Abstract
In this paper, we develop a three-species food web model that incorporates the use of interactions between diseased predator–prey models. The logistically growing prey populations are susceptible and diseased prey. Prey populations are assumed to grow logistically in the absence of predators. We investigate the effect of fear on susceptible prey through infected prey populations. In Crowley–Martin-type interactions, it is assumed that interdependence between predators happens regardless of whether an individual predator is searching for prey or handling prey at the time. Also, the prey harvesting of susceptible and infected prey has been considered. The existence of all possible equilibrium points for biological systems has been established. The criteria for the local and global stability of equilibrium points are examined. Additionally, we look at Hopf-bifurcation analysis for the suggested model in relation to the existence of harvesting rate . Numerical simulations are provided in order to explain the phenomenon and comprehend the complex interactions between predators and prey.
1. Introduction
The predator–prey models developed by Lotka [1] and Volterra [2] are regarded as the earliest developments in contemporary mathematical ecology in coupled systems of non-linear differential equations. Since Kermack and Mckendrick’s pioneering work on SIRS, [3], epidemiological models have attracted much interest from researchers. Mathematical modeling of predator–prey interactions, known as “functional response,” is among the most important factors in predator–prey population modeling. Crowley–Martin functional responses take into account both prey and predators. In the recent era, some eminent authors, [4,5,6,7], have studied to understand the importance and interactions of prey. To make the model system more realistic and feasible in the ecosystem, they incorporated some functional responses, i.e., Crowley–Martin-type functional responses. Kadhim and Azhar [8] use a type II Holling function to represent two disease types in a predator population model with a linear functional response. In [9], a nonlinear analysis of a discrete effects predator–prey model is investigated. Prey refuge and prey harvest [10,11], with ratio-dependent and Holling type II functional responses. Several investigations have been conducted on the dynamic behavior of Crowley–Martin diseased predator–prey models. To our knowledge, only a few researchers have looked into three-species prey–predator models that take into account species interactions, including Crowley–Martin disease in prey populations. This study examines how fear affects a Crowley–Martin eco-epidemiological model with prey harvesting. The rest of the paper is structured as follows: In Section 2, we describe how the study’s model formation was created. In Section 3 and Section 4, we talk about boundary equilibrium points and their stability. In Section 5, the Hopf-bifurcation is the positive equilibrium point . Numerical simulations of the suggested model are examined in Section 6. The paper is concluded in Section 7, which also discusses the biological consequences of our mathematical findings.
2. Mathematical Model Formation
The model explains the relationship between the structure of the infected prey and the following equations. The proposed framework was used to discover a non-linear prey–predator mathematical model.
and the positive conditions are described as and .
The Table 1 displays the specific biological meanings of the parameters.

Table 1.
Biological representation of the model.
The condition for the impact of fear is . This refers to the infected’s fear effect on susceptible prey. Here, f is the amount of fear. It is appropriate to modify the variables as follows in order to decrease the number of systems (1) variables , and to consider the dimension time . Now, we apply the following transformations. Equation (1) can be represented in dimensionless form using the above transformations.
3. Existence of Equilibrium Points
The model (2) exhibits the following equilibrium points based on observation:
- is the trivial equilibrium point.
- is the boundary equilibrium point exists if , where .
- is the without predator equilibrium point, where Here, is the unique positive root of the equation, withIt is observed that is the unique positive root if and . exists for .
- The infection-free equilibrium point , where Thus, the conditions must exist for the infection-free equilibrium point are and (assume and .
- The endemic equilibrium point , where Thus, the conditions must exist for the endemic equilibrium point are , , .
4. Stability Analysis
In order to determine local stability around various equilibrium points, we compute the Jacobian matrix. At each given point , the Jacobian matrix is given by
where,
Theorem 1.
The trivial equilibrium point is always unstable.
Proof.
Here, the eigenvalues of are , , . Hence, is locally asymptotically stable only if and unstable otherwise. □
Theorem 2.
is locally asymptotically stable if and .
Proof.
where Therefore, eigenvalues of are . If i.e., , , i.e., and , i.e., . Thus, is locally asymptotically stable if and . □
Theorem 3.
is locally asymptotically stable if , and .
Proof.
where,
Therefore, the characteristic equation of is , where and .
In the above characteristic equation, we obtain one of the eigenvalue is , which is negative as and the other two eigenvalues should be negative if and .
Hence, is locally asymptotically stable if , and . □
Theorem 4.
The diseased-prey free equilibrium point is locally asymptotically stable if , and .
Proof.
where Now, the characteristic equation for is
, where
In the above characteristic equation, we obtain one if the eigenvalue is , which is negative as and the other two eigenvalues should be negative if and . Therefore, is locally asymptotically stable if , , , otherwise the system (2) will be unstable. □
Theorem 5.
is locally asymptotically stable if , , and .
Proof.
where,
The characteristic equation is
where
According to the Routh–Hurwitz criterion, , , and must all be positive, and the characteristic of all the roots must be negative. Hence, is locally asymptotically stable. □
Global Stability Analysis
Theorem 6.
If is the endemic equilibrium point is globally asymptotically stable in .
Proof.
A positive Lyapunov function is defined as where are positive constants. Here, since for and .
Differentiating with respect to t, we obtain
Now, we see that . Whenever and Consequently, for all solutions in G, L is a Lyapunov function. □
5. Hopf-Bifurcation Analysis
Theorem 7.
If the critical value for the bifurcation parameter is exceeded, the model (2) will experience the Hopf-bifurcation. The following Hopf-bifurcation requirements are present for ,
- 1.
- 2.
- where is the zeros of the characteristic equation corresponding to the non-negative equilibrium point.
6. Numerical Analysis
We show some numerical simulations of the model (2) in this section. To accomplish this, we use Diethelm et al.’s predictor–corrector approach to solve the proposed model. The system (2) parameter values are . From Theorem 5, the positive equilibrium point exists for , and is locally asymptotically stable.
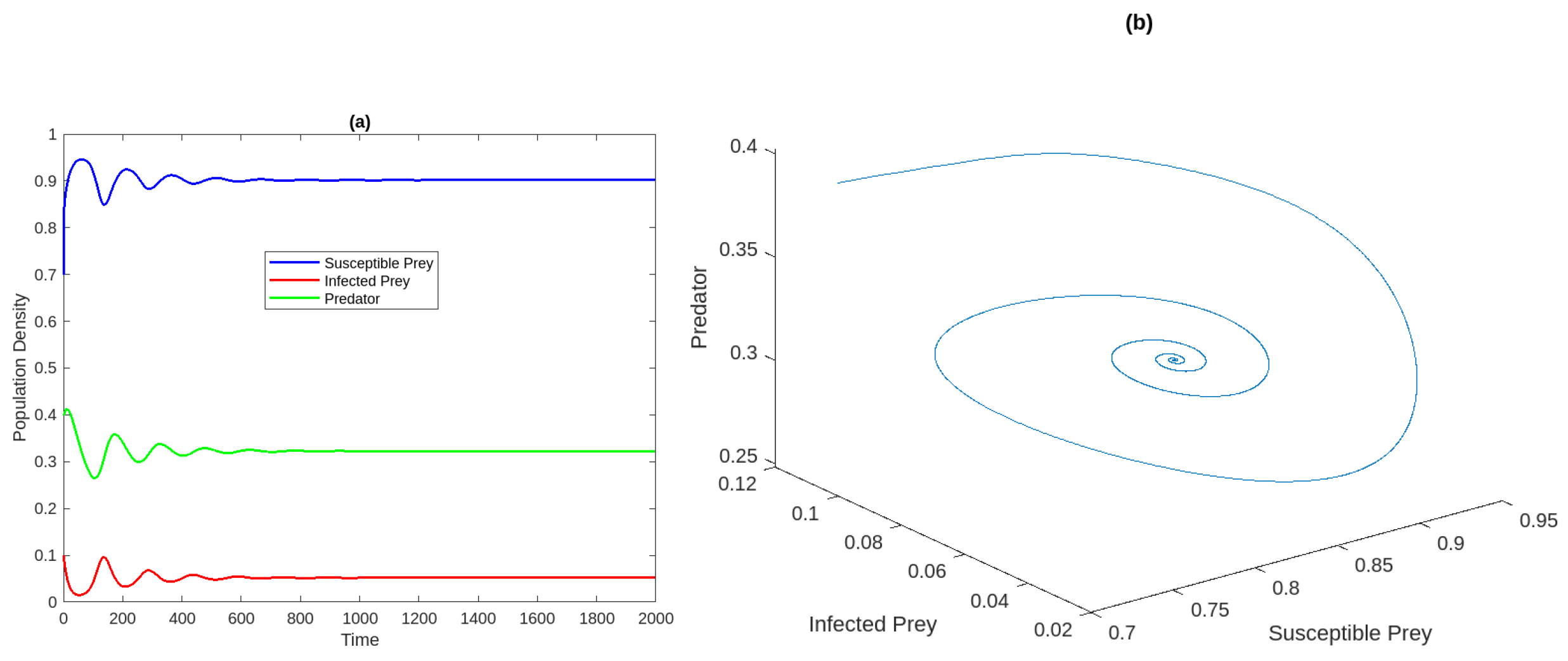
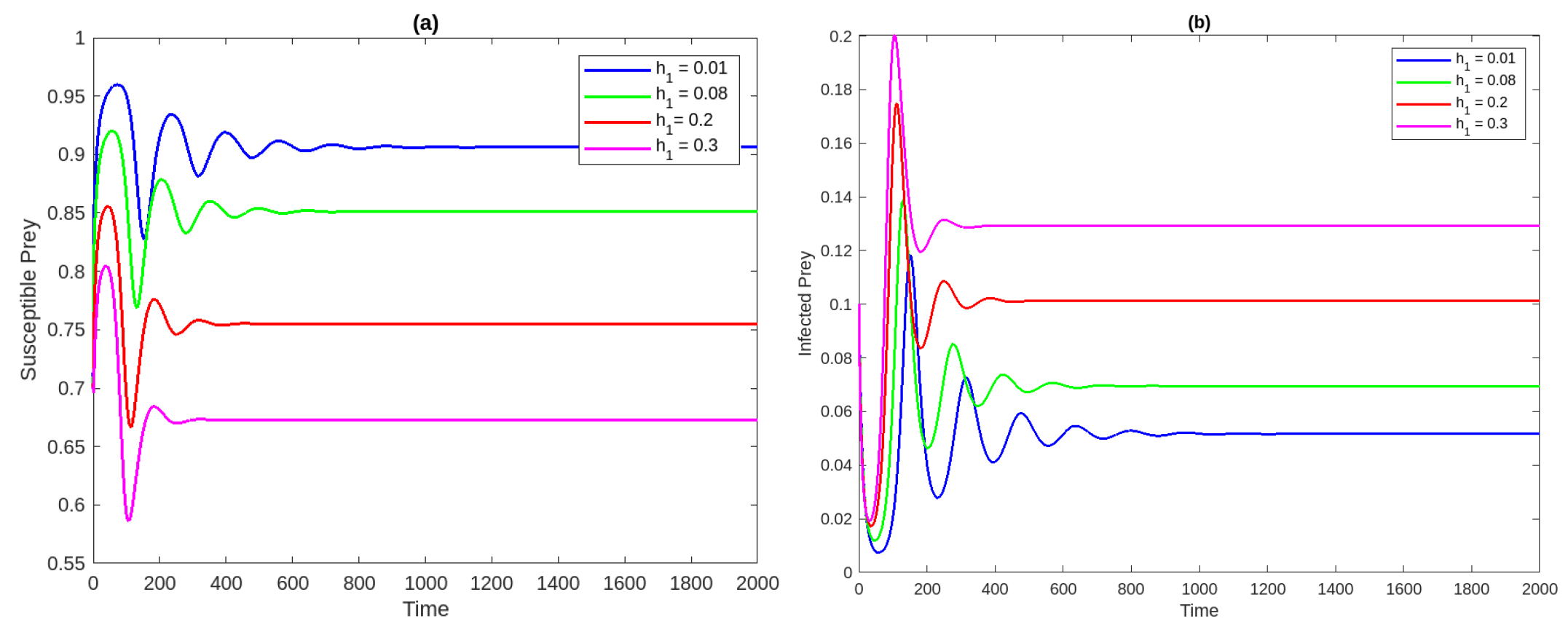
Figure 1a Time analysis for the system (2) for . Figure 1b Phase portrait of the system at . Figure 2a,b Susceptible and infected prey populations with different values for It shows that increasing the harvesting rate of susceptible prey leads to a decrease in the population of vulnerable prey and predators while increasing the population of diseased prey.
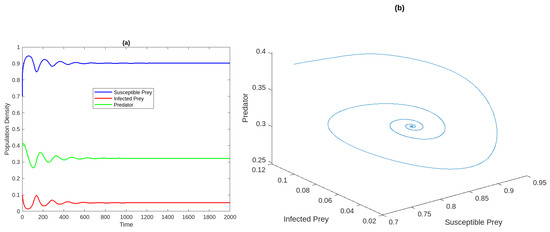
Figure 1.
(a) Time analysis for the system (2) for . (b) Phase diagram of the model system .
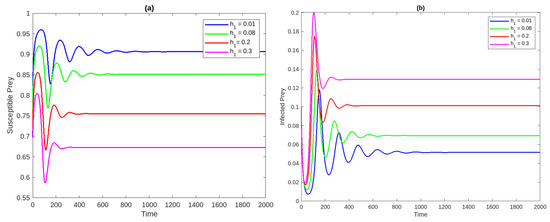
Figure 2.
The population concentrations of susceptible prey (a) and infected prey (b) populations with different values for
7. Conclusions
We investigated a three-species food-web model that involved the use of the interactions between diseased prey–predator model. The local and global stability of (2) is used for each set of biologically possible equilibrium points in the model. It is used to modify the harvesting rate and the level of fear as control parameters. In addition, we investigated the stability analysis of the model (2) and studied the Hopf-bifurcation phenomenon. As a result, we found that modifying the harvesting rate significantly affects the stability of the system (2). The analytical and numerical findings demonstrate that the harvesting rate has a significant impact on every population. A decrease in the population of susceptible prey and an increase in infected prey population density are the effects of increasing the harvesting rate. This study shows the complex behavior of the proposed model.
Author Contributions
D.A., S.M., A.A. and S.P.M. contribute equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lotka, A.J. Elements of Physical Biology; Williams and Wilkins: Philadelphia, PA, USA, 1925. [Google Scholar]
- Volterra, V. Variazioni e fluttuazioni del numero d’individui in specie animali conviventi mem. accad. lincei roma 2 31; fluctuations in the abundance of a species considered mathematically. Nature 1926, 118, 558. [Google Scholar] [CrossRef]
- Kermack, W.O.; McKendrick, A.G. A contribution to the mathematical theory of epidemics. Proc. R. Soc. Lond. Ser. Contain. Pap. Math. Phys. Character 1927, 115, 700–721. [Google Scholar]
- Dubey, B.; Agarwal, S.; Kumar, A. Optimal harvesting policy of a prey–predator model with crowley– martin-type functional response and stage structure in the predator. Nonlinear Anal. Model. Control. 2018, 23, 493–514. [Google Scholar] [CrossRef]
- Maiti, A.P.; Jana, C.; Maiti, D.K. A delayed eco-epidemiological model with nonlinear incidence rate and crowley—Martin functional response for infected prey and predator. Nonlinear Dyn. 2019, 98, 1137–1167. [Google Scholar]
- Tiwari, V.; Tripathi, J.P.; Abbas, S.; Wang, J.; Sun, G.; Jin, Z. Qualitative analysis of a diffusive crowley–martin predator–prey model: The role of nonlinear predator harvesting. Nonlinear Dyn. 2019, 98, 1169–1189. [Google Scholar] [CrossRef]
- Maiti, A.P.; Dubey, B.; Chakraborty, A. Global analysis of a delayed stage structure prey–predator model with crowley–martin type functional response. Math. Comput. Simul. 2019, 162, 58–84. [Google Scholar] [CrossRef]
- Kadhim, A.J.; Majeed, A.A. Epidemiological model involving two diseases in predator population with holling type-ii functional response. Int. J. Nonlinear Anal. Appl. 2021, 12, 2085–2107. [Google Scholar]
- Vinoth, S.; Sivasamy, R.; Sathiyanathan, K.; Rajchakit, G.; Hammachukiattikul, P.; Vadivel, R.; Gunasekaran, N. Dynamical analysis of a delayed food chain model with additive allee effect. Adv. Differ. Equations 2021, 2021, 54. [Google Scholar] [CrossRef]
- Ashwin, A.; Sivabalan, M.; Divya, A.; Siva Pradeep, M. Dynamics of Holling type II Eco-epidemiological model with fear effect, prey refuge, and prey harvesting. In Proceedings of the 1st International Conference on Mathematics and Applications, Online, 1–15 May 2023. [Google Scholar]
- Divya, A.; Sivabalan, M.; Ashwin, A.; Siva Pradeep, M. Dynamics of ratio dependent eco epidemiological model with prey refuge and prey harvesting. In Proceedings of the 1st International Conference on Mathematics and Applications, Online, 1–15 May 2023. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).