1. Introduction
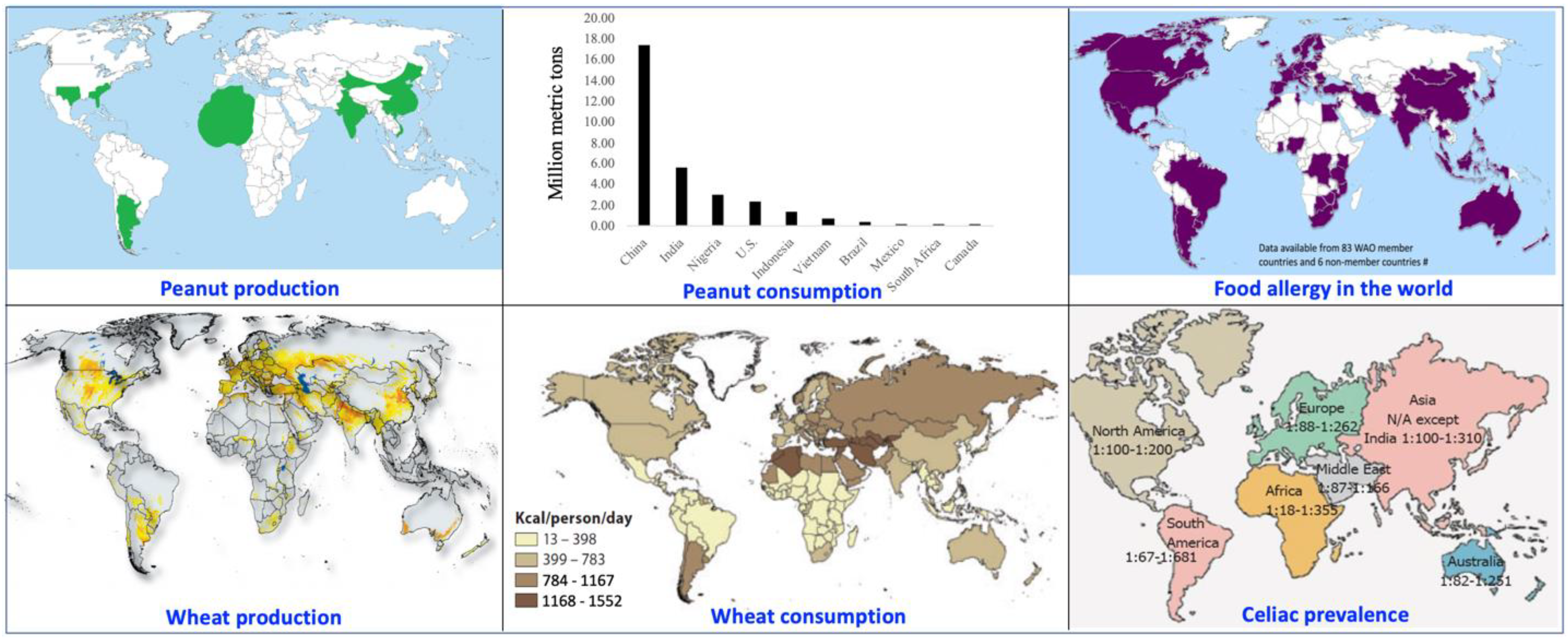
Sensitivity to wheat and peanut proteins is more common than initially thought and is not restricted to geographical areas where wheat and peanuts are produced (
Figure 1). It reflects on the widespread consumption and importance of these crops as a source of energy and proteins. Specifically, wheat is grown on over 15.5% of the world’s arable land [
1], provides dietary calories to 35% of the population, and feeds one-quarter of the global annual demand for dietary plant proteins [
2]. Similarly, the peanut is cultivated on over 3% of the world’s arable land [
3], and its oil (50% by weight) and protein (25% by weight)-rich kernels meet the protein needs of many worldwide, more specifically, the poor, earning it the reputation of the ‘poor man’s nut’ [
3,
4,
5,
6]. These two crops are vital for low- and middle-income countries and countries with large vegetarian and vegan populations (375 million individuals globally). Moreover, these crops are a significant component of the export profiles of industrialized countries.
Unfortunately, an uptick in the number of individuals diagnosed with celiac disease, a wheat-associated autoimmune disorder—and peanut allergy has been observed in the last two decades. According to a recent estimate, ~1.4% of the world population is diagnosed with celiac disease and ~2% of the Western population is diagnosed with peanut allergy [
7,
8]. Tragically, there is no therapy available for these sensitivities other than an abstinent diet, with a need to maintain an immunogen-free environment to prevent sensitive individuals’ accidental exposure to the antigen [
5,
9]. Following a gluten-free or peanut-free diet is a formidable task, due to the ubiquitous presence of wheat and peanuts in processed food, cosmetics, and even for medicines, social, and economic reasons [
5,
10]. Thus, this makes it essential to stay vigilant at all times, specifically in the case of children who are disproportionately affected by peanut allergies. However, an individual can develop sensitives to wheat and peanut later in life for several reasons, including changes in lifestyle that lead to alterations in the gut microbiome. Moreover, maintaining a gluten-free or peanut-free environment accommodating all individuals irrespective of their dietary restrictions is economically taxing.
Sensitivity to wheat can be classified into three kinds: (i) Celiac disease—a chronic disorder triggered by wheat gluten and allied proteins from related cereals such as barley and rye, which could develop into an autoimmune disorder if it remains untreated. (ii) Gluten allergy—an instant reaction to gluten exposure. iii) Non-celiac wheat sensitivity—a less-well-understood reaction to wheat and wheat products [
10,
11]. However, exposure to the peanut causes an instant allergic reaction in genetically predisposed individuals, which could vary in the severity of the response in different individuals [
5]. Primarily in peanut and wheat sensitivities, the trigger for these reactions is the indigestible proteins. A large family of proteins (~100 members), known as gluten in wheat [
9,
12], and about 16 seed-storage proteins in peanuts were recognized to trigger immune reactions in humans [
4]. Among wheat-gluten proteins, gliadins (α/β, γ, and ω), and peanut seed-storage proteins, Ara h1, Ara h2, Ara h3, and Ara h6 were identified as major immunogens [
5,
9].
2. Peanut Allergy
As mentioned earlier, peanuts are among the nine primary sources of food allergy recognized by the US Food and Drug Administration [
13], which, along with tree nuts, are responsible for 90% of allergy-related fatalities [
5]. Epidemiological studies show peanut allergy affects about 6.1 million children and adults in the US [
14]. In the last two decades, peanut allergy diagnoses have risen from 0.4% (1997) to 1.8% (2021) of the population, and the number is escalating [
8]. Raw peanuts and derived products lead to various allergic responses, the most common being immediate-onset anaphylaxis. Peanut allergy currently requires the complete exclusion of peanuts from the diet [
5]. Avoiding peanuts, other cross-reacting tree nuts, and legumes limits the options to obtain proteins. Moreover, avoiding the peanut is not straightforward due to its ubiquitous presence in processed foods, unintended contamination, and financial and social constraints. Sixteen proteins in peanut grains, namely Ara h1 to Ara h17 (Ara h4 is now characterized as an isoform of Ara h3 and renamed Ara h3.02), were recognized as allergens. Four proteins, Ara h1, Ara h2, Ara h3, and Ara h6, were identified as major allergens, as sera from 50% of patients respond to these allergens [
5].
Several efforts are underway to develop dietary alternatives, i.e., reduced immunogenicity peanuts and therapies for peanut-sensitive individuals [
5,
15]. These efforts vary significantly in the pursued approaches, such as physical (thermal processing, high-pressure processing, irradiation, electrical discharge, and ultrasonication) and chemical (chemical modification and enzyme processing) treatments of peanut kernels [
5] and genetic strategies to develop reduced-immunogenicity peanut genotypes [
16,
17,
18]. In parallel, efforts are underway to develop therapies for peanut allergy, in addition to a recently released vaccine (Palforzia
TM, Aimmune Therapeutics, Brisbane, Australia [
15]).
We undertook the genetics approach to address this problem from a prevention standpoint, unlike many previous attempts that focused on processing methods or therapeutics via developing reduced-immunogenicity peanut genotypes following a molecular-breeding approach. These genotypes would serve several purposes: (1) They could reduce the risk of an intense immune reaction in sensitive individuals upon accidental exposure. (2) If administered under observation, these genotypes could act as a booster for the peanut allergy vaccine recently approved by the U.S. Food and Drug Administration [
15]. (3) These peanuts could also facilitate building resistance against peanut allergens at an early age if consumed under guidance [
19]. Additionally, these genotypes would help the community by reducing the financial burden in maintaining a peanut-free environment, keeping the number of hospitalizations low, and decreasing the need for emergency treatments, such as epinephrine/adrenaline injection.
2.1. Peanut-Germplasm Screening for Reduced Content of Immunogenic Proteins
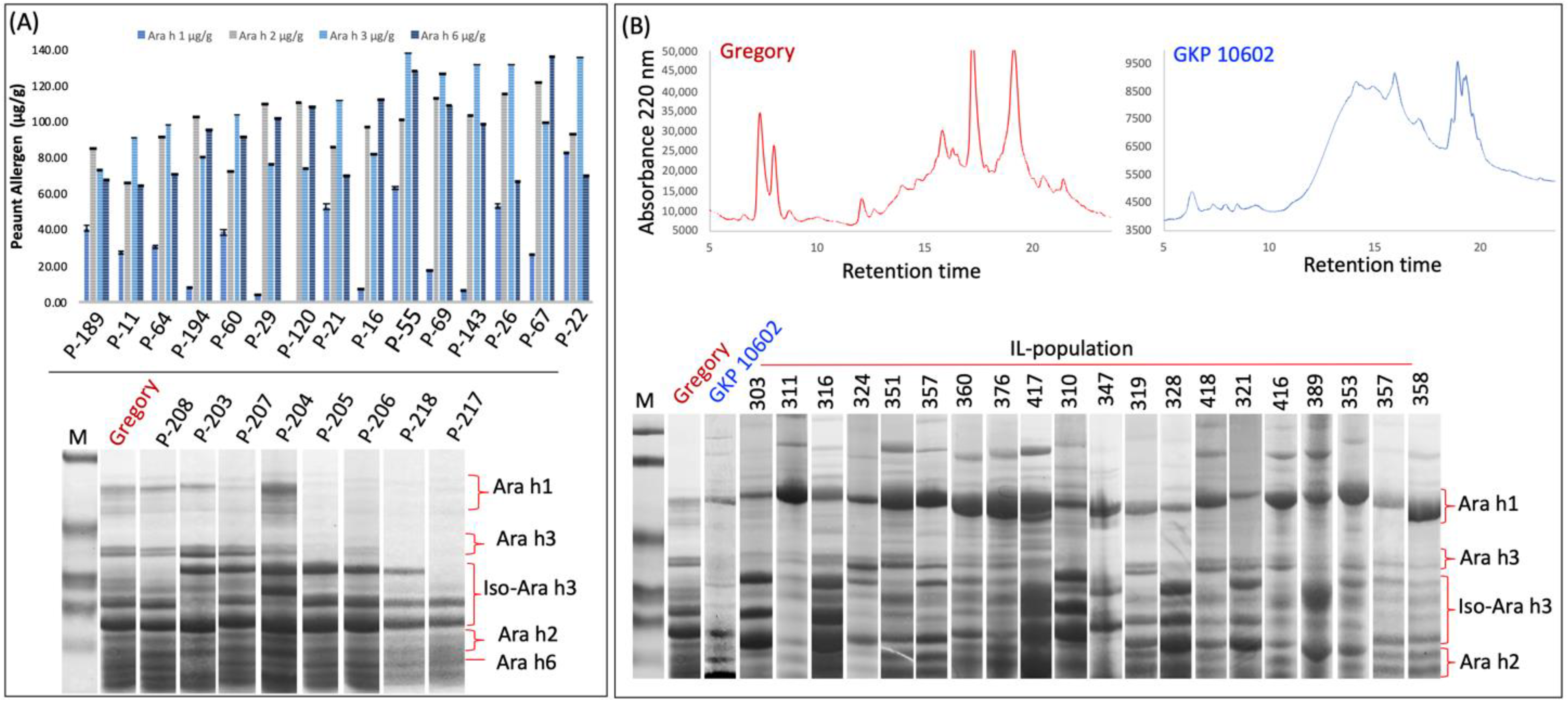
In a screen of peanut germplasm (including USDA and ICRISAT peanut mini-core collections and a set of South Asian lines) for reduced immunogen content, we identified several
Arachis hypogaea genotypes and an
Arachis diogoi line GKP 10602 that can serve as a source of a reduced-immunogenicity trait, as these genotypes exhibited deficiency in several immunogenic proteins (
Figure 2).
A. diogoi is a diploid wild peanut species similar to the ‘A’ sub-genome of the cultivated peanut (
A. hypogaea). Moreover,
A. diogoi was earlier identified as a source of pest and disease resistance, specifically early and late leaf spot and tomato-spotted wilt virus, to name a few [
20], and, hence, was deployed to develop a unique
A. hypogaea (Virginia-type peanut cultivar ‘Gregory’) ×
A. diogoi (accession GKP 10602) introgression population [
21]. This population of 84 introgression lines was characterized using DNA markers, and introgressions of
A. diogoi chromatin in
A. hypogaea chromosomes were identified. These introgressions measured about 3.00% to 18.14% of individual
A. hypogaea chromosomes and involved both peanut sub-genomes [
21]. A preliminary analysis of introgression lines for their seed-storage-protein profiles revealed the inheritance of the reduced-immunogenicity trait from the diploid parent to the tetraploid progeny (recorded as missing protein bands) (
Figure 2). These observations led us to hypothesize that it is possible to identify tetraploid (similar to cultivated peanut) peanut lines with the level of immunogenic proteins similar to the diploid parent,
A. diogoi. Moreover, since this population is genotyped heavily with a 48 K single nucleotide polymorphism (SNP) array, it is possible to map the locations of genes/protein quantity loci that regulate variation in the content of immunogenic proteins and identify DNA markers associated with reduced content of immunogenic-proteins, to facilitate transfer and breeding for this trait. Since different genotypes lacked different immunogenic proteins, it is possible to stack their protein phenotypes via a strategically planned genetic-crossing scheme and selection with protein and DNA markers.
2.2. Peanut Genetic Engineering for Reduced Content of Immunogenic Proteins
In parallel to the germplasm screening, we started efforts to edit out major allergenic proteins from the peanut genome. It required the development of genomic editing reagents for multi-gene editing in peanuts and gene-delivery methods, specifically those leading to the non-transgenic end product. Eliminating allergenic proteins from the peanut genome is complex due to its polyploid nature. As mentioned earlier, out of the 16 known allergenic proteins, four, Ara h1, Ara h2, Ara h3, and Ara h6, are major allergens. Genes for these major allergens map to six peanut chromosomes belonging to three homoeologous chromosome groups. We pulled out sequences of these genes from the public domain, clustered them, and used them for online identification of LbCas12a target sites per gene family, for genome editing using a CRISPR-based multi-gene editing approach [
22]. Before developing the construct for the delivery of the genome-editing reagents, we tested each of these gRNAs in vitro, and the ones performing as desired were assembled in the guide-RNA module. We developed two constructs, one each for the biolistic delivery in the peanut embryo-derived calli and the virus-mediated delivery to intact plants. We are using both of these methods to develop peanut lines with a reduced content of immunogenic proteins, with some initial success.
A somewhat similar effort by other groups, such as the two startups Aranex and IngateyGen, is to target Ara h1, Ara h2, or Ara h3, or all three genes, via the CRISPR approach, is underway. Whereas, Ukko, another startup, is trying to develop engineered peanut proteins for oral immunotherapy. On the other hand, little research has been undertaken so far on the conventional solutions by looking at natural genetic diversity, except for a recent study from ICRISAT, India [
23], where they identify some peanut genotypes reduced in one or more of major allergens.
2.3. Wheat Sensitivity (Celiac Disease, Wheat Allergy, and Non-Gluten Wheat Sensitivity)
Gluten proteins, technically known as prolamins, are the major seed-storage proteins in the wheat grains, which are also one of the most-consumed dietary proteins in the world. On the flip side, unfortunately, gluten is also the primary source of several wheat-associated dietary disorders [
11]. According to an estimate, about 1.4% of the world population is affected by celiac disease, and even a larger proportion is affected by wheat allergy and non-gluten wheat sensitivity [
7]. The only effective therapy is lifelong adherence to an abstinent diet, devoid of wheat and related cereals, such as barley and rye, and hybrid cereals such as Triticale and Tritordeum [
10]. However, this diet is difficult to follow due to the same reasons as discussed for the peanut [
24,
25]. Moreover, this solution is associated with penalties, for instance, (i) strict adherence to a diet totally devoid of gluten-containing grains deteriorates gut health by its negative influence on the gut microbiota, and (ii) long-term adherence to carbohydrate-rich gluten-free diet results in multiple deficiencies and change in patient’s body mass index (BMI) [
9]. Therefore, a gluten-free diet is only recommended to individuals with medically proven gluten sensitivity [
26].
No extant wheat genotype under cultivation or in the wild is safe for celiac patients [
9,
27,
28]. Therefore, in order to develop celiac-safe wheat cultivars, we undertook two approaches: genetic engineering and germplasm screening. The latter approach consists of two different strategies: (i) epigenetic elimination of gliadins and low-molecular-weight glutenins by silencing the wheat genes encoding a DNA glycosylase
DEMETER and an iron-sulfur biogenesis enzyme Dre2, which are responsible for transcriptional de-repression of prolamin genes in the developing endosperm [
29,
30,
31,
32] and (ii) post-translational detoxification of ‘gluten’ proteins by ectopic expression of glutenases in wheat endosperm [
33,
34].
2.4. Wheat Genetic Engineering for Reduced-Gluten Content and to Express Glutenases in Grains
Since gluten is a complex mixture of about 100 proteins encoded by a large number of genes, we devised a clever strategy to deal with this problem by silencing a master regulator of prolamin accumulation in wheat grains [
29]. This strategy is based on the fact that there are two categories of promoters for endosperm-specific gene expression: one that is silenced by methylation in vegetative tissues and must be demethylated before transcription in endosperm, and the other that depends solely on the removal of repressors or induction of endosperm-specific transcription factors [
29]. In wheat, the promoters of LMWgs and gliadins belong to the first category, and the promoters of HMWgs belong to the second category [
29]. It suggests that similar to the high-lysine barley mutant Risø 1508, the gliadins and LMW glutenin subunits in wheat can be eliminated by epigenetic regulation, without affecting the synthesis of HMW glutenin subunits, which are indispensable for baking and are largely non-immunogenic [
29].
Given DEMETER’s vital role in pollen development in Arabidopsis [
35,
36] and in wheat (see below), we adapted an RNA-interference-based method for tissue-specific silencing of the
DME gene. For this purpose, a 938 bp hairpin with a 185 bp stem and a 568 bp loop was synthesized and cloned under the control of the wheat HMWgs promoter. The construct was used for biolistic transformation of wheat scutellar calli, and 118 putative transformants were obtained. Out of these 118 candidates, only 7 plants exhibited a 45–76% reduction in the amount of immunogenic prolamins [
29]. Interestingly, these transformants showed reductions in the amounts of different prolamins. As hairpin construct could lead to the off targeting of genes, we designed the miRNA from the active site and the N-terminal region of the wheat
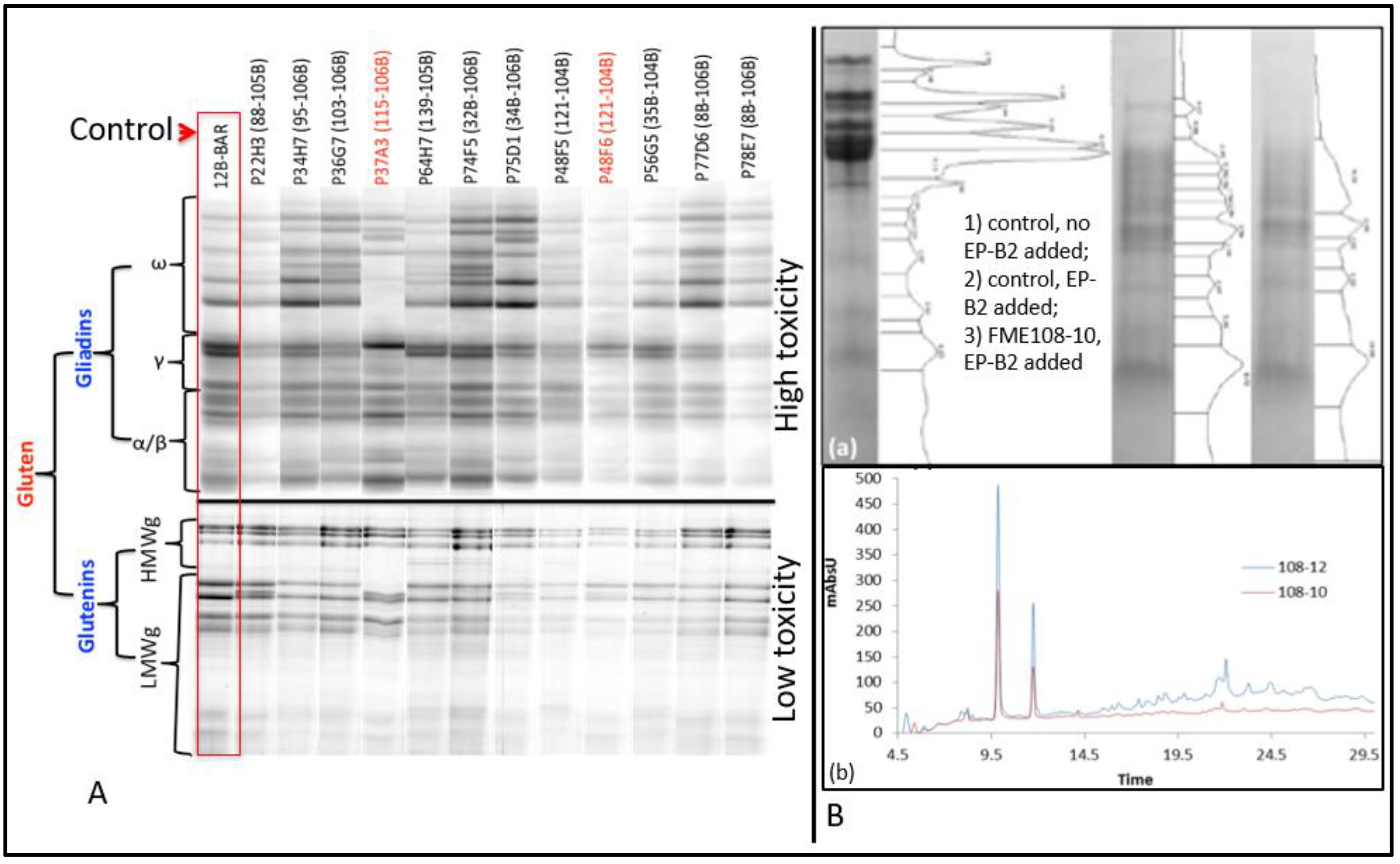
DEMETER homoeologues. Three amiRNA sequences were selected and assembled on a rice MIR528 template using overlapping primers and cloned with a wheat HMWgs promoter. The constructs were delivered to the wheat via a biolistic approach. The selection scheme yielded a total of 215 candidate transformants and 39 lines showing transgene integrations. Out of these 39 putative transformants, 12 lines showed a 40–75% reduction in the amount of immunogenic prolamins (
Figure 3A) [
30,
31]. Protein profiling of these transformants exhibited elimination of specific prolamins and/or prolamin groups. Additionally, these lines showed many desirable characteristics, such as compensatory increases in the amount of HMW glutenin subunits, albumins, and globulins, grain lysine content, and gluten strength [
9].
To develop a dietary therapy for celiac patients, we decided to express glutenases in wheat endosperm [
31,
33,
34]. Based on the parameters, such as target specificity, substrate length, optimum pH, and site of action, a prolyl endopeptidase from
Flavobacterium meningosepticum and a glutamine-specific endoprotease from barley were selected for expression in wheat endosperm [
33]. Wheat transformants expressing these glutenases were obtained, and a few transformants exhibited significant reductions in the amount of indigestible gluten peptides on the tricine PAGE gels and RP-HPLC (
Figure 3B) [
33]. The wheat grains expressing ‘glutenases’ are expected to benefit all consumers due to the improved bioavailability of storage proteins.
2.5. Wheat-Germplasm Screening for Reduced-Gluten Content
Thirty-four wheat landraces and twenty-seven ABA-biosynthesis or hypersensitive mutants (ABH) and ABA-signaling or insensitive mutants (ABI) were tested for their protein profiles, using SDS-PAGE-based densitometric analysis and high-performance liquid chromatography [
37]. Gluten-banding patterns from the genotypes of a landrace or across landraces were compared, and the mutant types were identified. Mutants with different missing gluten proteins were selected for the genetic crossing, to stack their effects on gluten content into a single genotype. The differences in the banding pattern observed in these genotypes were divided into two categories: qualitative differences and quantitative differences. The deficiency of a protein subunit (qualitative differences) was determined by comparing the protein profile of different genotypes of a landrace (analyzed at three to four half-seeds per landrace). On the other hand, the quantitative differences were determined via a densitometric analysis of the corresponding bands (determined based on molecular weights) among the genotypes belonging to a landrace. Similar criteria were used to study the quantitative and qualitative differences in the protein profiles of the wheat ABA-insensitive and -hypersensitive mutants, except, in this case, comparisons were made with the respective wild-type controls [
37].
Based on the knowledge that the enzyme coded by the wheat
DEMETER gene is responsible for the transcriptional activation of the gluten genes in developing wheat grains [
29], we screened two TILLING populations, one each in the ‘Kronos’ and ‘Express’ backgrounds, for mutations in the
DEMETER genes [
31,
32]. To reduce the workload, we confined our search for the mutants to one kb region in the active site of the
DEMETER genes. Screening for mutations in the ‘Kronos’ and ‘Express’ populations, respectively, yielded 77 and 149 mutations. The single mutations in the
DEMETER homoeologues showed reductions in specific prolamins [
32]. Crosses were made between the mutants carrying substitutions, premature stop codons, and splice-site variations in A and B sub-genome
DEMETER homoeologues, to pyramid the effect of the individual mutations in a single genotype. Unfortunately, the crosses between the mutants carrying the premature stop codons or splice-site variations never yielded a double mutant [
31,
32]. To determine the cause of the observed segregation distortion, we closely monitored the anther morphology, number of viable pollens, and pollen germination. We observed that the nonsense mutations in wheat
DEMETER genes have significantly reduced the anther locule size and the number of viable pollen and negatively affected the pollen germinate. Thus, this analysis also suggested a vital role of
DEMETER in pollen development and germination [
31,
32,
37].
Dre2, or Derepressed for Ribosomal protein S14 Expression, facilitates the deposition of the iron-sulfur (Fe-S) cluster to the DEMETER apoenzyme, which is vital for its interaction with genomic DNA and its subsequent demethylation [
38]. Given Dre2′s role in DEMETER activation, we decided to test its effect on prolamin accumulation [
37]. As a first step, the TILLING population in the ‘Kronos’ background was screened for mutations in the wheat
Dre2 homoeologues. Thirteen mutants in the 2AL copy, and three mutants in the 2BL copy of the
Dre2 gene were identified [
32,
37]. The single mutations in the
Dre2 homoeologues showed reductions in the specific prolamins. These single mutations are currently being crossed in the greenhouse to pyramid their effects. During these attempts, it was continuously noticed that the anther size for the Dre2-2AL mutants was significantly smaller than the wild-type control [
32].
Similar efforts are underway in different parts of the world to develop reduced-immunogenicity wheat genotypes suitable for celiac patients and wheat-allergic individuals [
9,
27,
28,
39]. Some noteworthy efforts from the public sector were from Francisco Barro’s group at the Institute for Sustainable Agriculture, Córdoba, Spain; Susan Altenbach’s group at the Western Regional Research Center, USDA-ARS, Albany, CA, USA; Hetty van den Broeck’s group and Marinus Smulders’s group at Wageningen University & Research, The Netherlands; and Domenico Lafiandra’s group at the University of Tuscia, Italy. Similar remarkable efforts are underway in the private sector at Calyxt in Roseville, MN, USA; Arcadia Biosciences in Davis, CA, USA; and the Bayer-supported startup Ukko in Rehovot, Israel. The earlier efforts primarily focused on pyramiding the reduced-gluten wheat mutants (natural and induced) and the later efforts focused on genetic engineering (including RNA interference and gene editing). However, the different efforts differed in the genes targeted and the adopted strategies. For instance, some focused on editing the specific motifs (epitopes) in gluten proteins rather than eliminating them. Recently, via their persistent efforts, Barro and co-workers demonstrated that a reduced-gliadin RNAi line (E82) did not elicit an immune response in celiac patients under a pilot, blind, randomized crossover gluten challenge [
40]. It gives hope that the reduced-immunogenicity wheat genotypes in the pipeline will become helpful for celiac patients.
3. Public Perspective
We focus on developing reduced-immunogenicity wheat and peanut genotypes that can significantly reduce the scope of severe reaction to accidental exposure to wheat/peanut proteins. However, one major question that we often encounter is the need for reduced immunogenicity crops, while gluten-/peanut-free products are already an option. There are several reasons to pursue this line of research: (i) for many in low-income countries, a gluten/peanut-free diet is not an option due to the unavailability of these products or financial reasons. This is particularly true in the case of sustenance farmers, who also are limited by the choice of crops they can grow, either due to market-driven reasons or adaptability of crops in their region. (ii) Removing cereals (wheat, barley, and oats) or peanuts from the diet deprives the consumer of many other essential nutrients, such as fibers, antioxidants, vitamins, and minerals. Indeed, several recent studies suggested that removing wheat from the diet, in the long run, enhances the risk of developing colorectal cancer due to a deficiency of the fermentable fibers and essential phenolic compounds from the diet, and long-term adherence to a gluten-free diet increases the chances of adverse changes in the body mass index and composition of the gut microbiota [
9,
28]. Further, removing wheat and peanut and cross-reacting grains from the diet limits vegetarians’ and vegans’ options to get the required level of dietary proteins. On the other hand, the reduced immunogenicity genotypes of wheat and peanuts could bring back the balance in the lives of individuals with wheat/peanut sensitivities, who are forced to remove these grains/nuts from their diets under the current regimen. As mentioned earlier, these genotypes will retain conventional wheat and peanut benefits, while not triggering an immune reaction. Additionally, these genotypes will reduce the risk of accidental exposure to allergens and reduce, upon exposure, a severe response, lowering consumers’ financial burden. Especially in a vegetarian/vegan diet, limited by choice, these genotypes will reintroduce the option of incorporating wheat and peanut. Further, these reduced immunogenicity lines can be effectively utilized to develop early immunity in children and even adults in supervised settings, specifically individuals vaccinated for peanut allergy. Under controlled settings and after necessary testing, the measured doses of reduced-immunogenicity wheat/peanut genotypes could be administered as vaccines.
Another big question that often arises is the fear of spoiling palatability/flavor, nutritional quality, processing attributes, or field performance of wheat and peanut, by making alterations in their protein composition. Earlier studies have addressed this, to some extent. For instance, Jianmei Yu and co-workers treated roasted/raw peanut kernels with a cocktail of digestive enzymes. The process solubilized (removed) 100% of the Ara h1 proteins and 98% of the Ara h2 proteins, essentially producing Ara h1- and Ara h2-free peanut kernels [
41]. After analysis, the authors of this work concluded that the enzymatic removal of immunogenic proteins produces hypoallergenic kernels, without an adverse effect on peanut’s nutritional quality and flavor. In another study, using RNA interference, Hortense Dodo and group silenced three major allergen genes: Ara h1, Ara h2, and Ara h3 [
16]. This study concluded that the transgenic line significantly reduced in Ara h1, Ara h2, and Ara h3 content is on par in its agronomical performance and end-use quality with the untransformed control. After field testing, this line was named ‘Serenity’ and is nearing its release later this year. However, with the genotype being transgenic, its public acceptance is debatable. Further, Peggy Ozias-Akins and co-workers demonstrated that contrary to their suspicion, the suppression of Ara h2 (a trypsin inhibitor) and Ara h6 did not render peanut susceptible to soil-born fungi,
Aspergillus spp., indicating that elimination of immunogenic proteins from peanut kernels is a viable approach [
18]. In peanuts, Ara h1, Ara h2, and Ara h3 cumulatively represent 30% of the total seed proteins [
42], where Ara h1 constitutes 12–16% and Ara h2 constitutes 6–9% of it [
5]. Thus, Ara h6 accounts for only a tiny amount of grain proteins. To sum up, the major allergens constitute about 30% of the grain proteins, and based on the existing knowledge, we anticipate that other proteins or free amino acids would compensate for their loss; hence we do not expect these changes to reduce the peanut’s nutritional quality or flavor.
Similar studies were performed in wheat. These studies suggested that eliminating proteins from the wheat grains does not lead to a gross reduction in the protein content, since it generally stays the same as other proteins compensate for the loss [
9]. Further, studies were performed in different market classes of wheat to determine the effects of eliminating specific glutenins and gliadins on their end-use performance. For instance, Kieffer and co-workers demonstrated that it was possible to bake a bread with reasonable crumb and crust structures, after eliminating all gluten except the HMW subunits from the wheat flour. This was shown by baking bread from a mixture of the washed-out wheat-flour residues (containing starch, soluble protein, fat, fiber, and minerals) and known quantities of recombinant HMW subunits (1Dx5 and 1Dy10). The dough kneaded from the mixture showed good elasticity and, after baking, resulted in bread rolls with reasonable volume and internal structure [
43]. Similar conclusions were reached when flour, derived from wheat transformants lacking one or more families of the gluten proteins, was baked into normal-looking loaves with acceptable organoleptic properties [
44]. Additionally, the authors of this study reported that silencing the glutamine and proline-rich gluten proteins from wheat grains consequently increases the grain-lysine content, due to the compensation of the missing gluten proteins via the more balanced albumins, globulins, and free amino acids. This is an observation that was concurred by the high-lysine barley mutants (
lys3a and
lys1), which exhibit reduced hordein (gluten homolog in barley) content [
9,
28]. Collectively, these studies suggested that deletions in genes encoding different gluten proteins, in general, bake into bread/cookies with reasonable textural and organoleptic properties. Additionally, further improvement in the functional properties of reduced-gluten wheat flours could be achieved by blending them with the proteins of non-toxic cereals or non-cereals [
9,
28].




{kind=link}
{kind=link}
{kind=link}


