
Non-Herbivore-Induced Plant Organic Volatiles of Tomato Cultivars and Their Effect on Pest Biological Control

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Insects
2.3. Olfactory Response of T. achaeae and N. tenuis to Different Tomato Cultivars
2.4. Bioassay Procedure—T. achaeae
2.5. Bioassays Procedure—N. tenuis
2.6. Volatile Collection
2.7. Tomato Cultivar Effects on T. achaeae and N. tenuis Population Dynamics
2.8. Statistical Analysis
3. Results
3.1. Olfactory Response of T. achaeae and N. tenuis to Different Tomato Varieties
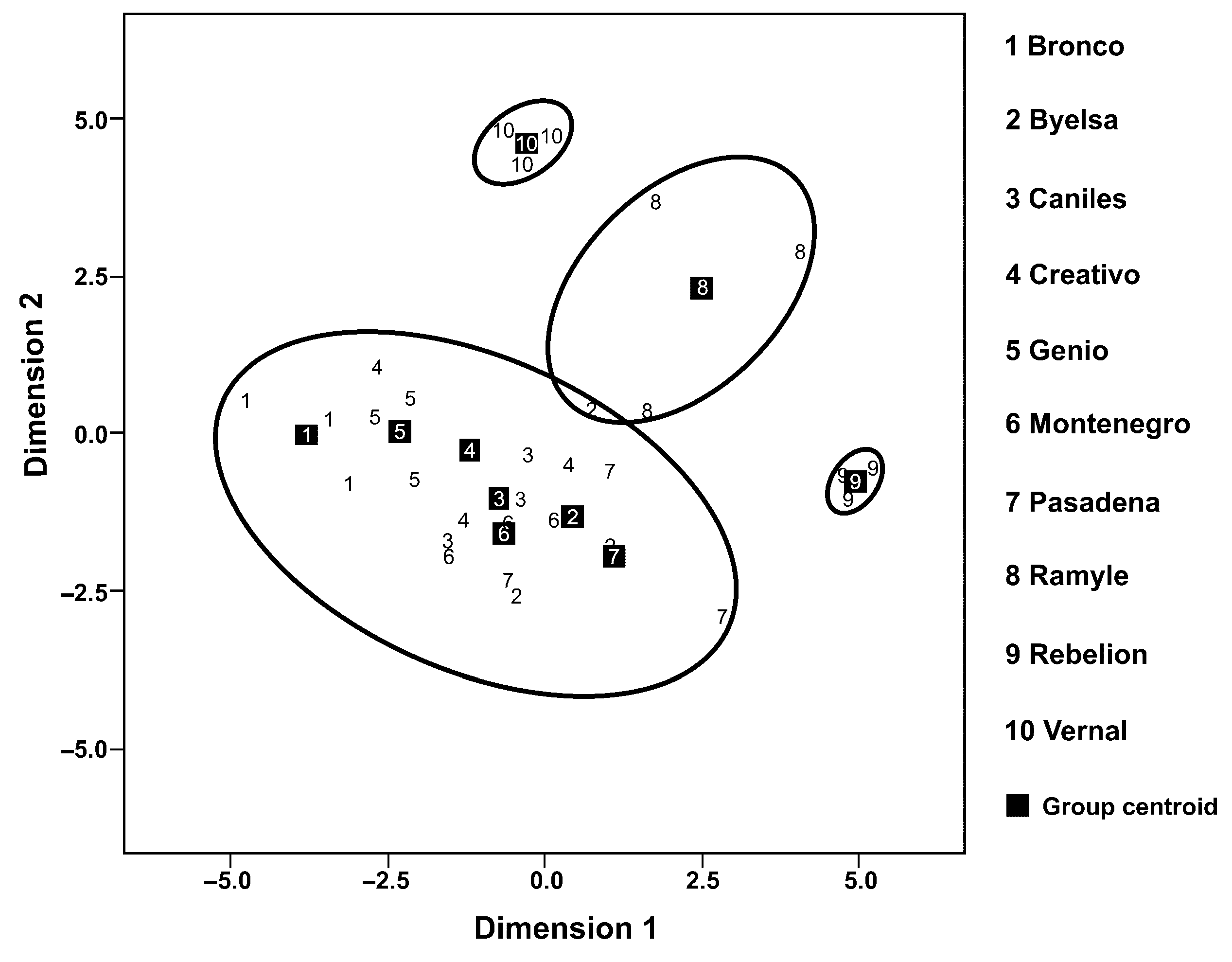
3.2. Volatile Collection
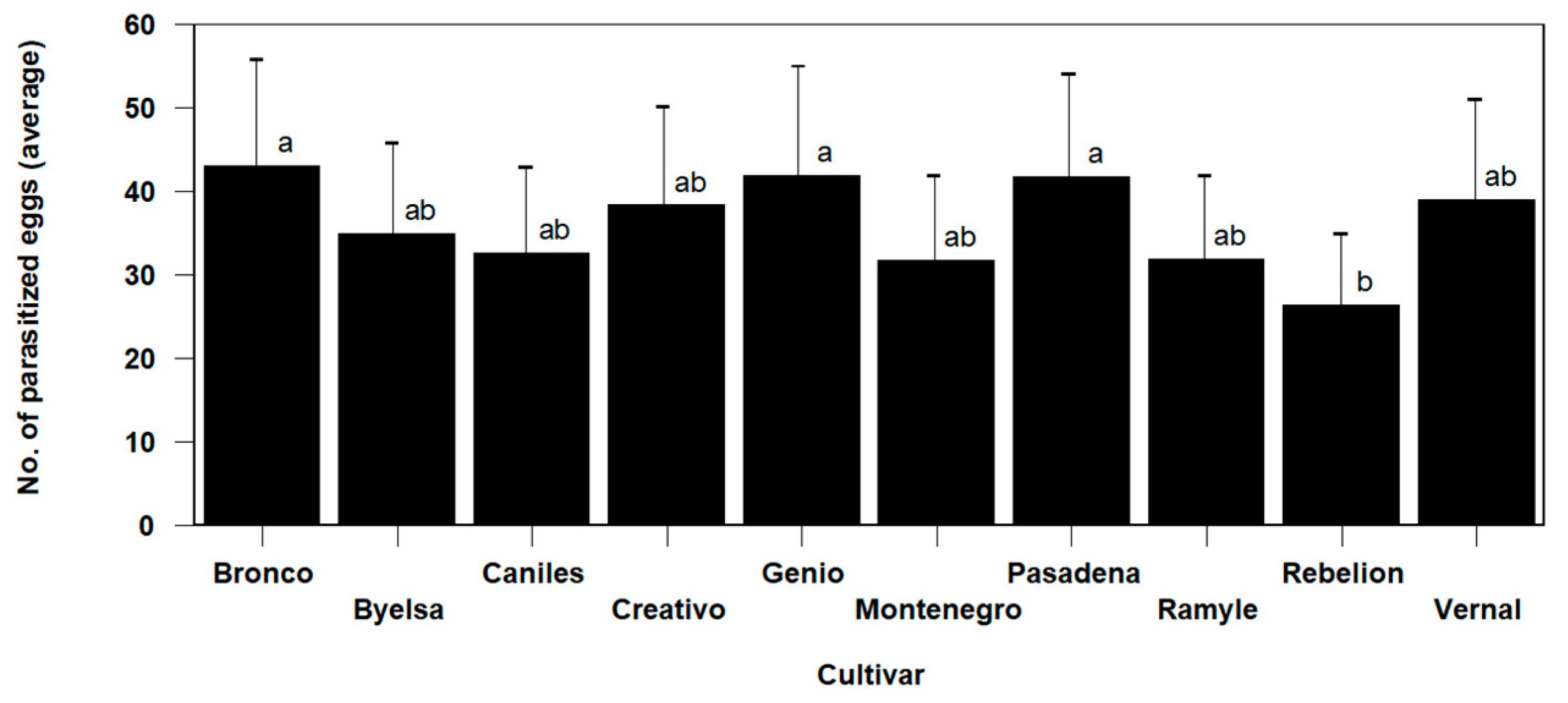
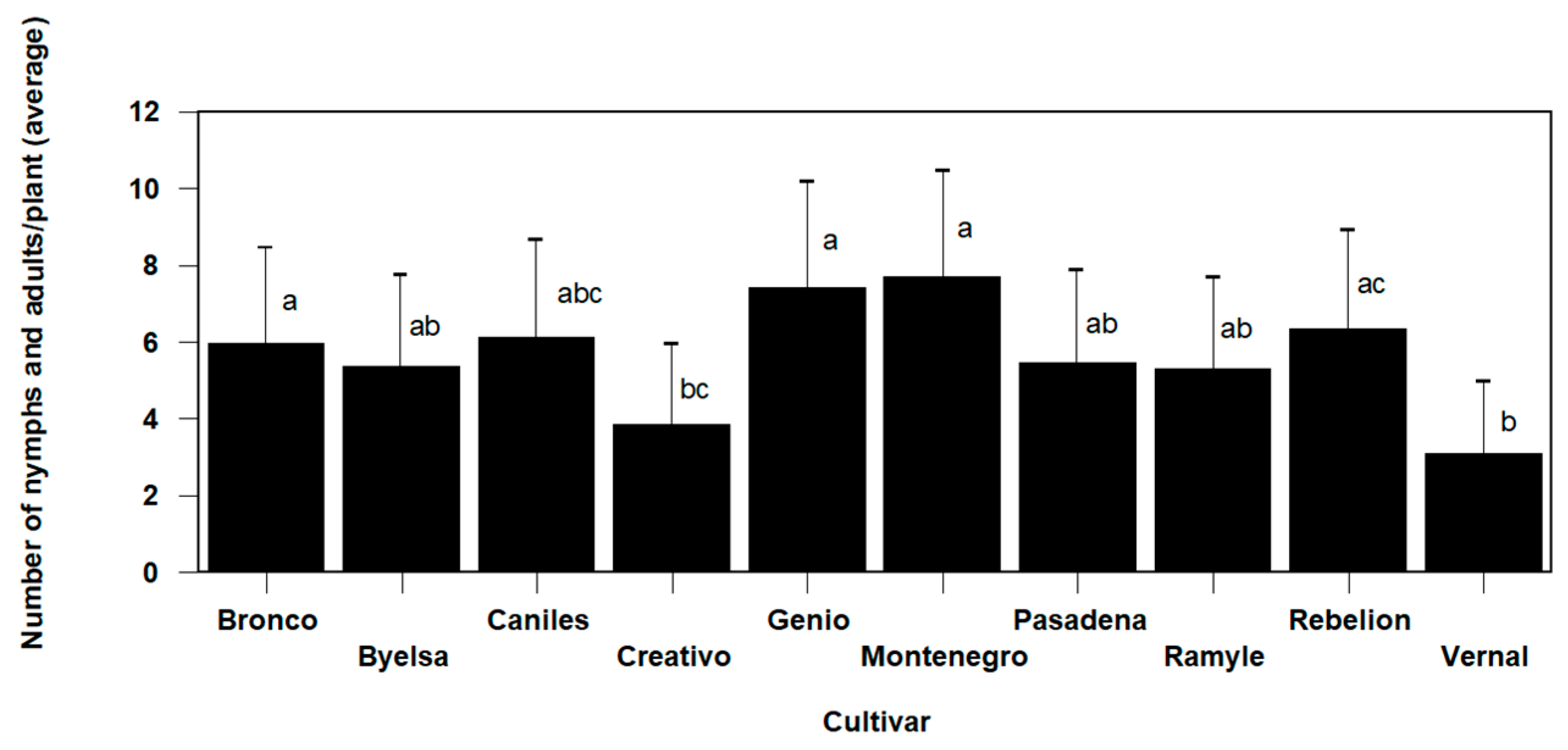
3.3. Tomato Cultivar Effects on T. achaeae and N. tenuis Population Dynamic
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brilli, F.; Loreto, F.; Baccellin, I. Exploiting plant volatile organic compounds (VOCs) in Agriculture to improve sustainable defence strategies and productivity of crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Ode, P.J. Plant chemistry and natural enemy fitness: Effects on herbivore and natural enemy interaction. Annu. Rev. Entomol. 2006, 51, 163–185. [Google Scholar] [CrossRef] [PubMed]
- Fellowes, M.D.E.; van Alphen, J.M.; Shameer, K.S. Foraging behaviour. In Jervis’s Insects as Natural Enemies: Practical Perspectives, 3rd ed.; Hardy, I.C.W., Wajnberg, E., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 1–101. [Google Scholar]
- Razo-Belman, R.; Ozuna, C. Volatile organic compounds: A review of their current applications as pest biocontrol and disease management. Horticulturae 2023, 9, 441. [Google Scholar] [CrossRef]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Bleeker, P.M.; Van Wijk, M.; Schuurink, R.C.; Haring, M.A. Plant volatiles in defence. Adv. Bot. Res. 2009, 51, 613–666. [Google Scholar] [CrossRef]
- Shrivastave, G.; Rogers, M.; Wszelaki, A.; Panthee, D.R.; Chen, F. Plant volatiles-based insect pest management in organic farming. Crit. Rev. Plant Sci. 2010, 29, 123–133. [Google Scholar] [CrossRef]
- Takabayashi, J.; Dicke, M. Plant—Carnivore mutualism through herbivore-induced carnivore attractants. Trends Plant Sci. 1996, 1, 109–113. [Google Scholar] [CrossRef]
- Ferreira, V.; Lopez, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Hoballah, M.E.F.; Tamo, C.; Turlings, T.C. Differential attractiveness of induced odors emitted by eight maize varieties for the parasitoid Cotesia marginiventris: Is quality or quantity important? J. Chem. Ecol. 2002, 28, 951–968. [Google Scholar] [CrossRef]
- Kappers, I.F.; Hoogerbrugge, H.; Bouwmeester, H.J.; Dicke, M. Variation in herbivory-induced volatiles among cucumber (Cucumis sativus L.) varieties has consequences for the attraction of carnivorous natural enemies. J. Chem. Ecol. 2011, 37, 150–160. [Google Scholar] [CrossRef]
- Kappers, I.F.; Verstappen, F.W.; Luckerhoff, L.L.; Bouwmeester, H.J.; Dicke, M. Genetic variation in jasmonic acid-and spider mite-induced plant volatile emission of cucumber accessions and attraction of the predator Phytoseiulus persimilis. J. Chem. Ecol. 2010, 36, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Duffey, S.S.; Stout, M.J. Antinutritive and toxic components of plant defense against insects. Arch. Insect Biochem. Physiol. 1996, 32, 3–37. [Google Scholar] [CrossRef]
- Glas, J.J.; Schimmel, B.C.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef]
- Scott, J.; Gardner, R. Breeding for resistance to fungal pathogens. In Genetic Improvement of Solanaceous Crops; Publ. Science: Enfield, NH, USA, 2007; pp. 421–456. [Google Scholar]
- Cortesero, A.; Stapel, J.; Lewis, W. Understanding and manipulating plant attributes to enhance biological control. Biol. Control 2000, 17, 35–49. [Google Scholar] [CrossRef]
- Hare, J.D. Plant genetic variation in tritrophic interactions. In Multitrophic Level Interactions; Cambridge University Press: Cambridge, UK, 2002; pp. 8–43. [Google Scholar]
- Vet, L.E.; Dicke, M. Ecology of infochemical use by natural enemies in a tritrophic context. Annu. Rev. Entomol. 1992, 37, 141–172. [Google Scholar] [CrossRef]
- Calvo, F.J.; Lorente, M.J.; Stansly, P.A.; Belda, J.E. Preplant release of Nesidiocoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisa tabaci in greenhouse tomato. Entomol. Exp. Appl. 2012, 143, 111–119. [Google Scholar] [CrossRef]
- Sanchez, C.; Gamez, M.; Burguilo, F.J.; Garay, J.; Cabello, T. Comparison of predator-parasitoid-prey interaction models for different host plant qualities. Community Ecol. 2018, 19, 125–132. [Google Scholar] [CrossRef]
- Cabello, T. Biologia de dos Especies de Trichogramma (Hym.: Trichogrammatidae) Parasitas de Helicoverpa spp. (Lep.: Noctuidae) en Algodonero: Posibilidades de su Empleo como Agentes de Control Biológico. Ph.D. Thesis, Universidad de Córdoba, Córdoba, Spain, 1985. [Google Scholar]
- Cabello, T.; Vargas, P. Estudio con olfactometro de la influencia de la planta y del insecto huesped en la actividad de busqueda de Trichogramma cordubensis y T. sp. p. buesi (Hym.: Trichogrammatidae). Bol. Sanid. Veg. Plagas 1985, 11, 237–241. [Google Scholar]
- Vet, L.E.; Lenteren, J.V.; Heymans, M.; Meelis, E. An airflow olfactometer for measuring olfactory responses of hymenopterous parasitoids and other small insects. Physiol. Entomol. 1983, 8, 97–106. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Cooley, S.S.; Phelan, P.L. Bioassay approaches to assessing behavioral responses of plum curculio adults (Coleoptera: Curculionidae) to host fruit odor. J. Chem. Ecol. 1995, 21, 1073–1084. [Google Scholar] [CrossRef]
- Van Tol, R.; Visser, J.; Sabelis, M. Olfactory responses of the vine weevil, Otiorhynchus sulcatus, to tree odours. Physiol. Entomol. 2002, 27, 213–222. [Google Scholar] [CrossRef]
- Van Tol, R.; Visser, J.; Sabelis, M. Behavioural responses of the vine weevil, Otiorhynchus sulcatus, to semiochemicals from conspecifics, Otiorhynchus salicicola, and host plants. Entomol. Exp. Appl. 2004, 110, 145–150. [Google Scholar] [CrossRef]
- Pettersson, J. An aphid sex attractant. Insect Syst. Evol. 1970, 1, 63–73. [Google Scholar] [CrossRef]
- Sanchez, C.; Gallego, J.R.; Gamez, M.; Cabello, T. Intensive Biological Control in Spanish Greenhouses. Problems of the Success. Int. J. Agric. Biosyst. Eng. 2014, 8, 1123–1127. Available online: https://Waset.org/Publication/9999557 (accessed on 19 February 2024).
- Cabello, T.; Gallego, J.R.; Fernandez, F.J.; Gamez, M.; Vila, E.; Del Pino, M.; Hernandez–Suarez, E. Biological control strategies for the South American tomato moth (Lepidoptera: Gelechiidae) in greenhouse tomatoes. J. Econ. Entomol. 2012, 105, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Buttery, R.G.; Ling, L.C.; Light, D.M. Tomato leaf volatile aroma components. J. Agric. Food Chem. 1987, 35, 1039–1042. [Google Scholar] [CrossRef]
- Buttery, R.G.; Ling, L.C. Volatile components of tomato fruit and plant parts: Relationship and biogenesis. In Bioactive Volatile Compounds from Plant; Teranishi, R., Buttery, R.G., Sugisawa, H., Eds.; American Chemical Society: New York, NY, USA, 1993; Volume 525, pp. 23–34. [Google Scholar]
- Proffit, M.; Birgersson, G.; Bengtsson, M.; Reis, R.; Witzgall, P.; Lima, E. Attraction and oviposition of Tuta absoluta females in response to tomato leaf volatiles. J. Chem. Ecol. 2011, 37, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Vila, E.; Cabello, T. Lucha biologica: Utilizacion de entomofagos en cultivos en invernaderos. Agricultura 2012, 81, 10–15. [Google Scholar]
- Hendriks, M.M.; Cruz-Juarez, L.; De Bont, D.; Hall, R.D. Preprocessing and exploratory analysis of chromatographic profiles of plant extracts. Anal. Chim. Acta 2005, 545, 53–64. [Google Scholar] [CrossRef]
- Cusumano, A.; Weldegergis, B.T.; Colazza, S.; Dicke, M.; Fatouros, N.E. Attraction of egg-killing parasitoids toward induced plant volatiles in a multi-herbivore context. Oecologia 2015, 179, 163–174. [Google Scholar] [CrossRef]
- Lins, J.C.; van Loon, J.J.; Bueno, V.H.; Lucas-Barbosa, D.; Dicke, M.; van Lenteren, J.C. Response of the zoophytophagous predators Macrolophus pygmaeus and Nesidiocoris tenuis to volatiles of uninfested plants and to plants infested by prey or conspecifics. BioControl 2014, 59, 707–718. [Google Scholar] [CrossRef]
- Visser, J. Host-plant finding by insects: Orientation, sensory input and search patterns. J. Insect Physiol. 1988, 34, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Okubo, N.; Tanaka, F. Analysis of internal pools of plant volatiles. In Biology of Plant Volatiles, 2nd ed.; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 27–38. [Google Scholar]
- Gasparini, K.; Gasparini, J.; Therezan, R.; Vicente, M.H.; Sakamoto, T.; Figueira, A.; Zsögön, A.; Peres, L.E. Natural genetic variationin the hairs absent (H) gene increases type-VI glandular trichomes in both wild and domesticated tomatoes. J. Plant Physiol. 2023, 280, 153859. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Garvey, M.; Kaplan, I. Domestication of tomato has reduced the attraction of herbivores natural enemies to pest-damage plants. Agric. For. Entomol. 2018, 20, 390–401. [Google Scholar] [CrossRef]
- Kennedy, G.G. Tomato, pests, parasitoids, and predators: Tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol. 2023, 48, 51–72. [Google Scholar] [CrossRef]
- Romeis, J.; Babendreier, D.; Wackers, F.L.; Shanower, T.G. Habitat and plant specificity of Trichogramma egg parasitoids—Underlying mechanisms and implications. Basic Appl. Ecol. 2005, 6, 215–236. [Google Scholar] [CrossRef]
- Nordlund, D.A.; Chalfant, R.B.; Lewis, W. Response of Trichogramma pretiosum females to volatile synomones from tomato plants. J. Entomol. Sci. 1985, 20, 372–376. [Google Scholar] [CrossRef]
- Raghava, T.; Ravikumar, P.; Hegde, R.; Kush, A. Spatial and temporal volatile organic compound response of select tomato cultivars to herbivory and mechanical injury. Plant Sci. 2010, 179, 520–526. [Google Scholar] [CrossRef]
- Romeis, J.; Shanower, T.G.; Zebit, C.P.W. A simple and effective bioassay to study the effect of plant surface chemicals on the behavior of Trichogramma spp. Z. Pflanzenkr. Pflanzenschutz 1996, 103, 213–216. [Google Scholar]
- Romeis, J.; Shanower, T.; Zebitz, C. Physical and chemical plant characters inhibiting the searching behaviour of Trichogramma chilonis. Entomol. Exp. Appl. 1998, 87, 275–284. [Google Scholar] [CrossRef]
- Bar, D.; Gerling, D.; Rossler, Y. Bionomics of the principal natural enemies attacking Heliothis armigera in cotton fields in Israel. Environ. Entomol. 1979, 8, 468–474. [Google Scholar] [CrossRef]
- Tandon, P.; Bakthavatsalam, N. Plant volatile diversity in different tomato genotypes and its influence on parasitization efficiency of Trichogramma chilonis Ishii on Helicoverpa armigera (Hübner). J. Biol. Control 2007, 21, 271–281. [Google Scholar]
- Glinwood, R.; Ahmed, E.; Qvarfordt, E.; Ninkovic, V. Olfactory learning of plant genotypes by a polyphagous insect predator. Oecologia 2011, 166, 637–647. [Google Scholar] [CrossRef]
- Bjorksten, T.A.; Hoffmann, A.A. Separating the effects of experience, size, egg load, and genotype on host response in Trichogramma (Hymenoptera: Trichogrammatidae). J. Insect Behav. 1998, 11, 129–148. [Google Scholar] [CrossRef]
- Kauffman, W.C.; Kennedy, G.G. Relationship between trichome density in tomato and parasitism of Heliothis spp. (Lepidoptera: Noctuidae) eggs by Trichogramma spp. (Hymenoptera: Trichogrammatidae). Environ. Entom. 1989, 18, 698–704. [Google Scholar] [CrossRef]
- Naselli, M.; Zappala, L.; Gugliuzzo, A.; Garzia, G.T.; Biondi, A.; Rapisarda, C.; Cincotta, F.; Condurso, C.; Verzera, A.; Siscaro, G. Olfactory response of the zoophytophagous mirid Nesidiocoris tenuis to tomato and alternative host plants. Arthropod Plant Interact. 2017, 11, 121–131. [Google Scholar] [CrossRef]
- Urbaneja, A.; Tapia, G.; Stansly, P. Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae). Biocontrol Sci. Technol. 2005, 15, 513–518. [Google Scholar] [CrossRef]
- Loughrin, J.H.; Manukian, A.; Heath, R.R.; Tumlinson, J.H. Volatiles emitted by different cotton varieties damaged by feeding beet armyworm larvae. J. Chem. Ecol. 1995, 21, 1217–1227. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Genetic Background | Supplier (Location) |
|---|---|---|
| Bronco® | F1 Hybrid, cocktail type | Ramiro Arnedo Semillas (Calahorra, Spain) |
| Byelsa® | F1 Hybrid, plum type | Semillas Fitó (Barcelona, Spain) |
| Caniles® | F1 Hybrid, plum type | Zeraim Iberica, S.A. (Valencia, Spain) |
| Creativo® | F1 Hybrid, cherry type | Hm Clause (Portes-les-Valence, France) |
| Genio® | F1 Hybrid, cherry type | Hm Clause (Portes-les-Valence, France) |
| Montenegro® | F1 Hybrid, beef type | Rijk Zwaan Iberica, S.A. (Almeria, Spain) |
| Pasadena® | F1 Hybrid, beef type | Ramiro Arnedo Semillas (Calahorra, Spain) |
| Ramyle® | F1 Hybrid, on the vine type | Rijk Zwaan Iberica, S.A. (Almeria, Spain) |
| Rebelion® | F1 Hybrid, Marmande type | Vilmorin-Mikado Iberica, S.A. (Alicante, Spain) |
| Vernal® | F1 Hybrid, beef type | Enza Zaden España, S.L. (Almeria, Spain) |
| Trial | Cultivar | Average (*) | SE (±) |
|---|---|---|---|
| 1 | Bronco | 5.66 c | 1.37 |
| Byelsa | 5.99 c | 1.41 | |
| Caniles | 5.99 c | 1.41 | |
| Creativo | 9.66 b | 1.79 | |
| Montenegro | 16.98 a | 2.38 | |
| Rebelion | 2.00 d | 0.81 | |
| Vernal | 10.66 b | 1.88 | |
| Control | 0.67 d | 0.47 | |
| 2 | Bronco | 9.00 b | 1.00 |
| Byelsa | 6.67 b | 1.45 | |
| Genio | 19.67 a | 2.56 | |
| Pasadena | 1.67 c | 0.75 | |
| Ramyle | 2.67 c | 0.94 | |
| Rebelion | 1.33 c | 0.67 | |
| Vernal | 6.67 a | 1.49 | |
| Control | 1.33 bc | 0.67 |
| Trial | Cultivar | Average (*) | SE (±) |
|---|---|---|---|
| 1 | Bronco | 27.48 a | 5.73 |
| Byelsa | 21.11 a | 4.40 | |
| Ramyle | 23.70 a | 5.57 | |
| Vernal | 24.70 a | 5.15 | |
| 2 | Caniles | 21.72 b | 3.31 |
| Montenegro | 17.79 b | 2.71 | |
| Rebelion | 38.67 a | 5.88 | |
| Vernal | 21.81 b | 3.32 | |
| 3 | Creativo | 30.93 a | 6.56 |
| Genio | 28.33 a | 6.01 | |
| Pasadena | 27.78 a | 5.89 | |
| Vernal | 22.41 a | 4.75 |
| Cultivar | Average (*) | SE (±) |
|---|---|---|
| Bronco | 532.7 c | 114.6 |
| Byelsa | 2218.3 bc | 477.1 |
| Caniles | 2987.0 abc | 642.4 |
| Creativo | 532.7 c | 114.6 |
| Genio | 678.7 bc | 146.0 |
| Montenegro | 1312.7 bc | 282.3 |
| Pasadena | 3621.7 abc | 778.9 |
| Ramyle | 3979.7 abc | 855.9 |
| Rebelion | 6930.0 a | 1490.4 |
| Vernal | 5031.7 ab | 1082.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabello, T.; Gamez, M.; Gallego, J.R.; Lopez, I.; Sanchez, C.; Garay, J. Non-Herbivore-Induced Plant Organic Volatiles of Tomato Cultivars and Their Effect on Pest Biological Control. AgriEngineering 2024, 6, 1497-1510. https://doi.org/10.3390/agriengineering6020085
Cabello T, Gamez M, Gallego JR, Lopez I, Sanchez C, Garay J. Non-Herbivore-Induced Plant Organic Volatiles of Tomato Cultivars and Their Effect on Pest Biological Control. AgriEngineering. 2024; 6(2):1497-1510. https://doi.org/10.3390/agriengineering6020085
Chicago/Turabian StyleCabello, Tomas, Manuel Gamez, Juan Ramón Gallego, Inmaculada Lopez, Carolina Sanchez, and Jozsef Garay. 2024. "Non-Herbivore-Induced Plant Organic Volatiles of Tomato Cultivars and Their Effect on Pest Biological Control" AgriEngineering 6, no. 2: 1497-1510. https://doi.org/10.3390/agriengineering6020085