunately, the 2021 spring sown crops could not be harvested due to wild animal (wild boars) invasion damage at the early maturity stage. At harvest, grain yield components, such as the number of grain rows, grain numbers per row and thousand grain weight were determined in only 2022 and 2023.
2.3. Forage Quality Analyses
Plant samples were systematically fractionated into distinct parts and dried at 70 °C for more than 3 days in a forced air oven. Once dried, plant fractions were ground to pass through a 1 mm sieve; samples were processed in duplicate for each replication. Acid detergent fiber (ADF) content was determined using a detergent method [21] and the ANKOM filter bag method [22] using an ANKOM-200 Fiber Analyzer (ANKOM Technology Co., Ltd., Macedon, NY, USA). The total digestible nutrient (TDN) content of maize was estimated from the ADF content using the following regression equation [23]:
TDN = 89.89 − 0.752 × ADF
In vitro dry matter (DM) digestibility (IVDMD) was determined by a pepsin-cellulase assay [24] using the filter-bag method and an in vitro incubator (ANKOM Technology Co., Ltd., Macedon, NY, USA).
2.4. Statistical Analyses
Statistical analyzes were performed to compare growth attributes, DM yield, IVDMD, ADF content and grain yield components for all treatments from 2021 to 2023. Differences in mean values between treatments were compared using a one-way analysis of variance, at 5% probability, by Fisher’s least significant difference test, using Excel Statistics 2020 software (Microsoft® Excel, Redmond, Seattle, WA, USA).
3. Results
3.1. Changes in Plant Height
Changes in plant height over time after sowing were analyzed across three cropping seasons and defoliation treatments (DF1, DF2 and DF3) and a no-treatment control (Cont) in 2022 (Figure 3). In the spring sowing season, defoliation (DF1) led to a decrease in plant height that did not recover by harvest (Figure 3a). Conversely, in the late sowing season (Figure 3b), the differences in plant height among defoliation treatments were less pronounced compared to the spring sowing, and a trend was similar in the summer sowing season (Figure 3c).
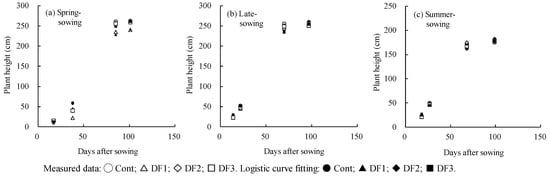
Figure 3.
Changes in plant height over time (days) after sowing and the logistic curve fitting (y = K/(1 + bexp(-ax))) between days after sowing (x) and plant height (y) across defoliation treatments (DF1, DF2 and DF3) and a no-treatment control (Cont) for three cropping seasons in 2022. Cropping seasons: spring sowing, planted in early April; late sowing, planted in early June; and summer sowing, planted in early August. Defoliation treatments: Cont (no treatment); DF1 (defoliation of the 1st–3rd expanded leaves at the 3rd–4th leaf stage); DF2 (defoliation of the 5th–6th expanded leaves by leaf punch at the 6th–7th leaf stage); and DF3 (defoliation of the expanding leaves in addition to DF2 treatment at the 6th–7th leaf stage).
Changes in plant height over days after sowing were modeled using a logistic curve, with results including multiple correlation coefficients and p values from 2022 and 2023 presented in Table 1. However, the modelling cannot apply to the results for plant height in 2021 due to there being few datasets. The multiple correlation coefficients were significant at the 5% level, indicating that the logistic curve effectively fits the changes in plant height as a function of days after sowing, except for the late sowing crops in 2023. This exception was due to significant disruption caused by heavy winds associated with Typhoon No. 6 in early August 2023. Notably, the K values, representing the carrying capacity of the logistic model, were lower in the DF1 treatment in 2022 for spring sowing, and in DF3 treatment in 2023 across the spring sowing, late sowing and summer sowing, compared to the Cont.

Table 1.
Logistic curve parameters (y = K/(1 + bexp(-ax))) between days after sowing (x) and plant height (y), multiple regression coefficients and p values for defoliation treatments across three cropping seasons in 2022 and 2023.
3.2. Plant Dry Matter Yields and Total Digestible Nutrient (TDN) Contents
Figure 4 shows plant DM yields and TDN contents at harvest for defoliation treatments (DF1, DF2 and DF3) and a no-treatment control (Cont) across three cropping seasons from 2021 to 2023. In the spring sowing season, DM yields decreased significantly in DF1 compared to Cont, followed by DF3, whereas the differences between Cont and DF2 were negligible. In the late sowing season, the variation in DM yield was diminished among treatments, except for the decrease in DF3 plot in 2021. In the summer sowing season, DM yields were the lowest in DF3, followed by DF2 and DF1. Differences in TDN content between treatments were not significant, ranging from 65% to 74% DM, while TDN contents tended to be lower in DF1 and DF3 for the summer sowing crops across the three years.
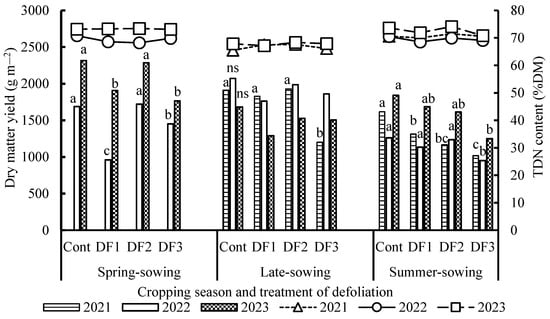
Figure 4.
Dry matter yield and total digestible nutrient (TDN) content at harvest for defoliation treatments across three cropping seasons, 2021−2023. For details on cropping seasons and defoliation treatments, refer to the footnote of Figure 3. Symbols followed by the same letter indicate no significant difference between the defoliation treatments within the same year at the 5% probability level according to Fisher’s least significant difference test. ns: not significant (p > 0.05).
3.3. Grain Yield and Grain Yield Components
Figure 5 shows the changes in grain yield and grain yield components at harvest under defoliation treatments (DF1, DF2 and DF3) and a no-treatment control (Cont) for three cropping seasons in 2022 and 2023. In the spring sowing season, significant reductions in grain yields were observed in DF1, followed by DF3, which corresponded to changes in DM yields. For the late sowing season, grain yields showed no significant differences between treatments in either year. In the summer sowing season, grain yields decreased significantly in DF3 in both years. The differences in grain yields among treatments were mainly attributed to differences in the number of grains per row, while the thousand grain weight did not differ significantly between treatments, except for a significant decrease in DF1 during the spring sowing season in 2022.
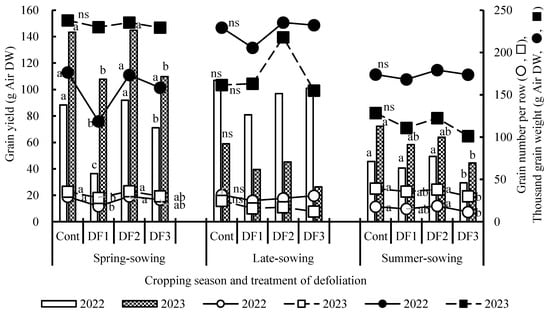
Figure 5.
Grain yield and grain yield components at harvest in the defoliation treatments for three cropping seasons, 2021–2023. For details on cropping seasons and defoliation treatments, refer to the footnote of Figure 3. Symbols followed by the same letter indicate no significant difference between defoliation treatments in the same year at the 5% probability level, according to Fisher’s least significant difference test. ns: not significant (p > 0.05).
3.4. Plant DM Yields, and TDN Yields in Relation to Plant Leaf Area at Silking Stage
Considering the relationship between ‘source’ and ‘sink’ in determining grain yields, plant DM yields and TDN yields were positively and significantly related with leaf area index (LAI) values at the silking stage in 2023, as shown in Figure 6. This correlation shows that reductions in LAI due to defoliation by insects lead to a linear decrease in both plant DM yields (a) and TDN yields (b).
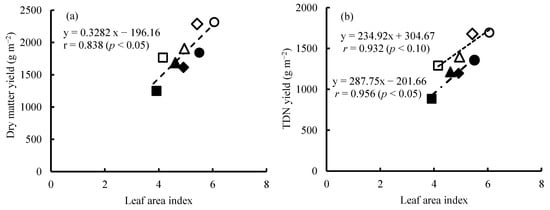
Figure 6.
Relationship between leaf area index at the silking stage and plant dry matter yield (a) and total digestible nutrient (TDN) yield (b) in 2023. Defoliation treatments: Cont (○, ●); DF1 (△, ▲); DF2 (◇, ◆); and DF3 (□, ■). Cropping seasons: spring sowing (○, △, ◇, □) and summer sowing (●, ▲, ◆, ■). Note: plant leaf area measurements were not possible for the late sowing season due to plant lodging caused by typhoon damage.
3.5. Relationship between Acid Detergent Fiber (ADF) Content and In Vitro Dry Matter Digestibility (IVDMD) at Harvest
Figure 7 shows the relationships between ADF content and IVDMD at harvest for three years. Plant IVDMD decreased with an increase in ADF content, resulting in significantly negative correlations between these two forage quality attributes. Consequently, an increase in defoliation intensity, which leads to an increase in ADF content, results in reduced digestibility of maize crops.
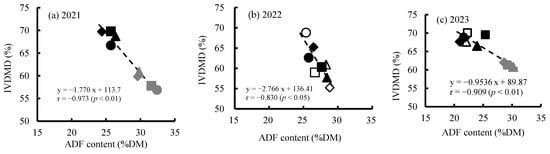
Figure 7.
Relationship between acid detergent fiber (ADF) content and in vitro dry matter digestibility (IVDMD) at harvest in 2021 (a), 2022 (b) and 2023 (c). Defoliation treatments: Cont (○, ●, ●); DF1 (△, ▲, ▲); DF2 (◇, ◆, ◆) and DF3 (□, ■, ■). Cropping seasons: spring sowing (○, △, ◇, □); late sowing (●, ▲, ◆, ■); and summer sowing (●, ▲, ◆, ■).
4. Discussion
Observations from Ecuador indicate that damage from the FAW on maize varies seasonally, with less damage occurring during the rainy season (>1700 mm of precipitation) compared to the dry season (<50 mm) [25]. In the present study, the rainfall distribution across the three cropping seasons was relatively uniform, except for the lower precipitation during the later growth stages of the summer sown crops. Consequently, it is important to consider the thermal response of maize growth across these seasons [11], especially given the marked increase in daily mean temperatures from early April through early or mid-August, as shown in Figure 1.
In the spring sowing crops, an increase in plant height was delayed in the DF1 treatment, followed by DF2, compared with the control (no-treatment), as shown in Figure 3a. Additionally, the variable K, representing the carrying capacity in the logistic curve fitting, was lower in DF1 than in the control for the spring sowing, and this pattern followed that observed for late sowing (Table 1). For the summer sowing crops, the rate of plant height increase did not vary significantly among treatments, although the K values were lower in DF3 than in the control for the 2023 season (Table 1). A significant decrease in plant height due to defoliation at the seedling stage (V4), comparable to that observed in the DF1 treatment in the present study, was observed in Liaoning, northeast China [26]. This reduction in height was beneficial in reducing lodging of maize crops caused by typhoons, due to shortening of the internode length, and led to a significant increase in grain yield [26].
In the present study, maize crops did not experience lodging, except during the late sowing in 2023 when severe lodging and leaf loss at the silking stage occurred due to Typhoon No. 6, irrespective of the defoliation treatments. Despite these conditions, total DM yield and TDN yield were positively correlated with LAI at the silking stage when the plant leaf area was at its maximum, as shown in Figure 6. These relationships correspond to the source manipulation to sink capacity during the grain-filling stage in maize [19,26]. Interestingly, a study in the USA found that grain yield was not adversely affected by early defoliation, even with leaf removal ranging from 0% to 66% at the 1st–2nd and the 3rd–4th leaf stages (V1–2 and V3–4, respectively) [18]. However, in the present study, maize plant height, DM and grain yields were reduced by DF1 for the spring sowing plots in 2022 and 2023. The discrepancy between the previous study [18] and the present study was derived from the lower growing temperature below 20 °C at the early vegetative stage in spring sowing maize for the current study. These findings provide a strong theoretical basis for recommending the application of insecticides at low levels of FAW infection during the seedling stages (V3–4). In the present study, defoliation treatments DF1 and DF2 for summer sowing crops did not decrease grain yields compared to the control in 2022–2023. Regarding the forage quality of maize, a previous study on spring sowing maize reported an increase in ADF content led to a decrease in IVDMD across several plant fractions over two years [12], suggesting a negative correlation between these two quality variables.
The efficacy of chemical pest control by insecticide application on maize yields varies markedly across different growth stages of maize, from 1 week (early vegetative stage) to 8 weeks (tasseling stage) after emergence. It has been established that maize plants protected earlier in the growth cycle exhibit higher grain yields compared to those treated at later growth stages [27]. This response to chemical control under high levels of natural pest pressure [27] contrasts with findings from a defoliation treatment study, which reported no decrease in grain yield [28]. However, it is critical to emphasize that early stage of protection against FAW infection is crucial for mitigating damage to maize crops by this highly invasive pest [2].
In addition to chemical sprays, seed treatments incorporating a combination of chemicals, such as Chlorantraniliprole, Cyantraniliprole and Thiodicarb with Imidacloprid, have proven effective for increasing FAW larval mortality above 50% and maintaining maize damage ratings below 5.0 for up to 14 days after emergence [29]. In Japan, seed treatment using the chemical ‘Lumivia TM FS’, which contains Chlorantraniliprole, was registered for use with maize on 23 August 2023 [30] and has been shown to be effective for controlling FAW on maize crops within two weeks after emergence.
After these early growth stages, the application of entomopathogenic nematodes mixed with sand [13] and biosilica fertilizer as foliar spray over the whorl area of maize crops at a rate of 2–3 L per ha three times from 10 days after emergence [31], has successfully reduced larval density [13], reduced leaf damage by FAW [13,31] and increased maize cob yields [13,31]. Consequently, by considering the variable costs and benefits associated with different protection strategies, it is advisable to adopt a multifaceted approach that integrates various protective measures, including biological maize hybrid selection [25], adjustments in cropping seasons and chemical techniques.
At the end of discussion section, the authors should address the limitation of the present experimental work on maize crops. For example, the influence of applying defoliation on maize was sometimes able to fully simulate the impact of FAW on the actual maize field, however, it may cause excessive stress on maize, compared with the actual FAW impact in the field. In addition, the authors faced to uncontrolled factors, such as damages by Typhoons and wild animal invasion, which blocked the completely paired comparison across three cropping seasons, even when three-year trials were accomplished.
5. Conclusions
The results of the present study suggest that defoliation due to insect damage, particularly at the early seedling stage in the spring sowing season and at the vegetative stage before stem elongation in the summer sowing season, is likely to reduce maize yield and forage quality. Consequently, it is advisable to apply registered insecticides at optimal growth stages of maize crops, specifically when insect damage is observed in the field. This timely intervention can help mitigate the adverse impacts on crop yield and quality.
Author Contributions
Conceptualization, Y.I.; methodology, Y.I. and M.N.; validation, K.T. and Y.I.; formal analysis, K.T.; investigation, K.T., M.I., S.M. and Y.I.; data curation, Y.I.; writing—original draft preparation, K.T. and Y.I.; writing—review and editing, M.N., M.T. and S.I.; visualization, Y.I.; supervision, T.A.-H.; project administration, T.A.-H.; funding acquisition, Y.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Livestock Promotional Subsidy from the Japan Racing Association.
Data Availability Statement
Data supporting the findings of this study are available from the corresponding author on reasonable request.
Acknowledgments
We are grateful to Yui Sakuraba for cooperation with the investigation during the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rane, R.; Walsh, T.K.; Lenancker, P.; Gock, A.; Dao, T.H.; Nguyen, V.L.; Khin, T.N.; Amalin, D.; Chittarath, K.; Faheem, M.; et al. Complex multiple introductions drive fall armyworm invasions into Asia and Australia. Sci. Rep. 2023, 13, 660. [Google Scholar] [CrossRef]
- Wan, J.; Huang, C.; Li, C.-Y.; Zhou, H.-X.; Ren, Y.-L.; Li, Z.-Y.; Xing, L.-S.; Zhang, B.; Qiao, X.; Liu, B.; et al. Biology, invasion and management of the agricultural invader: Fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Integr. Agric. 2021, 20, 646–663. [Google Scholar] [CrossRef]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread and impact of fall armyworm (Spodoptera frugiperda J. E. Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 2020, 292, 106804. [Google Scholar] [CrossRef]
- Girsang, S.S.; Nurzannah, S.E.; Girsang, M.A.; Effendi, R. The distribution and impact of fall army worm (Spodoptera frugiperda) on maize production in North Sumatera. IOP Conf. Ser. Earth Environ. Sci. 2020, 484, 012099. [Google Scholar] [CrossRef]
- Caasi-Lit, M.T.; Marmeto, A.M.D. Maize response to fall armyworm (Spodoptera frugiperda) and Asian corn borer (Ostrinia furnacalis) in the Philippines. SABRAO J. Breed. Genet. 2022, 54, 1231–1240. [Google Scholar] [CrossRef]
- Dahi, H.F.; Salem, S.A.R.; Gamil, W.E.; Mohamed, H.O. Heat requirements for the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) as a new invasive pest in Egypt. Egypt. Acad. J. Biol. Sci. A Entomol. 2020, 13, 73–85. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef]
- Otsuka, A. Prediction of the overseas migration of the fall armyworm, Spodoptera frugiperda, to Japan. Insects 2023, 14, 804. [Google Scholar] [CrossRef]
- Fan, Z.; Song, Y.; Zhao, S.; Wu, K. Invasion of fall armyworm led to the succession of maize pests in Southwest China. J. Integr. Agric. 2024, 23, 1300–1314. [Google Scholar] [CrossRef]
- Ministry of Agriculture, Forestry and Fisheries of Japan, MAFF. Outbreak of Fall Armyworm in 2020 (as of 6 November 2020). Available online: https://www.maff.go.jp/j/syouan/syokubo/keneki/k_kokunai/attach/pdf/tumajiro-146.pdf (accessed on 13 April 2024).
- Yang, Y.; Ishii, Y.; Idota, S. Year-round forage yield stability through a system combining triple-maize crops with winter barley in Kyushu, Japan. Am. J. Agric. Biol. Sci. 2016, 11, 19–28. [Google Scholar] [CrossRef][Green Version]
- Li, B.; Ishii, Y.; Idota, S.; Tobisa, M.; Niimi, M.; Yang, Y.; Nishimura, K. Yield and quality of forages in a triple cropping system in southern Kyushu, Japan. Agronomy 2019, 9, 277. [Google Scholar] [CrossRef]
- Ratnakala, B.; Kalleshwaraswamy, C.M.; Rajkumar, M.; Deshmukh, S.S.; Mallikarjuna, H.B.; Narasimhaiah, L. Field evaluation of whorl application of sand mixed entomopathogenic nematodes for the management of invasive fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in sweet corn. Egypt. J. Biol. Pest Control 2023, 33, 58. [Google Scholar] [CrossRef]
- Toepfer, S.; Fallet, P.; Kajuga, J.; Bazagwira, D.; Mukundwa, I.P.; Szalai, M.; Turlings, T.C.J. Streamlining leaf damage rating scales for the fall armyworm on maize. J. Pest Sci. 2021, 94, 1075–1089. [Google Scholar] [CrossRef]
- Soujanya, P.L.; Sekhar, J.C.; Yathish, K.R.; Karjagi, C.G.; Rao, K.S.; Suby, S.B.; Jat, S.L.; Kumar, B.; Kumar, K.; Vadessery, J.; et al. Leaf damage based phenotyping technique and its validation against fall armyworm, Spodoptera frugiperda (J. E. Smith), in maize. Front. Plant Sci. 2022, 13, 906207. [Google Scholar] [CrossRef]
- Supartha, I.W.; Susila, I.W.; Yuliadhi, K.A.; Haloho, E.S.M.; Yudha, I.K.W.; Utama, I.W.E.K.; Wiradana, P.A. Monitoring of damage and distribution of invasive fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on maize crop in Karo, North Sumatera, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2022, 951, 012010. [Google Scholar] [CrossRef]
- Chisonga, C.; Chipabika, G.; Philemon, H.; Sohati, P.H.; Harrison, R.D. Understanding the impact of fall armyworm (Spodoptera frugiperda J. E. Smith) leaf damage on maize yields. PLoS ONE 2023, 18, e0279138. [Google Scholar] [CrossRef]
- Blanco, C.A.; Conover, K.; Hernandez, G.; Valentini, G.; Portilla, M.; Abel, C.A.; Williams, P.; Nava-Camberos, U.; Hutchison, W.D.; Dively, G.P. Grain yield is not impacted by early defoliation of maize: Implications for fall armyworm action thresholds. Southwest. Entomol. 2022, 47, 335–344. [Google Scholar] [CrossRef]
- Echarte, L.; Andrade, F.H.; Sadras, V.O.; Abbate, P. Kernel weight and its response to source manipulations during grain filling in Argentinean maize hybrids released in different decades. Field Crops Res. 2006, 96, 307–312. [Google Scholar] [CrossRef]
- Japan Meteorological Agency. Meteorological Statistical Data for Miyazaki, Miyazaki Prefecture. Available online: https://www.jma.go.jp/jma/menu/report.html (accessed on 13 April 2024).
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- ANKOM Technology Corporation. ANKOM—Automation Made Simple; ANKOM Technology: Macedon, NY, USA; Available online: https://www.ankom.com/ (accessed on 13 April 2024).
- Committee for the Nutritive Value of Grass and Forage Crops. The Guidebook of Nutritive Values of Herbage and Forage Crop; Hokunoukai: Sapporo, Japan, 1991; pp. 41–45. [Google Scholar]
- Goto, I.; Minson, D.J. Prediction of the dry matter digestibility of tropical grasses using a pepsin-cellulase assay. Anim. Feed Sci. Technol. 1977, 2, 247–253. [Google Scholar] [CrossRef]
- Briones Ochoa, M.A.; Sánchez-Mora, F.D.; Chirinos Torres, D.T. Can fall armyworm damage decrease depending on the season, maize hybrid, and type of pesticides? Sci. Agropecu. 2023, 14, 313–320. [Google Scholar] [CrossRef]
- Xu, J.; Zou, X.; Xu, H.; Gong, L.; Sun, Z.; Zhang, L.; Niu, S.; Feng, L.; Han, L.; Wang, R. Defoliation at seedling stage enhances maize yield by reducing lodging. Agron. J. 2023, 115, 544–556. [Google Scholar] [CrossRef]
- Van den Berg, J.; Britz, C.; du Plessis, H. Maize yield responses to chemical control of Spodoptera frugiperda at different plant growth stages in South Africa. Agriculture 2021, 11, 826. [Google Scholar] [CrossRef]
- Kaewchuai, S.; Thobunluepop, P.; Nakasathien, S.; Lertmongkol, S.; Onwimol, D.; Chitbanchong, W.; Bredemeier, M. Management of source-sink balance for maintaining seed vigor and storability of maize. Online J. Biol. Sci. 2021, 21, 199–205. [Google Scholar] [CrossRef]
- Oliveira, C.; Orozco-Restrepo, S.M.; Alves, A.C.L.; Pinto, B.S.; Miranda, M.S.; Barbosa, M.H.P.; Picanço, M.C.; Pereira, E.J.G. Seed treatment for managing fall armyworm as a defoliator and cutworm on maize: Plant protection, residuality, and the insect life history. Pest Manag. Sci. 2022, 78, 1240–1250. [Google Scholar] [CrossRef]
- Corteva, Agriscience, Japan. LumiviaTM FS. Available online: https://www.corteva.jp/products-and-solutions/crop-protection/lumivia-fs.html (accessed on 16 April 2024).
- Ulina, E.S.; Manurung, E.D.; Hasibuan, M.; Nasution, L.Z. Biosilica fertilizer reduces fall armyworm damage. IOP Conf. Ser. Earth Environ. Sci. 2022, 985, 012049. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).