
Influence of Temperature and LED Light Spectra on Flavonoid Contents in Poa pratensis



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Transplantation
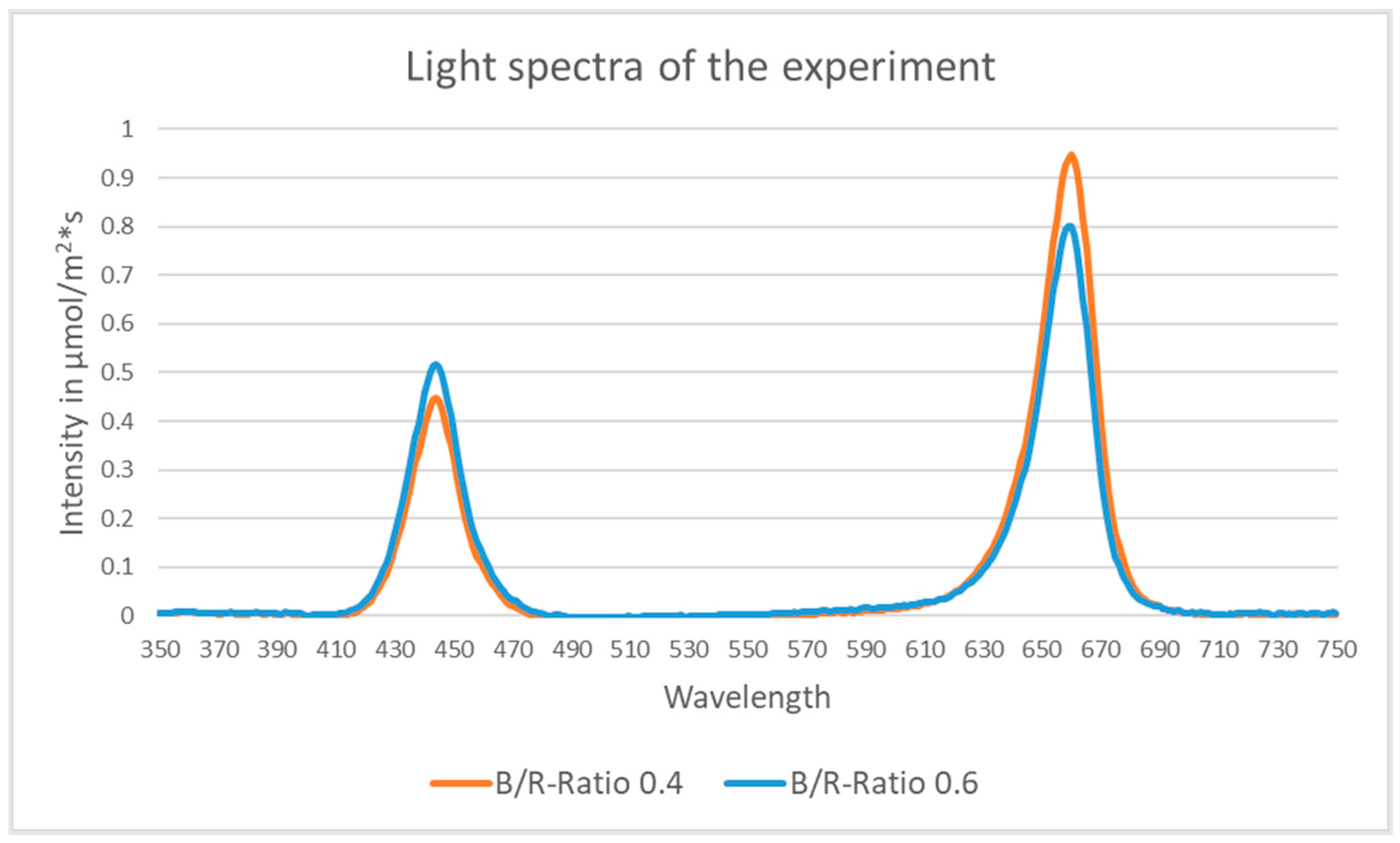
2.2. Experimental Design and Treatments
- Constant temperature of 4 °C over five weeks with three days of acclimatization (4 °C);
- Constant temperature of 12 °C over five weeks with three days of acclimatization (12 °C);
- Weekly temperature changes over five weeks, 12–8–4–8–12 °C, with three days of acclimatization (12 °C).
2.3. Internal Parameter Profile
2.4. Length Increase at Switching Temperature
2.5. Data Analysis
3. Results
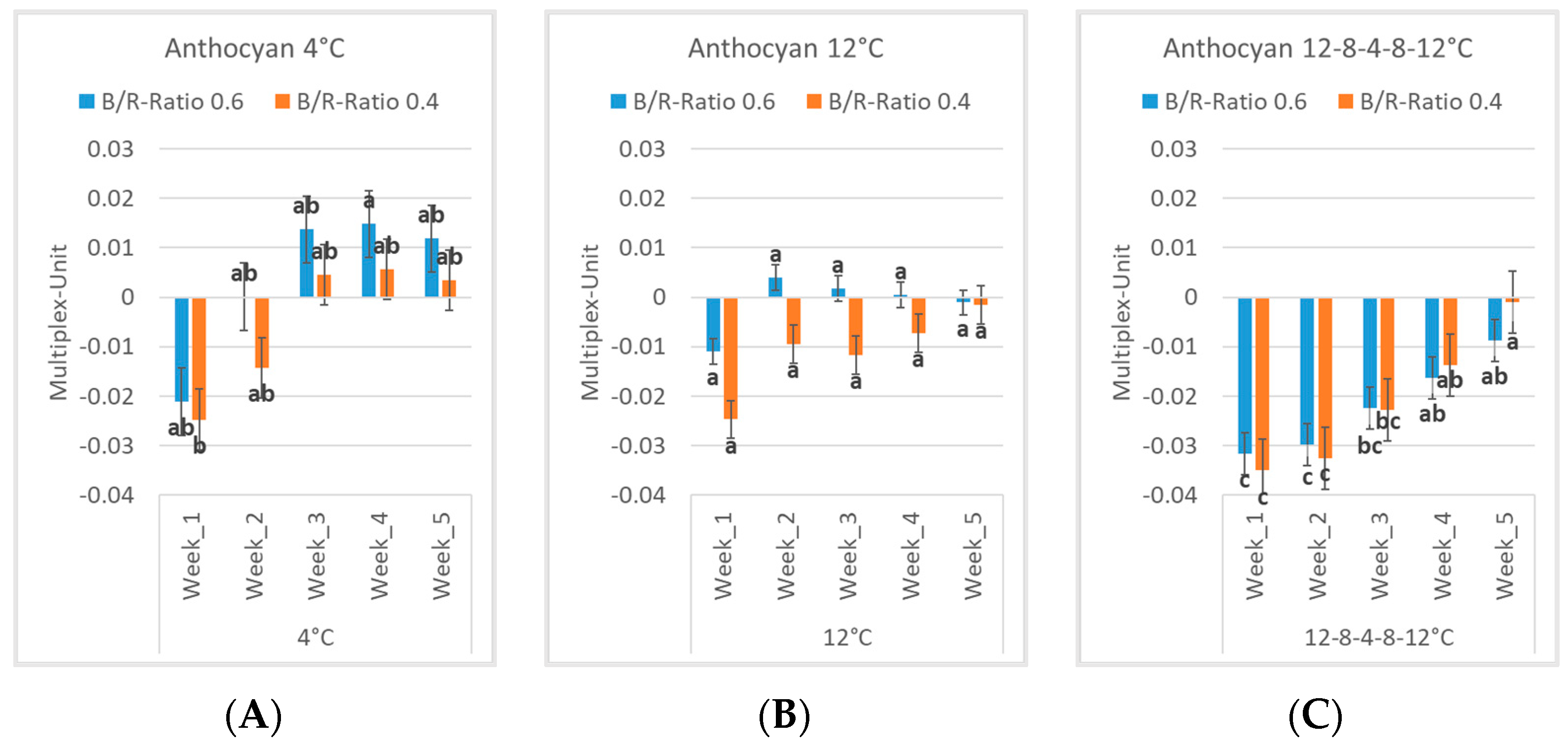
3.1. Internal Parameters: Flavonols and Anthocyanins
3.2. Differences among Experiments
3.3. Differences in Length Increase
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babla, M.; Cai, S.; Chen, G.; Tissue, D.T.; Cazzonelli, C.I.; Chen, Z.-H. Molecular Evolution and Interaction of Membrane Transport and Photoreception in plants. Front. Genet. 2019, 10, 956. [Google Scholar] [CrossRef]
- Saleem, M.; Atta, B.M.; Ali, Z.; Bilal, M. Laser-induced fluorescence spectroscopy for early disease detection in grapefruit plants. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2020, 19, 713–721. [Google Scholar] [CrossRef]
- Idris, A.; Linatoc, A.C.; Abu Bakar, M.F.; Takai, Z.I.; Audu, Y. Effect of Light Quality and Quantity on the Accumulation of Flavonoid in Plant Species. J. Sci. Technol. 2018, 10, 32–45. [Google Scholar] [CrossRef]
- Tan, Z.G.; Qian, Y.L. Light Intensity Affects Gibberellic Acid Content in Kentucky Bluegrass. HortScience 2003, 38, 113–116. [Google Scholar] [CrossRef]
- Tegg, R.S.; Lane, P.A. A comparison of the performance and growth of a range of turfgrass species under shade. Aust. J. Exp. Agric. 2004, 44, 353. [Google Scholar] [CrossRef]
- Cereti, C.F.; Ruggeri, R.; Rossini, F. Cool-Season Turfgrass Species and Cultivars: Response to Simulated Traffic in Central Italy. Ital. J. Agron. 2010, 5, 53. [Google Scholar] [CrossRef]
- Reyes, T.H.; Pompeiano, A.; Ranieri, A.; Volterrani, M.; Guglielminetti, L.; Scartazza, A. Photosynthetic performance of five cool-season turfgrasses under UV-B exposure. Plant Physiol. Biochem. 2020, 151, 181–187. [Google Scholar] [CrossRef]
- Moser, L.E.; Hoveland, C. Cool-Season Grass Overview. In Cool-Season Forage Grasses; Moser, L.E., Buxton, D.R., Casler, M.D., Eds.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America (Agronomy Monographs): Madison, WI, USA, 1996; pp. 1–14. [Google Scholar]
- Laboisse, S.; Combes, D.; Escobar-Gutierrez, A.; Hurlus, J.M. Spatial distribution of simulated turfgrass photosynthesis in football stadium pitch. In Proceedings of the 2018 6th International Symposium on Plant Growth Modeling, Simulation, Visualization and Applications (PMA), Hefei, China, 4–8 November 2018; pp. 111–114. [Google Scholar]
- Mann, R.L. Growing sports turf in shady environment. Acta Hortic. 2016, 83–90. [Google Scholar] [CrossRef]
- Gardner, D.S.; Goss, R.M. Management of Turfgrass in Shade. In Turfgrass: Biology, Use, and Management; Stier, J.C., Horgan, B.P., Bonos, S.A., Eds.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2013; pp. 219–247. [Google Scholar]
- Abélard, E.; Galbrun, C. The effects of artificial lighting on sports turf. Int. Turfgrass Soc. Res. J. 2022, 14, 1016–1021. [Google Scholar] [CrossRef]
- Bourget, C.M. An Introduction to Light-emitting Diode. HortScience 2008, 43, 1944–1946. [Google Scholar] [CrossRef]
- Cocetta, G.; Casciani, D.; Bulgari, R.; Musante, F.; Kołton, A.; Rossi, M.; Ferrante, A. Light use efficiency for vegetables production in protected and indoor environment. Eur. Phys. J. Plus 2017, 132, 43. [Google Scholar] [CrossRef]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Yang, X.; Montano, S.; Ren, Z. How Does Photoreceptor UVR8 Perceive a UV-B Signal? Photochem. Photobiol. 2015, 91, 993–1003. [Google Scholar] [CrossRef]
- Tilbrook, K.; Arongaus, A.B.; Binkert, M.; Heijde, M.; Yin, R.; Ulm, R. The UVR8 UV-B Photoreceptor: Perception, Signaling and Response. Arab. Book 2013, 11, e0164. [Google Scholar] [CrossRef]
- Su, J.; Liu, B.; Liao, J.; Yang, Z.; Lin, C.; Oka, Y. Coordination of Cryptochrome and Phytochrome Signals in the Regulation of Plant Light Response. Agronomy 2017, 7, 25. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R.; Nenadis, N.; Neugart, S.; Robson, M.; Agati, G.; Vepsäläinen, J.; Zipoli, G.; Nybakken, L.; Winkler, B.; Jansen, M.A.K. Assessing the response of plant flavonoids to UV radiation: An overview of appropriate technique. Phytochem. Rev. 2015, 14, 273–297. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants and developmental regulators: Relative significance in plants and human. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED System. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Agati, G.; Cerovic, Z.G.; Pinelli, P.; Tattini, M. Light-induced accumulation of ortho-dihydroxylated flavonoids as non-destructively monitored by chlorophyll fluorescence excitation technique. Environ. Exp. Bot. 2011, 73, 3–9. [Google Scholar] [CrossRef]
- Carlson, M.G.; Gaussoin, R.E.; Puntel, L.A. A review of precision management for golf course turfgrass. Crop Forage Turfgrass Manag. 2022, 8, e20183. [Google Scholar] [CrossRef]
- Fitz-Rodríguez, E.; Choi, C.Y. Monitoring turfgrass quality using multispectral radiometry. Trans. ASAE 2002, 45, 865. [Google Scholar] [CrossRef]
- Dong, L.; Xiong, L.; Sun, X.; Shah, S.; Guo, Z.; Zhao, X.; Liu, L.; Cheng, L.; Tian, Z.; Xie, F.; et al. Morphophysiological Responses of Two Cool-Season Turfgrasses with Different Shade Tolerance. Agronomy 2022, 12, 959. [Google Scholar] [CrossRef]
- Wang, T.; Chandra, A.; Jung, J.; Chang, A. UAV remote sensing based estimation of green cover during turfgrass establishment. Comput. Electron. Agric. 2022, 194, 106721. [Google Scholar] [CrossRef]
- Legris, M.; Nieto, C.; Sellaro, R.; Prat, S.; Casal, J.J. Perception and signalling of light and temperature cues in plant. Plant J. Cell Mol. Biol. 2017, 90, 683–697. [Google Scholar] [CrossRef]
- Balcerowicz, M. Phytochrome-interacting factors at the interface of light and temperature signalling. Physiol. Plant. 2020, 169, 347–356. [Google Scholar] [CrossRef]
- Sarkar, D.; Bhowmik, P.C.; Kwon, Y.I.; Shetty, K. Cold Acclimation Responses of Three Cool-season Turfgrasses and the Role of Proline-associated Pentose Phosphate Pathway. J. Amer. Soc. Hort. Sci. 2009, 134, 210–220. [Google Scholar] [CrossRef]
- Ben Ghozlen, N.; Cerovic, Z.G.; Germain, C.; Toutain, S.; Latouche, G. Non-destructive optical monitoring of grape maturation by proximal sensing. Sensors 2010, 10, 10040–10068. [Google Scholar] [CrossRef]
- Chen, D.-Q.; Li, Z.-Y.; Pan, R.-C.; Wang, X.-J. Anthocyanin Accumulation Mediated by Blue Light and Cytokinin in Arabidopsis Seedlings. J. Integr. Plant Biol. 2006, 48, 420–425. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. Int. J. Exp. Plant Biol. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Martel, A.B.; Strugnell, C.A. Environmental Factors Regulate Plant Secondary Metabolites. Plants 2023, 12, 447. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Peer, W.A.; Bandyopadhyay, A.; Blakeslee, J.J.; Makam, S.N.; Chen, R.J.; Masson, P.H.; Murphy, A.S. Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana. Plant Cell 2004, 16, 1898–1911. [Google Scholar] [CrossRef] [PubMed]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Xu, F.; He, S.; Zhang, J.; Mao, Z.; Wang, W.; Li, T.; Hua, J.; Du, S.; Xu, P.; Li, L.; et al. Photoactivated CRY1 and phyB Interact Directly with AUX/IAA Proteins to Inhibit Auxin Signaling in Arabidopsis. Mol. Plant 2018, 11, 523–541. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 4 °C | 12 °C | 12–8–4–8–12 °C | ||||
|---|---|---|---|---|---|---|
| Flavonol | Anthocyanin | Flavonol | Anthocyanin | Flavonol | Anthocyanin | |
| Week | 1.90 × 10−12 *** | 0.00 *** | 1.55 × 10−10 *** | 0.40 | 8.81 × 10−10 *** | 9.93 × 10−9 *** |
| Spectrum | 0.00 *** | 0.09 | 0.50 | 0.11 | 0.31 | 0.75 |
| Interaction Week/Spectrum | 0.14 | 0.98 | 0.78 | 0.94 | 0.21 | 0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schweiger, A.; Bernhardt, H. Influence of Temperature and LED Light Spectra on Flavonoid Contents in Poa pratensis. AgriEngineering 2024, 6, 2167-2178. https://doi.org/10.3390/agriengineering6030127
Schweiger A, Bernhardt H. Influence of Temperature and LED Light Spectra on Flavonoid Contents in Poa pratensis. AgriEngineering. 2024; 6(3):2167-2178. https://doi.org/10.3390/agriengineering6030127
Chicago/Turabian StyleSchweiger, Andreas, and Heinz Bernhardt. 2024. "Influence of Temperature and LED Light Spectra on Flavonoid Contents in Poa pratensis" AgriEngineering 6, no. 3: 2167-2178. https://doi.org/10.3390/agriengineering6030127
APA StyleSchweiger, A., & Bernhardt, H. (2024). Influence of Temperature and LED Light Spectra on Flavonoid Contents in Poa pratensis. AgriEngineering, 6(3), 2167-2178. https://doi.org/10.3390/agriengineering6030127