1. Introduction
In the Mediterranean basin, where dairy sheep and goats breeding is widespread, the use of agricultural by-products in the diet of small ruminants is an ancient practice. Today the great availability of agro-industrial by-products produced worldwide opens the door to other products to be tested. Noteworthy, the use of co-feeds can contribute to reducing the impact associated with ruminant’s management. Several positive aspects were observed when agro-industrial by-products (e.g., grape, olive, tomato, citrus pulp and myrtle residues) were included in the diet of small dairy ruminants. In particular, it has been reported that the diet implementation with these by-products has beneficial effects on ruminal metabolism, animal health, and quality of derived products [
1,
2,
3].
Cocoa husks represent the part of cocoa pod left over and are the principal by-products derived from
Theobroma cacao L., representing an important crop for many tropical developing countries. Cocoa husks are obtained after the removal of the cocoa beans, the principal commercial product of cocoa processing [
3]. Husks represent 70–75% of the fruit weight [
4] and, besides limited local use, are considered as waste [
5]. Cocoa pod husks along with other industrial by-products such as cocoa bean shells, cocoa bean meal and cocoa germ can be used as feed especially in the developing countries.
Purine alkaloids, including theobromine, theophylline and caffeine, and other methyl-xanthines, play a substantial role in pharmacology and food chemistry. A limited number of plant species accumulates purine alkaloids, such as caffeine and theobromine, which are synthesized from xanthosine, a catabolite of purine nucleotides. The most widely distributed methylxanthine in the plant kingdom is the antioxidant caffeine, which accumulates in leaves and seeds of tea (
Camellia sinensis), coffee (
Coffea arabica) and a limited number of other species. Considerable amounts of theobromine are stored in the seeds of cacao (
Theobroma cacao) [
6]. Moreover, from a pharmacokinetic point of view, theobromine can be easily adsorbed, with a low protein binding affinity and distributed into body tissues with no specific tissue accumulation [
7]. Despite the positive health benefits of consuming foods containing purine alkaloids [
8] one of the limits in the use of cocoa husks as supplementary feed for animals is the fact that in certain amount, theobromine, can be toxic [
9]. The EU Scientific Opinion of the Panel on Contaminants in the Food Chain reported that when exposed to theobromine dairy cows showed reduced milk yield, increase in fat levels, and adverse effects such as hyperexcitability, sweating, increase in respiration and heart rates [
10]. Reported levels of theobromine in cocoa husk meal are in the range of 1.5–4.0 g per kg of feed material and UE regulations set for ruminants the maximum levels of theobromine in feed materials at 300 mg/kg. On the other side, cocoa husks on a dry matter basis (DM) are a good source of neutral detergent fiber (NFC 38–44%) and high variable concentrations of non-fiber carbohydrate (NFC 17.5–47% on DM), crude proteins (CP 2.1–9.1% on DM), and lipids (ethyl esters 0.6–4.7% on DM); phenolic compounds account for 4.6–6.9% of gallic acid/100 g DM. However, the high content of acid detergent lignin (19.6% on DM) derived from cocoa pericarp affects the digestibility of this by-product, limiting its further use in animal feeding [
11].
Metabolomics is a largely used branch of omics sciences used to explore the composition, relative levels, dynamics, and interactions amongst metabolites in an organism or biological system in response to various stimuli, diet or treatments. In the field of dairy, metabolomics has been used for example in the authentication of beef production systems [
12], in the milk to investigate metabolite differences of healthy, subclinical, and clinical mastitis cows [
13] or sheep milk grazed on different grazing systems [
14], while in cheese samples used for discriminate cheese produced from raw or thermized milk [
15].
To our knowledge, experimental data on theobromine carry over in milk are not available, as well as the metabolic effects of cocoa husks diet supplementation. In a previous work, milk and blood parameters of ewes supplemented with different doses of cocoa husks were evaluated [
5]. In this experiment DMI and milk yield were not affected; however, milk protein content increased with cocoa husk supplementation. Furthermore, the cocoa husks supplementation lowered milk somatic cell count (SCC), suggesting a beneficial role of these by-products on mammary health status. Both systemic and local health conditions were good as evidenced by hematological parameters and electrophoretic profile of serum protein fractions [
5]. Both systemic and local health conditions were good as evidenced by hematological parameters and electrophoretic profile of serum protein fractions [
5]. Taking into consideration these results, we decided to investigate the potential beneficial nutritional outcomes by exploring, for the first time, milk metabolite profile changes of healthy ewes fed with a diet supplemented with 100 g/d per head soybean hulls that were replaced with 50 and 100 g/d per head of cocoa husks, respectively. In order to pursue these objectives, we used a liquid chromatography mass spectrometry metabolomics platform and multivariate statistical analysis.
2. Materials and Methods
The experiment was approved by the Ethics Committee of the University of Sassari (no. 54584/2018).
Animals and diets. As previously described [
5], twenty-four Sarda dairy ewes in mid lactation stage (120 days in milking), at third parity, with an average body weight of 42.5 kg were divided into three homogenous groups. Each experimental group received a total mixed ration (2.58 kg/d per head) as the basal diet and received (a) a supplement of 100 g/d per head of soybean hulls (CH0 group); (b) half of the soybean hulls supplement was replaced with 50 g/d per head of cocoa husks (CH50 group); and (c) soybean hulls was totally replaced with 100 g/d per head of cocoa husks (CH100 group). Theobromine concentration was 130 ± 12 and 253 ± 27 mg/kg of DM for the CH50 and CH100 diets, respectively. The CH was administered individually during milking, mixed with beet pulp to increase palatability. The experiment lasted 8 weeks (from May to June 2018), with the last 4 weeks of sampling collection. Individual milk samples were collected weekly at morning milking.
Chemicals. Sigma Aldrich (Milan, Italy) reagents were used for the analysis. Bi-distilled water was obtained with a MilliQ purification system (Millipore, Milan, Italy).
Sample preparation for UHPLC-QTOF/MS analysis. Individual milk samples from the morning milking were stored into sterile plastic Falcon tubes at −80 °C before analysis. Prior to UHPLC-QTOF/MS analysis, the 107 individual sheep milk samples were thawed and thoroughly vortex mixed. A total of 150 µL of milk samples were transferred to Eppendorf tube containing 10 uL of the internal mixture of standards (Splash, Lipidomics, Sigma Aldrich, Milan, Italy) and added with 525 µL of methanol and 525 µL of MTBE. Samples were then mixed by vortexing for 1 min. Next, samples were centrifuged at 4000 rpm for 15 min. Subsequently, a second round of centrifuge was performed at 12000 rpm for 5 min. Before transferring to autosampler vials, the supernatant was filtered through a 0.22 µm MS nylon syringe filter.
UHPLC-QTOF/MS analysis. After extraction with MTBE/methanol, the supernatant of milk samples was analyzed with a 6560 Q-TOF (Agilent Technologies, Palo Alto, CA, USA) coupled with an Agilent 1290 Infinity II LC system. An aliquot of 1.0 μL from each sample was injected in a Kinetex 5 µm EVO C18 100 A, 150 mm × 2.1 μm column (Agilent Technologies, Palo Alto, CA, USA). The column was maintained at 50 °C at a flow rate of 0.5 mL/min. The mobile phases consisted of (A) methanol:water (90:10, v/v) with ammonium acetate (10 mM) and (B) acetonitrile:methanol:2-propanol (20:30:50, v/v) with ammonium acetate (10 mM). The chromatographic separation was obtained using the following gradient—0 min 70% B kept for 1 min; 1–3.5 min 86% B; 3.5–10 min 86% B; 10.1–17 min 100% B; 17.1–19 min 70% B. The analytical setup used was equipped with an Agilent jet stream technology source which was operated in both positive and negative ion modes with the following parameters: gas temperature,200 °C; gas flow (nitrogen) 10 L/min; nebulizer gas (nitrogen), 50 psig; sheath gas temperature, 300 °C; sheath gas flow, 12 L/min; capillary voltage 3500 V for positive and 3000 V for negative; nozzle voltage 0 V; fragmentor 150 V; skimmer 65 V, octapole RF 7550 V; mass range, 40–1700 m/z; capillary voltage, 3.5 kV; collision energy 20 eV in positive and 25 eV in negative mode. An Agilent MassHunter software was used for instrument control (revision B.09.00).
Multivariate statistical data analysis (MVA). The QTOF-MS data were submitted to the web platform XCMS [
16]. The workflow allows feature detection, retention time correction, alignment, annotation, statistical analysis, and data visualization. Retention time,
m/z values, and intensities of each feature obtained from XCMS were then submitted to multivariate statistical analysis (MVA), as implemented in SIMCA-P+ software (version 14.1, Umetrics, Umeå, Sweden). Prior to MVA, QTOF-MS features were mean centered and scaled to unit variance column-wise. Principal component analysis (PCA) was performed to investigate sample distributions, deviating features and prevailing trends. This technique reduced the dimensionality of data set retaining most of the variance. Results are shown as scatter plots of scores and loadings in the first principal components, reporting sample and variable displacement in the hyperplane, respectively. The partial least squares-discriminant analysis (PLS-DA) and its orthogonal variant (OPLS-DA) were performed for classification of samples and identification of the most discriminant variables. The quality of the models was evaluated based on the cumulative parameters R
2Y and Q
2Y, the latter estimated by cross validation, and tested for overfitting using a y-table permutation test (
n = 400). The variable importance in projection (VIP) score summarizes the contribution of each variable to the model. The VIP scores in the predictive component were analyzed and only those metabolites having VIP values > 1 were considered as discriminant between the classes [
17].
3. Results and Discussion
This untargeted metabolomics approach on dairy ewes was conceived to provide a molecular and biological insights on milk metabolite changes induced by a diet integrated with 50 or 100 g/d of cacao husks. Moreover, we designed this cost-efficient metabolomics study to measure predictive metabolite biomarkers to be used in exploring feed suitability and efficiency.
For these purposes, a total of 107 sheep milk samples were analyzed with a UHPLC-QTOF/MS platform, (
Figure S1 reported the LC-MS chromatograms). After analysis, raw mass spectrometry data acquired were then uploaded to the XCMS platform, which generated an average of 7700 and 3300 features for the positive and negative modes, respectively. The XCMS outputs were submitted to multivariate statistical analysis using the software SIMCA.
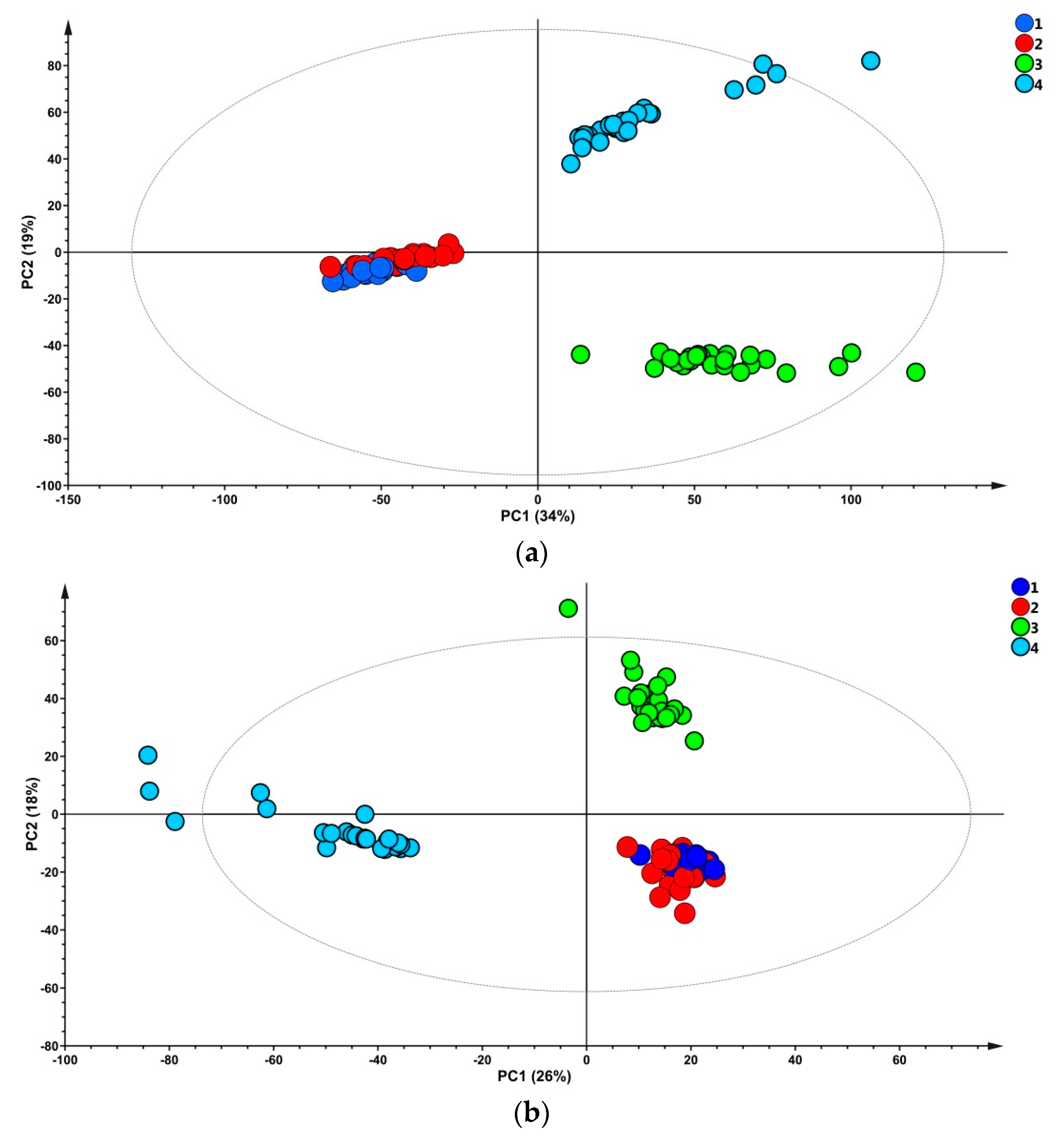
A PCA was performed to explore similarities of milk sample metabolic profiles of the three diets over the experimental sampling period. Results indicated that metabolic similarities of milk samples were mainly due to the time of sampling, as clearly visible in the score plots shown in
Figure 1 for the positive and negative ionization modes.
Information collected for the two ionization modes yielded similar results (i.e., samples collected in the two first weeks of treatment clustered together as a result of similar metabolite profile, while milk samples from the third and the last week of experiment can be easily differentiated along the second principal component (PC2) in the positive and negative ionization modes). Interestingly, the same pattern was observed for milk yield, as shown in
Figure S2, whereby the first two weeks gave comparable average values for all the group samples, followed by a drastic milk yield drop after the second week of sampling (from the 13 to 26 of June). A decrease of milk yield and lactose content and an increase in fat and protein content, due to a milk concentration effect, during the late-spring/summer seasons were already reported as a natural evolution of the lactation curve in dairy Sarda sheep [
18]. Probably these milk qualitative and quantitative changes due to lactation seasonality have caused an overall modification of the metabolic profiles captured by the PCA.
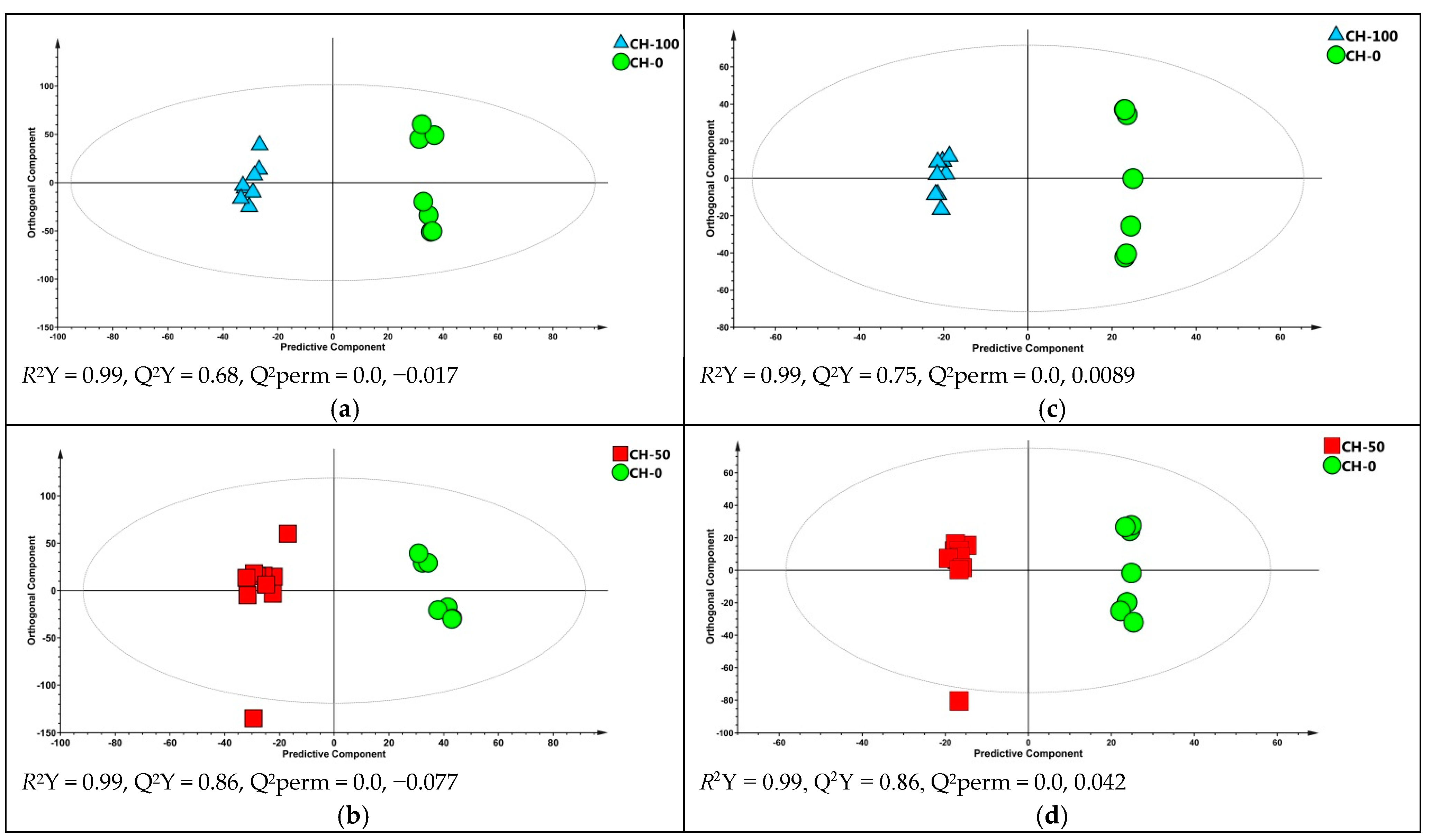
Attempts to differentiate samples based on the diets (CH0, CH50, and CH100) were carried out by performing three-classes PLS-DA for each time point for positive and negative modes. The multivariate statistical analysis failed to discriminate samples for the first three weeks of sampling (data not shown) whereas at the fourth week of treatment, samples were properly classified, with high statistical confidence (
Figure S3), indicating that the effects of the cocoa husks supplementation on milk metabolic profile were evident only in the last time point. Subsequently, with the purpose of finding discriminant metabolites, pairwise OPLS-DA were carried out comparing the three groups of milk samples collected in the last week of treatment (
Figure 2). In
Table 1 and
Table 2 we reported the list of discriminant metabolites, their biological significance, and regulation in sheep milk samples.
From the analysis of discriminant metabolites, as summarized in
Table 1 and
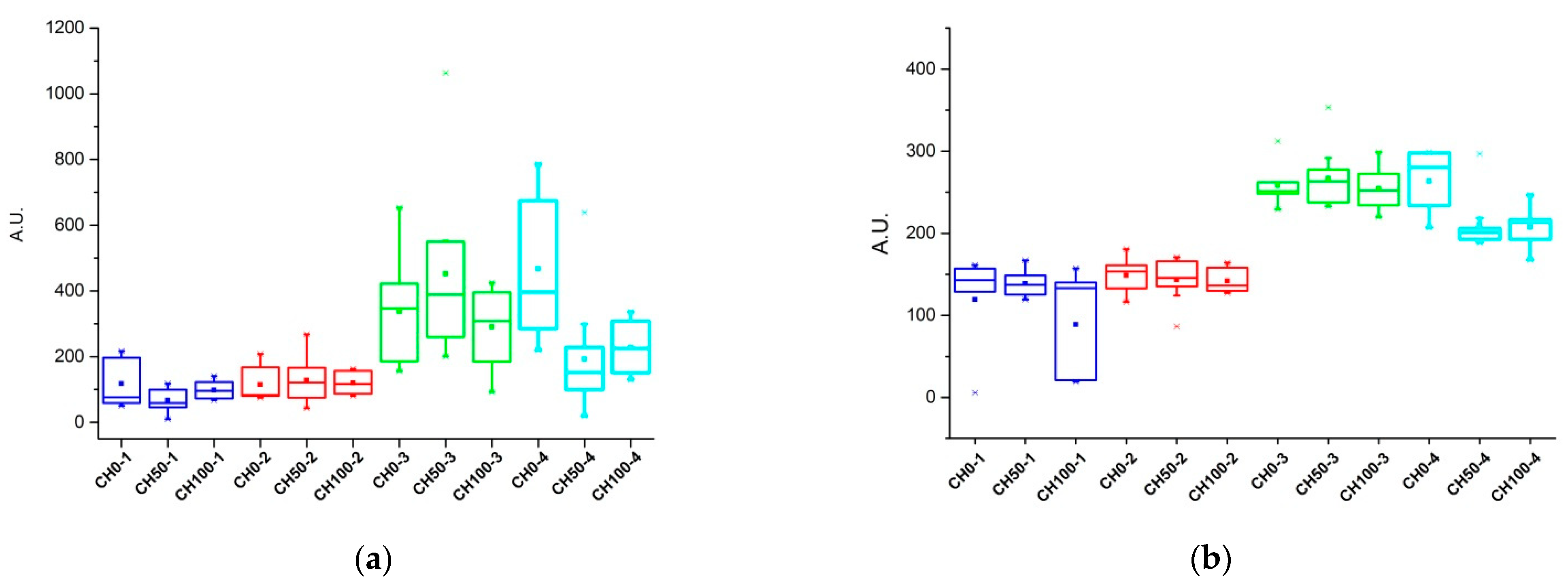
Table 2, we found that the addition of cocoa husks in the dairy lactating sheep diet affected levels of different metabolites which are involved in different metabolic pathways, including ubiquinol synthesis and thyroid hormone metabolism. In particular, we found that in milk samples of lactating ewes the thyroid hormone L-thyroxine (T4) and tetraiodothyroacetate, or tetrac (T4A), were down regulated for both CH50 and CH100 groups, thus suggesting a modification of the thyroid hormone metabolism. Levels of T4 and T4A are shown in
Figure 3, indicating that, in the first two weeks of treatment, group samples showed similar average values. Levels where higher in the third week and in the last week CH50 and CH100 groups showed lower values when compared to CH0.
The thyroid hormones T4 and 3,5,3′-triiodo-L-thyronine (T3) are regulated at many metabolic levels. In addition to deiodination (thyroid hormone metabolism I pathway), other routes of thyroid hormone metabolism have been described and metabolically active metabolites have been identified. In the thyroid hormone metabolism pathway (via degradation), small amounts of T4 may also be oxidatively deaminated at the alanine side chain to yield T4A. The oxidative deamination due the kidney L-amino acid oxidase seems the most probable explanation [
19,
20]. The acetic acid analogues of thyroid hormones are formed in the liver and other peripheral tissue by a two-stage process involving transamination followed by oxidative decarboxylation [
21]. T4A may be conjugated to form ether glucuronides or ester glucuronides of these metabolites may form by conjugation of the phenolic hydroxyl group, or carboxyl group respectively [
20].
Thyroid hormones in milk should derive from circulating blood, then undergoing both desiodative and non-desiodative metabolism. A reduction in milk T4 could; therefore, be derived from a reduction in circulating T4 levels or from the reduction in T4 level available at the mammary gland, due to the effect of interferents on the transporters of thyroid hormones, or, finally, by increased desiodation of T4. The reduction of milk T4A level seems the logical direct consequence of the reduction of T4, deriving the T4A directly from T4. Thyroid hormones play a relatively important role in lactational processes. In small ruminants and, in particular, in free grazing animals which are subjected to seasonality, endogenous (breed, age, gender, physiological state) and environmental factors (climate, season, with a primary role of nutrition) are able to impact on thyroid activity and hormone blood levels [
22]. During heat stress, an increase of blood T4 concentration and a decrease of milk production were reported in sheep [
23,
24]. This trend was evident also in our milk samples, which were collected in June, where, during the experimental period, in the CH0 group the milk T4 levels inversely follow the milk production trend (
Figures S2 and S4). Differently from CH0, in the last week of sampling, milk T4 levels dropped in the CH50 and CH100 groups (
Figure S4), thus indicating a goitrogenic effect of cocoa husks on milk T4. In the rat, Wolff and Varrone (1969) reported that methyl-xanthines (i.e., theobromine, theophylline, and caffeine) showed a goitrogenic effect and are; therefore, to be considered weak anti-thyroid agents if present in the diet at concentrations varying between 5–20 microM [
25]. Further studies will be carried out to better evaluate this effect in sheep, measuring the volume of the thyroid glands and the concentration of T4 and TSH in ewes’ blood after consumption of cocoa husks. At the milk level, which only contains traces of iodothyronines, which; therefore, do not represent a significant nutrient of the milk itself, as opposed to iodine, any reduction in concentration should not be particularly relevant for its nutritional value.
Notably, phenylalanine was found up-regulated in CH50 and CH100 milk samples. Considering that phenylalanine is the precursor of tyrosine and the thyroid hormone T4 is a tetraiodinated derivative of tyrosine [
22], this fact confirms a dysregulation of thyroid hormones metabolism.
The discriminant metabolites: 2-methoxy-6-(all-trans-octaprenyl)phenol, 6-methoxy-3-methyl-2-all-trans-decaprenyl-1,4-benzoquinol, and 3,4-dihydroxy-5-all-trans-decaprenylbenzoate, involved in the ubiquinol-10 biosynthesis, were found down regulated in milk samples of the CH50 and CH100 groups with respect to CH0 (
Table 1). Coenzyme Q (CoQ10), also known as ubiquinone, is an essential component of the mitochondrial respiratory chain and participates in aerobic cell respiration. The reduced ubiquinol form of CoQ10 is a chain-breaking antioxidant, decreasing oxidative damage caused by lipid peroxidation within mitochondria. The cocoa husks feeding regime, showed markedly lower levels of intermediate metabolites of the CoQ10 pathway compared to the controls. To the best of our knowledge, no studies of theobromine effects on CoQ10 pathway have been widely reported, but interference of phenolic compounds has been found in liver of rats [
26]. Specifically, the authors found that pentagalloylglucose, which is a component of tannic acid, was able to interfere with the CoQ10 pathway: At low concentration inhibiting the electron transport system, while at high concentration impairing the structural integrity of the mitochondrial membrane. Consistently, CoQ10 was found to affect the pharmacokinetic parameters of theophylline [
27], an alkaloid of the xanthine family.
Remarkably, in the negative ionization mode we found different discriminant metabolites involved in the purine metabolism (
Table 1). Inosine is a purine nucleoside formed by deamination of adenosine and, in the context of purine metabolism, is further transformed to hypoxanthine by the enzyme phosphate alpha-D-ribosyltransferase and then to xanthine and to uric acid. The fact that inosine was found upregulated in milk samples of sheep consuming cocoa husks may be associated to the inhibition of the enzyme phosphate alpha-D-ribosyltransferase, in order to reduce the increase of xanthines and uric acid derived from the metabolism of theobromine.
Our experimental findings are in accordance with scientific opinion of EFSA [
10] on theobromine as an undesired substance in animal feed. In the EFSA report, theobromine was reported as showing moderate acute toxicity on dog and rodents, while in horses, which are particularly susceptible to theobromine, the liver and thyroid were affected, and pigs showed growth retardation, diarrhea and lethargy. Particular attention should be paid on the levels of theobromine and other alkaloids present in the industrial by-products in the preparing of the dairy sheep’s diet.

 ,
, 






{kind=link}
{kind=link}
{kind=link}