
Microsatellite Loci Reveal Heterozygosis and Population Structure in the Critically Endangered Southern River Terrapin (Batagur affinis ssp.) of Peninsular Malaysia †



Abstract
:1. Introduction
2. Materials and Methods
2.1. Populations Description
2.2. DNA Extraction and Microsatellite Analysis
2.3. Statistical Analysis
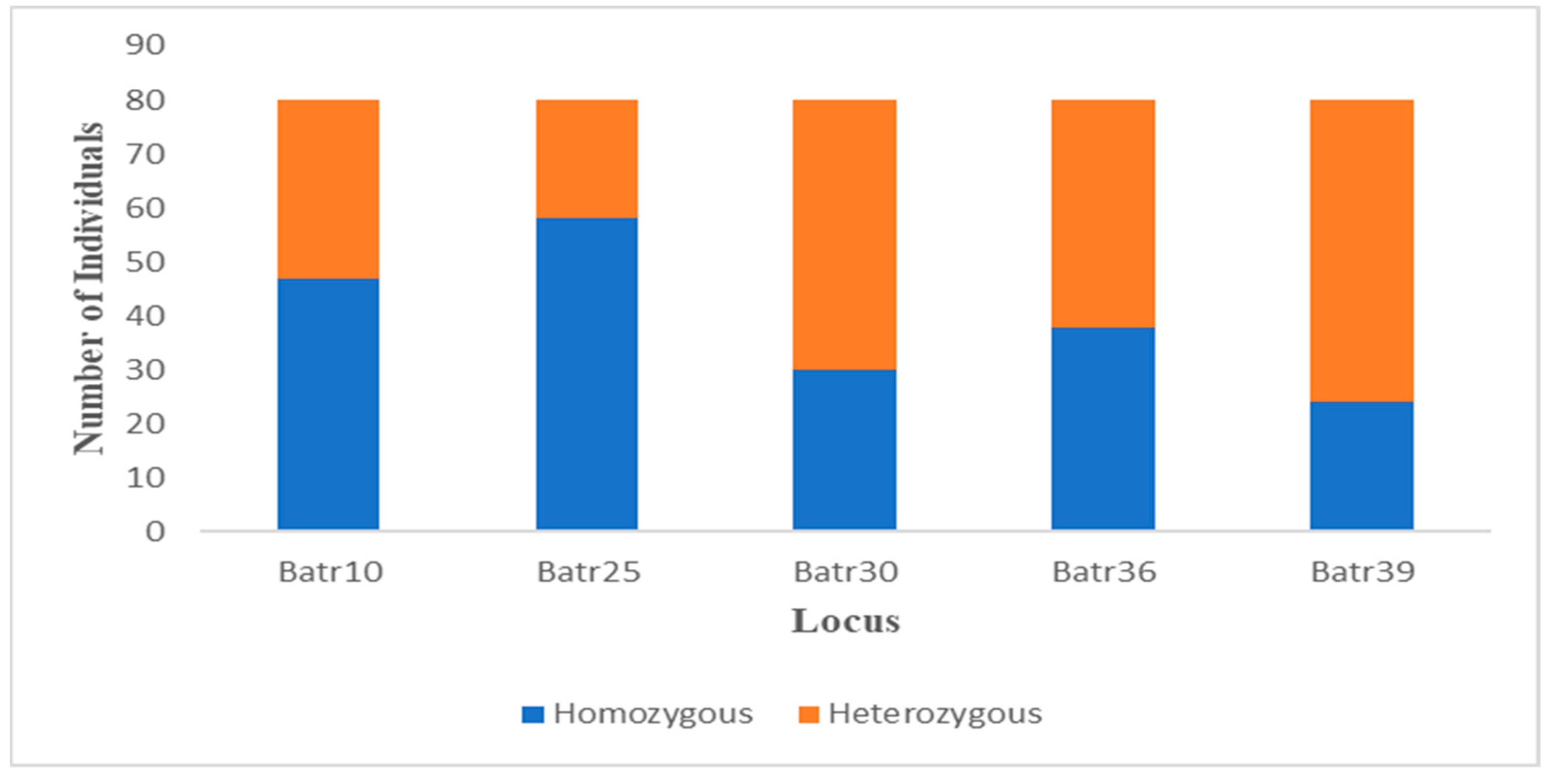
2.3.1. Genetic Diversity
2.3.2. Population Structure
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moll, E.; Platt, S.; Chan, E.H.; Horne, B.; Platt, K.; Praschag, P.; Chen, P.N.; Van Dijk, P.P. Batagur affinis (Cantor 1847)—Southern River Terrapin, Tuntong. Chelonian Res. Monogr. 2015, 5, 90–91. [Google Scholar] [CrossRef] [Green Version]
- Batagur affinis. The IUCN Red List of Threatened Species. Available online: https://doi.org/10.2305/IUCN.UK.2016-1.RLTS.T170501A1315041.en (accessed on 1 November 2021).
- Praschag, P.; Holloway, R.; Georges, A.; Päckert, M.; Hundsdörfer, A.K.; Fritz, U. A new subspecies of Batagur affinis (Cantor, 1847), one of the world’s most critically endangered chelonians (Testudines: Geoemydidae). Zootaxa 2009, 2233, 57–68. [Google Scholar] [CrossRef]
- Salleh, M.H.M.; Esa, Y. The mtDNA D-loop Marker Identifies the Genetic Variability of Indochina’s Batagur affinis. In 1st Postgraduate Seminar on Agriculture and Forestry 2021 (PSAF 2021); UPM Press: Serdang, Selangor, Malaysia, 2021; p. 82. [Google Scholar]
- Mistar Siregar, A.J.; Singleton, I. Presence and Distribution of the Southern River Terrapin Batagur Affinis and Painted Terrapin Batagur Borneoensis in Eastern Coast of Sumatra, Auckland, New Zealand; 25p, Unpublished Report to Auckland Zoo.
- Abdul Muneer, P.M. Molecular Genetic Characterisation of Endemic Yellow Catfish, Horabagrus brachysoma (Gunther). Ph.D. Thesis, Cochin University of Science and Technology, Cochin, India, 2005. [Google Scholar]
- Love, C.N.; Hagen, C.; Horne, B.D.; Jones, K.L.; Lance, S.L. Development and characterization of thirty novel microsatellite markers for the critically endangered Myanmar Roofed Turtle, Batagur trivittata, and cross-amplification in the Painted River Terrapin, B. borneoensis, and the Southern River Terrapin, B. affinis, using paired-end Illumina shotgun sequencing. Conserv. Genet. Resour. 2013, 5, 383–387. [Google Scholar] [CrossRef]
- Chatterji, S.; Pachter, L. Reference based annotation with GeneMapper. Genome Biol. 2006, 7, R29. [Google Scholar] [CrossRef] [Green Version]
- Glaubitz, J.C. convert: A user-friendly program to reformat diploid genotypic data for commonly used population genetic software packages. Mol. Ecol. Notes 2004, 4, 309–310. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT (Version 1.2): A Computer Program to Calculate F-Statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Dupanloup, I.; Schneider, S.; Excoffier, L. A simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef]
- Crawford, M.H. Genetic structure of circumpolar populations: A synthesis. Am. J. Hum. Biol. 2007, 19, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Magri, D.; Vendramin, G.G.; Comps, B.; Dupanloup, I.; Geburek, T.; Gömöry, D.; Latałowa, M.; Litt, T.; Paule, L.; Roure, J.M.; et al. A new scenario for the Quaternary history of European beech populations: Palaeobotanical evidence and genetic consequences. New Phytol. 2006, 171, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Neff, B.D.; Gross, M.R. Microsatellite evolution in vertebrates: Inference from ac dinucleotide repeats. Evolution 2001, 55, 1717–1733. [Google Scholar] [CrossRef]
- Esa, Y.B.; Siraj, S.S.; Rahim, K.A.A.; Daud, S.K.; Ho, G.H.; Tan, S.G.; Syukri, M.F. Genetic Characterisation of Two Mahseer Species (Tor douronensis and Tor tambroides) Using Microsatellite Markers from Other Cyprinids. Sains Malays. 2011, 40, 1087–1095. [Google Scholar]
- Xu, Q.; Liu, R. Development and Characterization of Microsatellite Markers for Genetic Analysis of the Swimming Crab, Portunus trituberculatus. Biochem. Genet. 2010, 49, 202–212. [Google Scholar] [CrossRef]
- Spitzweg, C.; Praschag, P.; DiRuzzo, S.; Fritz, U. Conservation genetics of the northern river terrapin (Batagur baska) breeding project using a microsatellite marker system. Salamandra 2018, 54, 63–70. [Google Scholar]
- Callen, D.F.; Thompson, A.D.; Shen, Y.; Phillips, H.A.; Richards, R.I.; Mulley, J.C.; Sutherland, G.R. Incidence and origin of “null” alleles in the (AC)n microsatellite markers. Am. J. Hum. Genet. 1993, 52, 922–927. [Google Scholar]
- Chen, P.N. Conservation of the Southern River Terrapin Batagur affinis (Reptilia: Testudines: Geoemydidae) in Malaysia: A case study involving local community participation. J. Threat. Taxa 2017, 9, 10035. [Google Scholar] [CrossRef]
- Hale, M.L.; Burg, T.M.; Steeves, T.E. Sampling for Microsatellite-Based Population Genetic Studies: 25 to 30 Individuals per Population Is Enough to Accurately Estimate Allele Frequencies. PLoS ONE 2012, 7, e45170. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Samani, N.K.; Esa, Y.; Amin, S.N.; Ikhsan, N.F.M. Phylogenetics and population genetics of Plotosus canius (Siluriformes: Plotosidae) from Malaysian coastal waters. PeerJ 2016, 4, e1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, C.J.; Bin Esa, Y.; Ismail, S.; Kamarudin, M.S. Population Structure of the Blue Swimmer Crab Portunus pelagicus in Coastal Areas of Malaysia Inferred from Microsatellites. Zool. Stud. 2017, 56, e26. [Google Scholar] [CrossRef]
- Pedall, I.; Fritz, U.; Stuckas, H.; Valdeón, A.; Wink, M. Gene flow across secondary contact zones of the Emys orbicularis complex in the Western Mediterranean and evidence for extinction and re-introduction of pond turtles on Corsica and Sardinia (Testudines: Emydidae). J. Zool. Syst. Evol. Res. 2010, 49, 44–57. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Huchard, E.; Alvergne, A.; Féjan, D.; Knapp, L.A.; Cowlishaw, G.; Raymond, M. More than friends? Behavioural and genetic aspects of heterosexual associations in wild chacma baboons. Behav. Ecol. Sociobiol. 2010, 64, 769–781. [Google Scholar] [CrossRef]
- Smith, E.M.; Hoffman, J.I.; Green, L.E.; Amos, W. Preliminary association of microsatellite heterozygosity with footrot in domestic sheep. Livest. Sci. 2012, 143, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Romero-Nava, C.; León-Paniagua, L.; Ortega, J. Microsatellites loci reveal heterozygosis and population structure in vampire bats (Desmodus rotundus) (Chiroptera: Phyllostomidae) of Mexico. Revista Biol. Trop. 2014, 62, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Das, D.; Singh, S.K.; Bierstedt, J.; Erickson, A.; Galli, G.L.; Crossley, D.A.; Rhen, T. Draft Genome of the Common Snapping Turtle, Chelydra serpentina, a Model for Phenotypic Plasticity in Reptiles. G3 Genes Genomes Genet. 2020, 10, 4299–4314. [Google Scholar] [CrossRef]
- Stoeckel, S.; Grange, J.; Fernández-Manjarres, J.F.; Bilger, I.; Frascaria-Lacoste, N.; Mariette, S. Heterozygote excess in a self-incompatible and partially clonal forest tree species—Prunus aviumL. Mol. Ecol. 2006, 15, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Medellín, R.A.; Equihua, M.; Amin, M.A. Bat diversity and abundance as indicators of disturbance in Neotropical rain forests. Conserv. Biol. 2000, 14, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Balloux, F.; Amos, W.; Coulson, T. Does heterozygosity estimate inbreeding in real populations? Mol. Ecol. 2004, 13, 3021–3031. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Locality | Statistical | ||||||
|---|---|---|---|---|---|---|---|
| Ho | He | Allele Range | p Value | Rs | Hs | N | |
| KE | 0.66 | 0.85 | 7–17 | 0.02 | 11.4 | 0.86 | 20 |
| KB | 0.49 | 0.87 | 9–16 | 0 | 13.6 | 0.88 | 20 |
| BP | 0.51 | 0.8 | 8–11 | 0 | 9 | 0.81 | 20 |
| BK | 0.37 | 0.81 | 7–13 | 0.01 | 10 | 0.81 | 20 |
| Region | Kemaman | Kuala Berang | Bukit Pinang | Bota Kanan |
|---|---|---|---|---|
| Kemaman | - | 0.962 | 0.894 | 0.823 |
| Kuala Berang | 0.080 * | - | 0.749 | 0.574 |
| Bukit Pinang | 0.096 * | 0.083 * | - | 1.011 |
| Bota Kanan | 0.088 * | 0.064 * | 0.122 * | - |
| Source of Variation | df | Sum of Squares | Variance Components | Percentage of Variance | Fixation Indices |
|---|---|---|---|---|---|
| AMOVA. | |||||
| Among populations within groups | 3 | 30.869 | 0.18346 | 8.00 | FIS = 0.39871 |
| Among individuals within populations | 76 | 224.3 | 0.84128 | 36.68 | FST = 0.07999 |
| Among individuals within individuals | 80 | 101.5 | 1.26875 | 55.32 | FIT = 0.44680 |
| SAMOVA, 3 groups | |||||
| Among groups | 2 | 22.994 | 0.07244 | 3.14 | FIS = 0.39871 |
| Among populations within groups | 1 | 7.875 | 0.12309 | 5.34 | FSC = 0.05512 |
| Among individuals within populations | 76 | 224.3 | 0.84128 | 36.49 | FCT = 0.03142 |
| Among individuals within individuals | 80 | 101.5 | 1.26875 | 55.03 | FIT = 0.44970 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salleh, M.H.M.; Esa, Y. Microsatellite Loci Reveal Heterozygosis and Population Structure in the Critically Endangered Southern River Terrapin (Batagur affinis ssp.) of Peninsular Malaysia. Chem. Proc. 2022, 10, 11. https://doi.org/10.3390/IOCAG2022-12230
Salleh MHM, Esa Y. Microsatellite Loci Reveal Heterozygosis and Population Structure in the Critically Endangered Southern River Terrapin (Batagur affinis ssp.) of Peninsular Malaysia. Chemistry Proceedings. 2022; 10(1):11. https://doi.org/10.3390/IOCAG2022-12230
Chicago/Turabian StyleSalleh, Mohd Hairul Mohd, and Yuzine Esa. 2022. "Microsatellite Loci Reveal Heterozygosis and Population Structure in the Critically Endangered Southern River Terrapin (Batagur affinis ssp.) of Peninsular Malaysia" Chemistry Proceedings 10, no. 1: 11. https://doi.org/10.3390/IOCAG2022-12230
APA StyleSalleh, M. H. M., & Esa, Y. (2022). Microsatellite Loci Reveal Heterozygosis and Population Structure in the Critically Endangered Southern River Terrapin (Batagur affinis ssp.) of Peninsular Malaysia. Chemistry Proceedings, 10(1), 11. https://doi.org/10.3390/IOCAG2022-12230