
Proteomic Blueprint of Atlantic Cod (Gadus morhua) Otoliths Revealing Environmental Stress Insights through Label-Free Quantitative Shotgun Proteomics

 , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Otoliths
2.2. Chemicals and Standards
2.3. Otolith Protein Extraction
2.4. Shotgun Proteomics by LC-ESI-MS/MS
2.5. Quantitative Analysis
Quantitative Expressed Protein Intensities Accuracy
2.6. Statistical Analysis
2.7. Bioinformatics Analyses
3. Results and Discussion
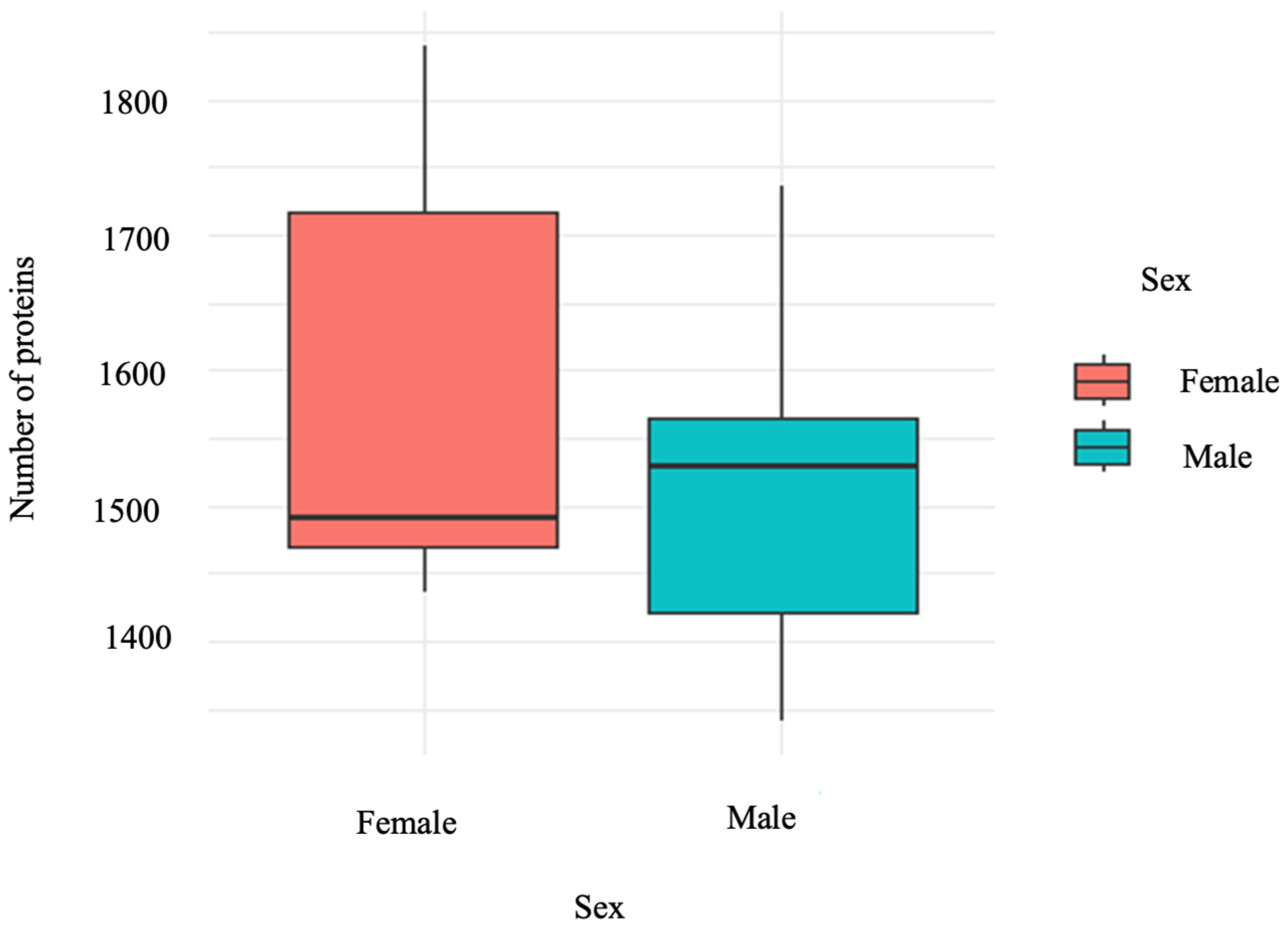
3.1. Protein Expression between Sexes
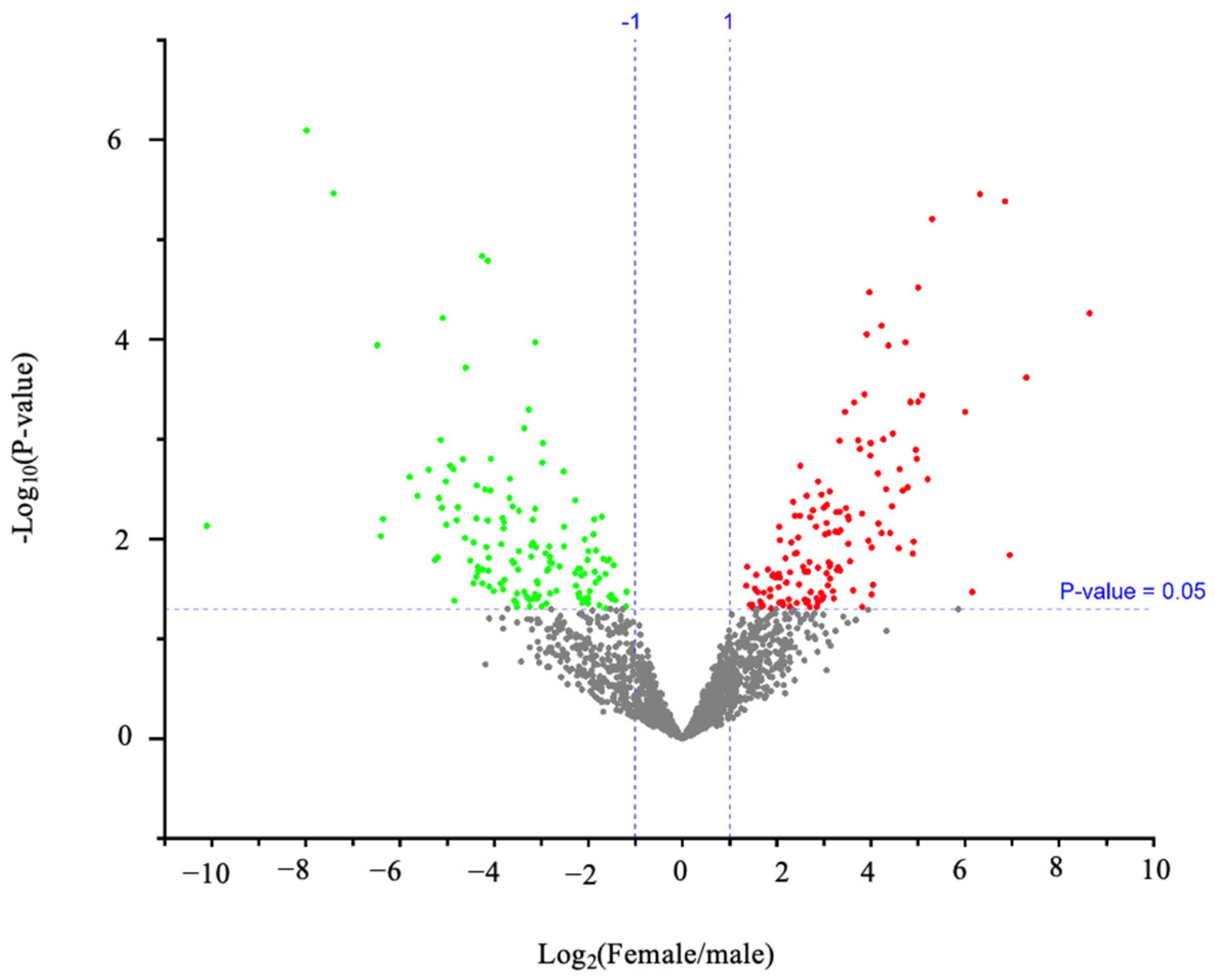
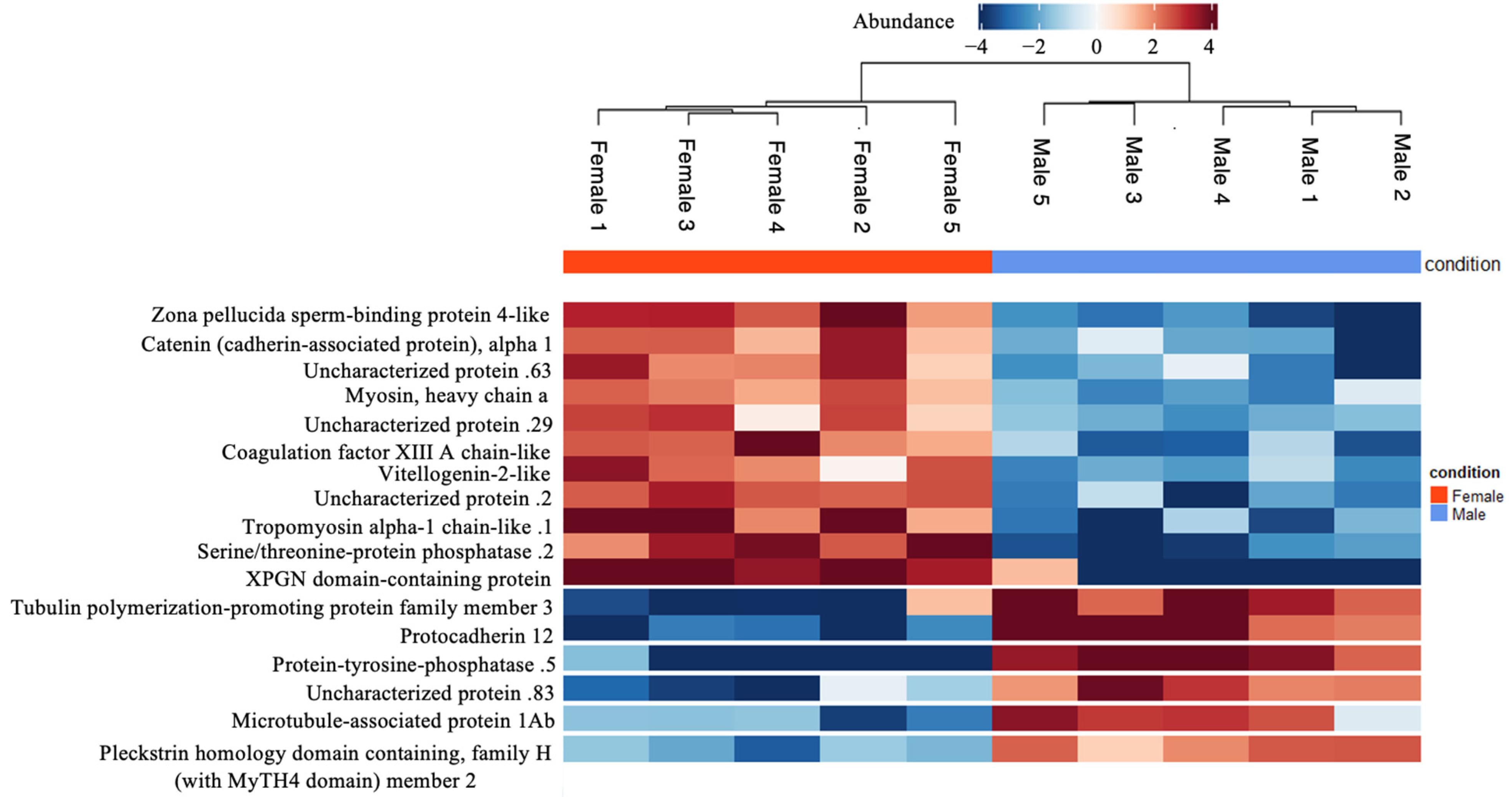
3.1.1. Otolith Proteins with Substantial Female-Biased Expression
3.1.2. Otolith Proteins with Substantial Male-Biased Expression
3.2. Quantitative Analysis of the Total Protein Profile
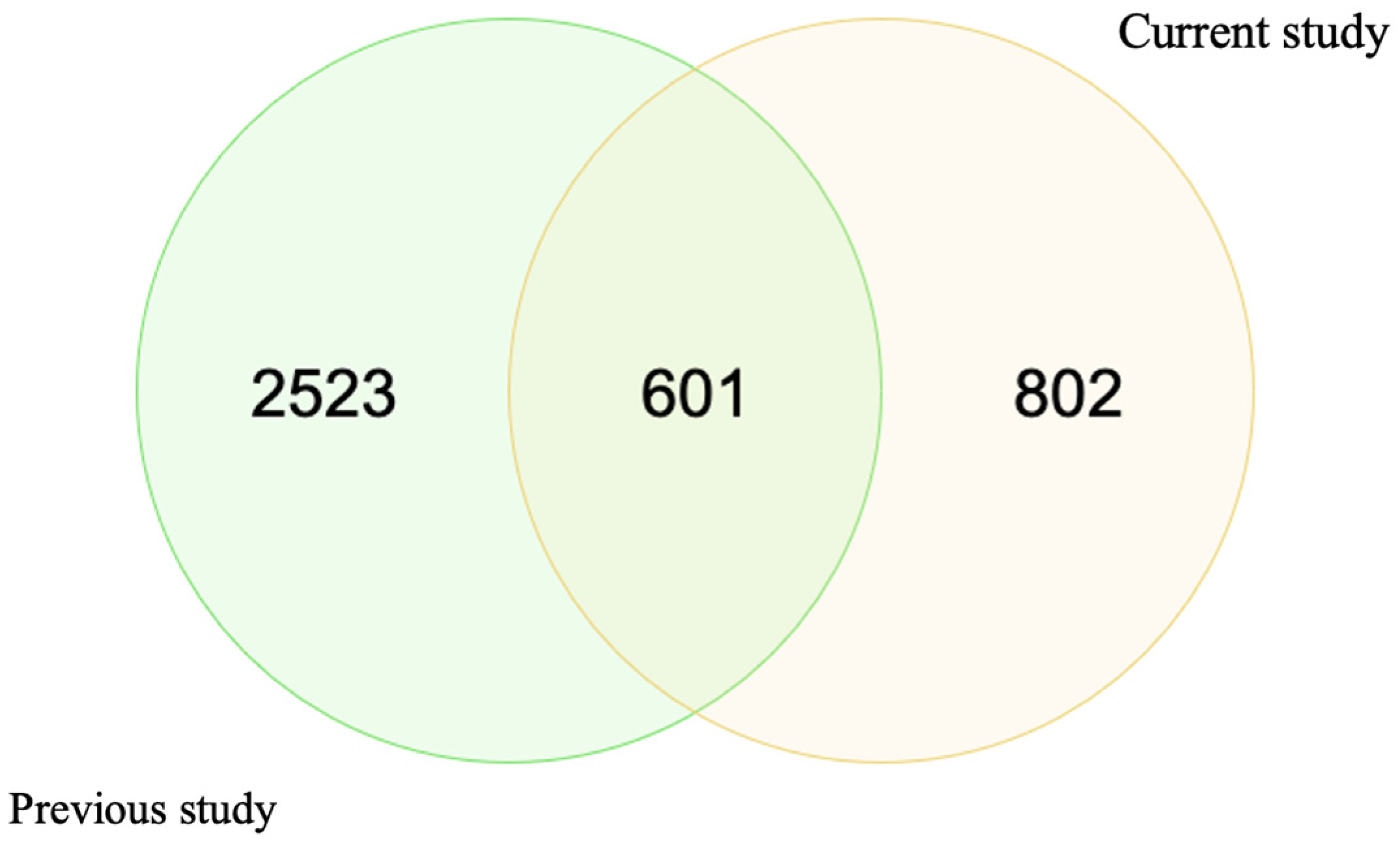
3.2.1. Identified Otolith Protein Profile Common in Both Sexes
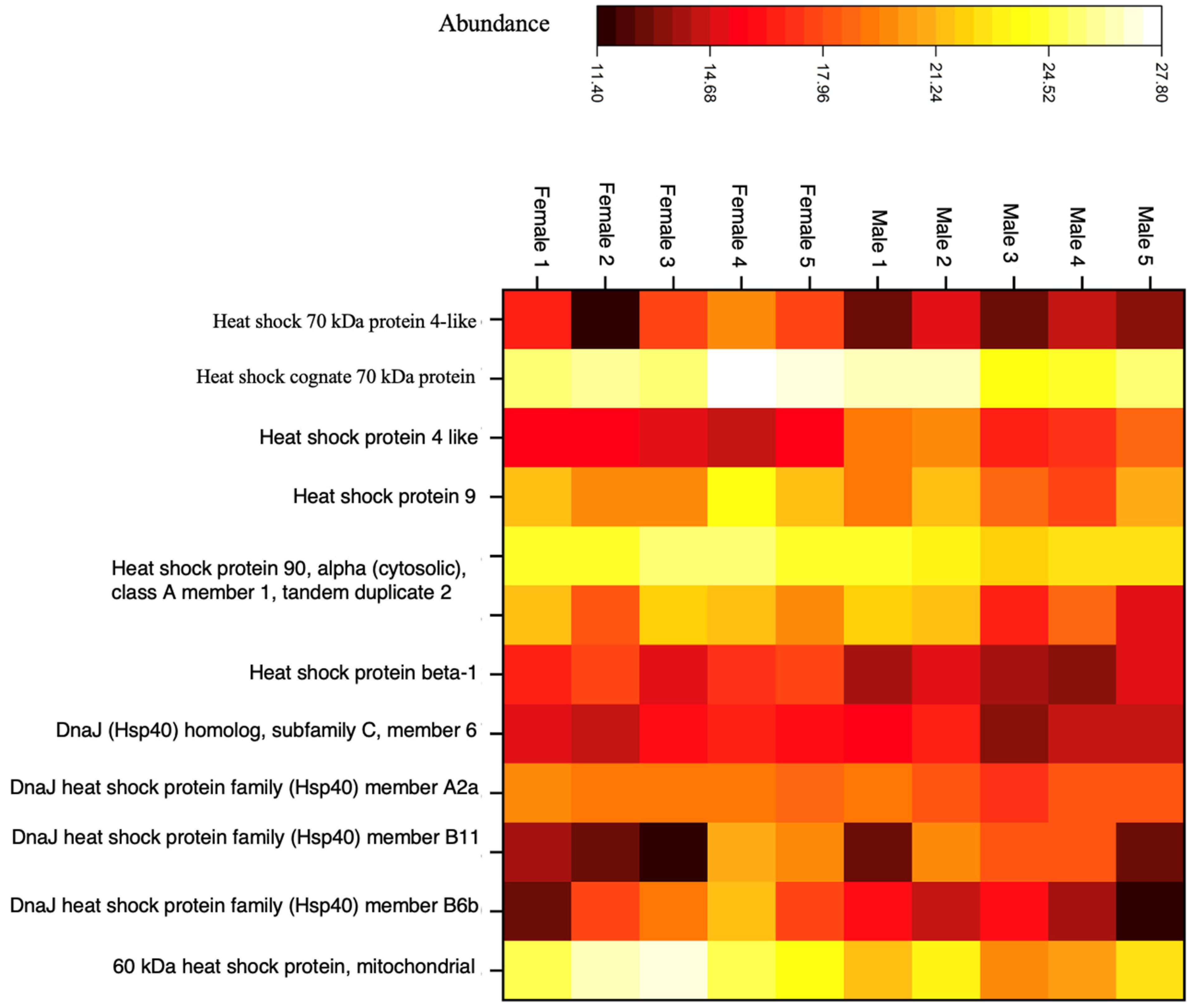
Otolith Heat Shock Proteins
Identification of Otolith Protein Bioindicators of Endocrine Disruptors
3.3. Bioinformatics Analyses
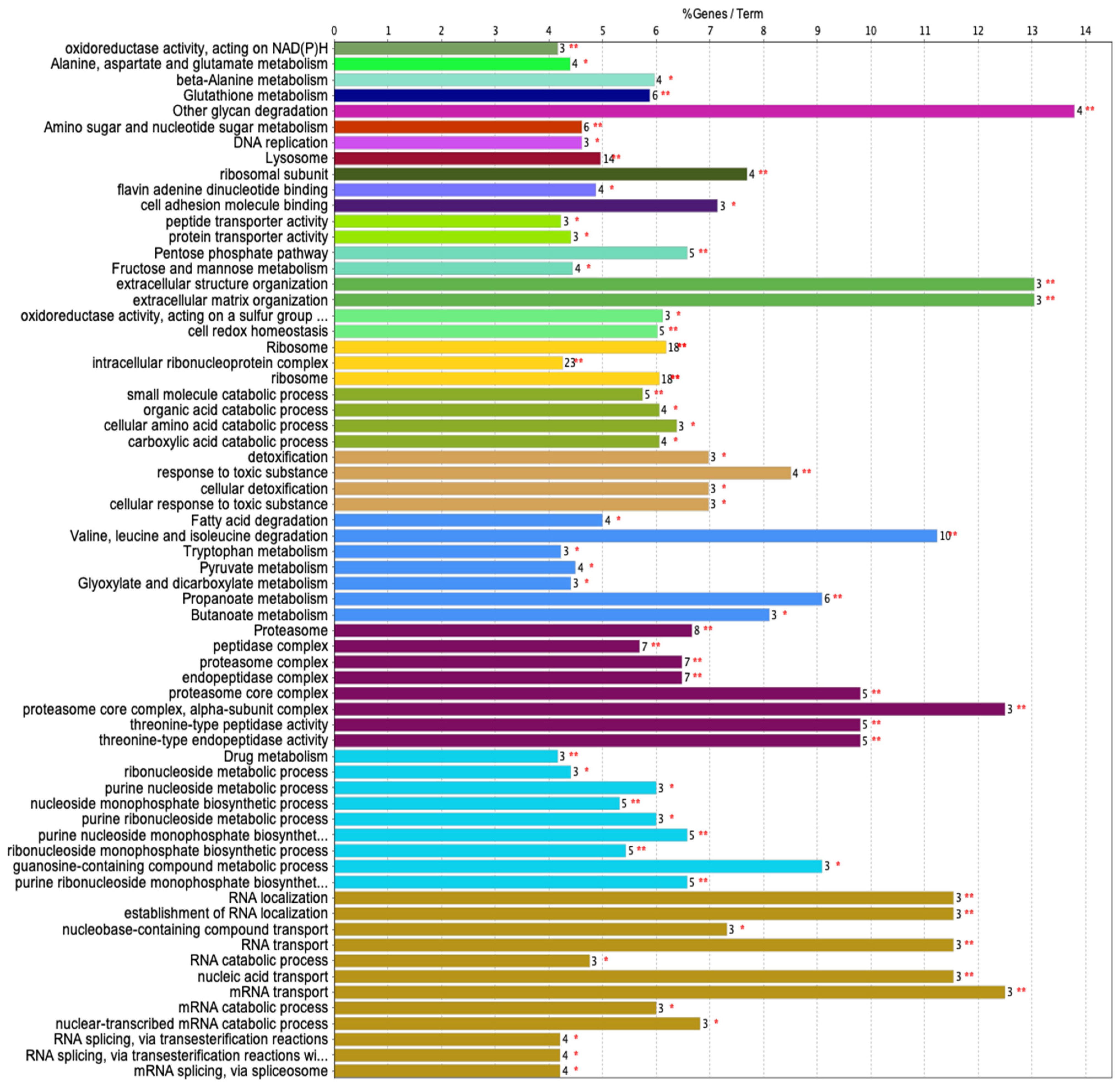
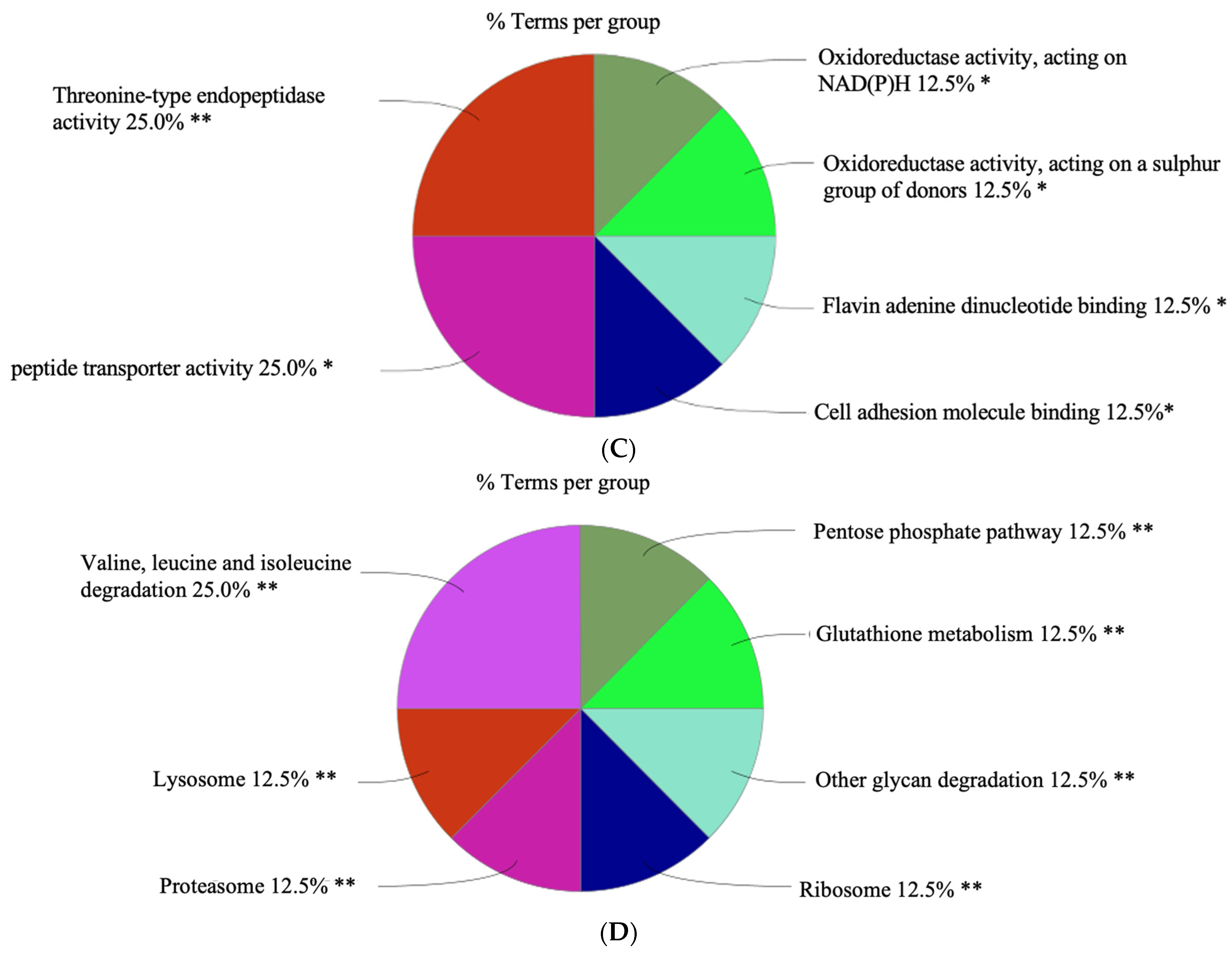
3.3.1. Gene Ontology (GO)
3.3.2. Protein–Protein Interaction (PPI) Analysis
4. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Norin, T.; Canada, P.; Bailey, J.A.; Gamperl, A.K. Thermal biology and swimming performance of Atlantic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus). PeerJ 2019, 7, e7784. [Google Scholar] [CrossRef]
- Pinhorn, A.T. Fishery and biology of Atlantic cod (Gadus morhua) off the Southwest coast of Newfoundland. J. Fish. Res. Board. Can. 1969, 26, 3133–3164. [Google Scholar] [CrossRef]
- Meidell, L.S.; Carvajal, A.K.; Rustad, T.; Falch, E. Upgrading marine oils from cod (Gadus morhua) on-board the deep-sea vessels-from waste to value. Foods 2023, 12, 1659. [Google Scholar] [CrossRef]
- Hawkins, A.D.; Popper, A.N. Sound detection by Atlantic cod: An overview. J. Acoust. Soc. Am. 2020, 148, 3027. [Google Scholar] [CrossRef]
- Dauphin, Y.; Dufour, E. Composition and properties of the soluble organic matrix of the otolith of a marine fish: Gadus morhua Linne, 1758 (Teleostei, Gadidae). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 134, 551–561. [Google Scholar] [CrossRef]
- Murayama, E.; Takagi, Y.; Ohira, T.; Davis, J.G.; Greene, M.I.; Nagasawa, H. Fish otolith contains a unique structural protein, otolin-1. Eur. J. Biochem. 2002, 269, 688–696. [Google Scholar] [CrossRef]
- Lewis, L.S.; Huang, J.L.; Willmes, M.; Fichman, R.A.; Hung, T.-C.; Ellison, L.T.; Stevenson, T.A.; Teh, S.J.; Hammock, B.G.; Schultz, A.A.; et al. Visual, spectral, and microchemical quantification of crystalline anomalies in otoliths of wild and cultured delta smelt. Sci. Rep. 2022, 12, 1–12. [Google Scholar]
- Degens, E.T.; Deuser, W.G.; Haedrich, R.L. Molecular structure and composition of fish otoliths. Mar. Biol. 1969, 2, 105–113. [Google Scholar] [CrossRef]
- Payan, P.; De Pontual, H.; Bœuf, G.; Mayer-Gostan, N. Endolymph chemistry and otolith growth in fish. Comptes Rendus. Palevol. 2004, 3, 535–547. [Google Scholar] [CrossRef]
- Ferri, J. Otoliths and Their Applications in Fishery Science. Fishes 2023, 8, 35. [Google Scholar] [CrossRef]
- Smolinski, S.; Deplanque-Lasserre, J.; Hjorleifsson, E.; Geffen, A.J.; Godiksen, J.A.; Campana, S.E. Century-long cod otolith biochronology reveals individual growth plasticity in response to temperature. Sci. Rep. 2020, 10, 16708. [Google Scholar] [CrossRef]
- Smoliński, S.; Mirny, Z. Otolith biochronology as an indicator of marine fish responses to hydroclimatic conditions and ecosystem regime shifts. Ecol. Indic. 2017, 79, 286–294. [Google Scholar] [CrossRef]
- D’Iglio, C.; Albano, M.; Famulari, S.; Savoca, S.; Panarello, G.; Di Paola, D.; Perdichizzi, A.; Rinelli, P.; Lanteri, G.; Spano, N.; et al. Intra- and interspecific variability among congeneric Pagellus otoliths. Sci. Rep. 2021, 11, 16315. [Google Scholar] [CrossRef]
- Santos, L.; Vaz-dos-Santos, A.M. Insights of otoliths morphology to reveal patterns of Teleostean fishes in the southern Atlantic. Fishes 2022, 8, 21. [Google Scholar] [CrossRef]
- Lundberg, Y.W.; Xu, Y.; Thiessen, K.D.; Kramer, K.L. Mechanisms of otoconia and otolith development. Dev. Dyn. 2015, 244, 239–253. [Google Scholar] [CrossRef]
- Serre, S.H.; Nielsen, K.E.; Thomsen, T.B.; Hüssy, K. Analysis of cod otolith microchemistry by continuous line transects using LA-ICP-MS. Geol. Surv. Den. Greenl. Bull. 2018, 41, 91–94. [Google Scholar] [CrossRef]
- Rideout, R.M.; Youssef, T.N.; Adamack, A.T.; John, R.; Cohen, A.M.; Fridgen, T.D.; Banoub, J.H. Qualitative shotgun proteomics strategy for protein expression profiling of fish otoliths. BioChem 2023, 3, 102–117. [Google Scholar] [CrossRef]
- Campana, S.E. Chemistry and composition of fish otoliths:pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef]
- Thomas, O.R.B.B.; Swearer, S.E.; Kapp, E.A.; Peng, P.; Tonkin-Hill, G.Q.; Papenfuss, A.; Roberts, A.; Bernard, P.; Roberts, B.R.; Tonkin-Hill, G.Q.; et al. The inner ear proteome of fish. FEBS J. 2019, 286, 66–81. [Google Scholar] [CrossRef]
- Huang, S.; Qian, S. Advances in otolith-related protein research. Front. Neurosci. 2022, 16, 1–9. [Google Scholar]
- Bielak, K.; Benkowska-Biernacka, D.; Ptak, M.; Stolarski, J.; Kalka, M.; Ozyhar, A.; Dobryszycki, P. Otolin-1, an otolith- and otoconia-related protein, controls calcium carbonate bioinspired mineralization. Biochim. Biophys. Acta Gen. Subj. 2023, 1867, 130327. [Google Scholar] [CrossRef]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.C.; Yates, J.R., 3rd. Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 2013, 113, 2343–2394. [Google Scholar] [CrossRef]
- Wolters, D.A.; Washburn, M.P.; Yates, J.R. An automated multidimensional protein identification technology for shotgun proteomics. Anal. Chem. 2001, 73, 5683–5690. [Google Scholar] [CrossRef]
- Yates, J.R., 3rd. Mass spectral analysis in proteomics. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 297–316. [Google Scholar] [CrossRef]
- Link, A.J.; Eng, J.; Schieltz, D.M.; Carmack, E.; Mize, G.J.; Morris, D.R.; Garvik, B.M.; Yates, J.R., 3rd. Direct analysis of protein complexes using mass spectrometry. Nat. Biotechnol. 1999, 17, 676–682. [Google Scholar] [CrossRef]
- Yates, J.R., 3rd. Mass spectrometry and the age of the proteome. J. Mass. Spectrom. 1998, 33, 1–19. [Google Scholar] [CrossRef]
- Banoub, J.H.; Youssef, T.; Mikhael, A.; Mikhae, A. Proteomic technology applications for fisheries. Tech. Report. Fish. Aquat. Sci. 2022, 3465, xi + 668 p. [Google Scholar]
- Valikangas, T.; Suomi, T.; Elo, L.L. A systematic evaluation of normalization methods in quantitative label-free proteomics. Brief. Bioinform. 2018, 19, 1–11. [Google Scholar] [CrossRef]
- Wickham, H.; Henry, L. tidyr: Tidy Messy Data. R Package Version 1.1.3. 2021. Available online: https://CRAN.R-project.org/package=tidyr (accessed on 14 May 2024).
- Bache, S.M.; Wickham, H. magrittr: A Forward-Pipe Operator for R. R Package Version 2.0.1. 2021. Available online: https://CRAN.R-project.org/package=magrittr (accessed on 14 May 2024).
- Wickham, H. The Split-Apply-Combine Strategy for Data Analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation. R Package Version 1.1.4. 2023. Available online: https://github.com/tidyverse/dplyr (accessed on 14 May 2024).
- Hadley, W. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Tomanek, L. Variation in the heat shock response and its implication for predicting the effect of global climate change on species’ biogeographical distribution ranges and metabolic costs. J. Exp. Biol. 2010, 213, 971–979. [Google Scholar] [CrossRef]
- Litscher, E.S.; Wassarman, P.M. The fish egg’s zona pellucida. Curr. Top. Dev. Biol. 2018, 130, 275–305. [Google Scholar]
- Nathaniel Clarke, D.; Lowe, C.J.; James Nelson, W. The cadherin-catenin complex is necessary for cell adhesion and embryogenesis in Nematostella vectensis. Dev. Biol. 2019, 447, 170–181. [Google Scholar] [CrossRef]
- Hevrøy, E.M.; Jordal, A.E.O.; Hordvik, I.; Espe, M.; Hemre, G.I.; Olsvik, P.A. Myosin heavy chain mRNA expression correlates higher with muscle protein accretion than growth in Atlantic salmon, Salmo salar. Aquaculture 2006, 252, 453–461. [Google Scholar] [CrossRef]
- Casas, L.; Saborido-Rey, F.; Ryu, T.; Michell, C.; Ravasi, T.; Irigoien, X. Sex change in Clownfish: Molecular insights from transcriptome analysis. Sci. Rep. 2016, 6, 35461. [Google Scholar] [CrossRef]
- Cohen, A.M.; Mansour, A.A.H.; Banoub, J.H. ‘De novo’ sequencing of Atlantic cod vitellogenin tryptic peptides by matrix-assisted laser desorption/ionization quadrupole time-of-flight tandem mass spectrometry: Similarities with haddock vitellogenin. Rapid Commun. Mass. Spectrom. 2005, 19, 2454–2460. [Google Scholar] [CrossRef]
- Liu, W.B.; Yan, Q.; Liu, F.Y.; Tang, X.C.; Chen, H.G.; Liu, J.; Nie, L.; Zhang, X.W.; Ji, W.K.; Hu, X.H.; et al. Protein serine/threonine phosphotase-1 is essential in governing normal development of vertebrate eye. Curr. Mol. Med. 2012, 12, 1361–1371. [Google Scholar] [CrossRef]
- Schärer, O.D. XPG: Its Products and Biological Roles. In Molecular Mechanisms of Xeroderma Pigmentosum; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2009; pp. 83–92. [Google Scholar]
- Pancho, A.; Aerts, T.; Mitsogiannis, M.D.; Seuntjens, E. Protocadherins at the crossroad of signaling pathways. Front. Mol. Neurosci. 2020, 13, 117. [Google Scholar] [CrossRef]
- Pottmeier, P.; Nikolantonaki, D.; Lanner, F.; Peuckert, C.; Jazin, E. Sex-biased gene expression during neural differentiation of human embryonic stem cells. Front. Cell Dev. Biol. 2024, 12, 1341373. [Google Scholar] [CrossRef]
- Tonks, N.K. Protein tyrosine phosphatases: From genes, to function, to disease. Nat. Rev. Mol. Cell Biol. 2006, 7, 833–846. [Google Scholar] [CrossRef]
- Lenoir, M.; Kufareva, I.; Abagyan, R.; Overduin, M. Membrane and protein interactions of the Pleckstrin homology domain superfamily. Membr. 2015, 5, 646–663. [Google Scholar] [CrossRef]
- Staverosky, J.A.; Pryce, B.A.; Watson, S.S.; Schweitzer, R. Tubulin polymerization-promoting protein family member 3, Tppp3, is a specific marker of the differentiating tendon sheath and synovial joints. Dev. Dyn. 2009, 238, 685–692. [Google Scholar] [CrossRef]
- Orosz, F. On the tubulin polymerization promoting proteins of zebrafish. Biochem. Biophys. Res. Commun. 2015, 457, 267–272. [Google Scholar] [CrossRef]
- Noiges, R.; Eichinger, R.; Kutschera, W.; Fischer, I.; Nemeth, Z.; Wiche, G.; Propst, F. Microtubule-associated protein 1A (MAP1A) and MAP1B: Light chains determine distinct functional properties. J. Neurosci. 2002, 22, 2106–2114. [Google Scholar] [CrossRef]
- Sveinsdóttir, H.; Vilhelmsson, O.; Gudmundsdóttir, Á. Proteome analysis of abundant proteins in two age groups of early Atlantic cod (Gadus morhua) larvae. Comp. Biochem. Physiol. Part. D Genom. Proteom. 2008, 3, 243–250. [Google Scholar] [CrossRef]
- Focant, B.; Huriaux, F.; Baras, E.; Vandewalle, P. Expression of myofibrillar proteins and parvalbumin isoforms in white muscle of the developing turbot Scophthalmus maximus (Pisces, Pleuronectiformes). Basic. Appl. Myol. 2000, 10, 269–278. [Google Scholar]
- Huriaux, F.; Baras, E.; Vandewalle, P.; Focant, B. Expression of myofibrillar proteins and parvalbumin isoforms in white muscle of dorada during development. J. Fish. Biol. 2003, 62, 774–792. [Google Scholar] [CrossRef]
- Perry, S.V. Vertebrate tropomyosin: Distribution, properties and function. J. Muscle Res. Cell Motil. 2001, 22, 5–49. [Google Scholar] [CrossRef]
- Oehlmann, V.D.; Berger, S.; Sterner, C.; Korsching, S.I. Zebrafish beta tubulin 1 expression is limited to the nervous system throughout development, and in the adult brain is restricted to a subset of proliferative regions. Gene Expr. Patterns 2004, 4, 191–198. [Google Scholar]
- Meek, S.E.; Lane, W.S.; Piwnica-Worms, H. Comprehensive proteomic analysis of interphase and mitotic 14-3-3-binding proteins. J. Biol. Chem. 2004, 279, 32046–32054. [Google Scholar] [CrossRef]
- Van Hemert, M.J.; Steensma, H.Y.; van Heusden, G.P. 14-3-3 proteins: Key regulators of cell division, signalling and apoptosis. Bioessays 2001, 23, 936–946. [Google Scholar] [CrossRef]
- Suzuki, K.; Sato, K.; Katsu, K.; Hayashita, H.; Kristensen, D.B.; Yoshizato, K. Novel Rana keratin genes and their expression during larval to adult epidermal conversion in bullfrog tadpoles. Differentiation 2001, 68, 44–54. [Google Scholar] [CrossRef]
- Suzuki, K.; Utoh, R.; Kotani, K.; Obara, M.; Yoshizato, K. Lineage of anuran epidermal basal cells and their differentiation potential in relation to metamorphic skin remodeling. Dev. Growth Differ. 2002, 44, 225–238. [Google Scholar] [CrossRef]
- Ishida, Y.; Suzuki, K.; Utoh, R.; Obara, M.; Yoshizato, K. Molecular identification of the skin transformation center of anuran larval skin using genes of Rana adult keratin (RAK) and SPARC as probes. Dev. Growth Differ. 2003, 45, 515–526. [Google Scholar] [CrossRef]
- Jeyachandran, S.; Chellapandian, H.; Park, K.; Kwak, I.-S. A Review on the involvement of heat shock proteins (Extrinsic Chaperones) in response to stress conditions in aquatic organisms. Antioxidants 2023, 12, 1444. [Google Scholar] [CrossRef]
- Xie, Y.; Song, L.; Weng, Z.; Liu, S.; Liu, Z. Hsp90, Hsp60 and sHsp families of heat shock protein genes in channel catfish and their expression after bacterial infections. Fish. Shellfish. Immunol. 2015, 44, 642–651. [Google Scholar] [CrossRef]
- Song, L.; Li, C.; Xie, Y.; Liu, S.; Zhang, J.; Yao, J.; Jiang, C.; Li, Y.; Liu, Z. Genome-wide identification of Hsp70 genes in channel catfish and their regulated expression after bacterial infection. Fish. Shellfish. Immunol. 2016, 49, 154–162. [Google Scholar] [CrossRef]
- Chiosis, G.; Digwal, C.S.; Trepel, J.B.; Neckers, L. Structural and functional complexity of HSP90 in cellular homeostasis and disease. Nat. Rev. Mol. Cell Biol. 2023, 24, 797–815. [Google Scholar] [CrossRef]
- Didenko, T.; Duarte, A.M.; Karagoz, G.E.; Rudiger, S.G. Hsp90 structure and function studied by NMR spectroscopy. Biochim. Biophys. Acta 2012, 1823, 636–647. [Google Scholar] [CrossRef]
- Street, T.O.; Lavery, L.A.; Agard, D.A. Substrate binding drives large-scale conformational changes in the Hsp90 molecular chaperone. Mol. Cell 2011, 42, 96–105. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish. Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef]
- Duan, Y.; Tang, H.; Mitchell-Silbaugh, K.; Fang, X.; Han, Z.; Ouyang, K. Heat shock protein 60 in cardiovascular physiology and diseases. Front. Mol. Biosci. 2020, 7, 73. [Google Scholar] [CrossRef]
- Song, L.; Zhang, J.; Li, C.; Yao, J.; Jiang, C.; Li, Y.; Liu, S.; Liu, Z. Genome-wide identification of hsp40 genes in channel catfish and their regulated expression after bacterial infection. PLoS ONE 2014, 9, e115752. [Google Scholar] [CrossRef]
- Hori, T.S.; Gamperl, A.K.; Afonso, L.O.; Johnson, S.C.; Hubert, S.; Kimball, J.; Bowman, S.; Rise, M.L. Heat-shock responsive genes identified and validated in Atlantic cod (Gadus morhua) liver, head kidney and skeletal muscle using genomic techniques. BMC Genom. 2010, 11, 72. [Google Scholar] [CrossRef]
- Gollock, M.J.; Currie, S.; Petersen, L.H.; Gamperl, A.K. Cardiovascular and haematological responses of Atlantic cod (Gadus morhua) to acute temperature increase. J. Exp. Biol. 2006, 209, 2961–2970. [Google Scholar] [CrossRef]
- Perez-Casanova, J.C.; Rise, M.L.; Dixon, B.; Afonso, L.O.; Hall, J.R.; Johnson, S.C.; Gamperl, A.K. The immune and stress responses of Atlantic cod to long-term increases in water temperature. Fish. Shellfish. Immunol. 2008, 24, 600–609. [Google Scholar] [CrossRef]
- Kassahn, K.S.; Caley, M.J.; Ward, A.C.; Connolly, A.R.; Stone, G.; Crozier, R.H. Heterologous microarray experiments used to identify the early gene response to heat stress in a coral reef fish. Mol. Ecol. 2007, 16, 1749–1763. [Google Scholar] [CrossRef]
- Buckley, B.A.; Gracey, A.Y.; Somero, G.N. The cellular response to heat stress in the gobyGillichthys mirabilis: A cDNA microarray and protein-level analysis. J. Exp. Biol. 2006, 209, 2660–2677. [Google Scholar] [CrossRef]
- Podrabsky, J.E.; Somero, G.N. Changes in gene expression associated with acclimation to constant temperatures and fluctuating daily temperatures in an annual killifish Austrofundulus limnaeus. J. Exp. Biol. 2004, 207, 2237–2254. [Google Scholar] [CrossRef]
- Basu, N.; Todgham, A.E.; Ackerman, P.A.; Bibeau, M.R.; Nakano, K.; Schulte, P.M.; Iwama, G.K. Heat shock protein genes and their functional significance in fish. Gene 2002, 295, 173–183. [Google Scholar] [CrossRef]
- Yiangou, M.; Paraskeva, E.; Hsieh, C.C.; Markou, E.; Victoratos, P.; Scouras, Z.; Papaconstantinou, J. Induction of a subgroup of acute phase protein genes in mouse liver by hyperthermia. Biochim. Biophys. Acta 1998, 1396, 191–206. [Google Scholar] [CrossRef]
- Gross-Sorokin, M.Y.; Roast, S.D.; Brighty, G.C. Assessment of feminization of male fish in English rivers by the Environment Agency of England and Wales. Environ. Health Perspect. 2006, 114 (Suppl. S1), 147–151. [Google Scholar] [CrossRef]
- Mills, L.J.; Gutjahr-Gobell, R.E.; Horowitz, D.B.; Denslow, N.D.; Chow, M.C.; Zaroogian, G.E. Relationship between reproductive success and male plasma vitellogenin concentrations in cunner, Tautogolabrus adspersus. Env. Health Perspect. 2003, 111, 93–100. [Google Scholar] [CrossRef]
- Soffker, M.; Tyler, C.R. Endocrine disrupting chemicals and sexual behaviors in fish-a critical review on effects and possible consequences. Crit. Rev. Toxicol. 2012, 42, 653–668. [Google Scholar] [CrossRef]
- Montjean, D.; Neyroud, A.S.; Yefimova, M.G.; Benkhalifa, M.; Cabry, R.; Ravel, C. Impact of endocrine disruptors upon non-genetic inheritance. Int. J. Mol. Sci. 2022, 23, 3350. [Google Scholar] [CrossRef]
- Cohen, A.M.; Mansour, A.A.; Banoub, J.H. Absolute quantification of Atlantic salmon and rainbow trout vitellogenin by the ‘signature peptide’ approach using electrospray ionization QqToF tandem mass spectrometry. J. Mass. Spectrom. 2006, 41, 646–658. [Google Scholar] [CrossRef]
- Cohen, A.M.; Jahouh, F.; Sioud, S.; Rideout, R.M.; Morgan, M.J.; Banoub, J.H. Quantification of Greenland halibut serum vitellogenin: A trip from the deep sea to the mass spectrometer. Rapid Commun. Mass. Spectrom. 2009, 23, 1049–1060. [Google Scholar] [CrossRef]
- Banoub, J.; Thibault, P.; Mansour, A.; Cohen, A.; Heeley, D.H.; Jackman, D. Characterisation of the intact rainbow trout vitellogenin protein and analysis of its derived tryptic and cyanogen bromide peptides by matrix-assisted laser desorption/ionisation time-of-flight-mass spectrometry and electrospray ionisation quadrupole/time. Eur. J. Mass. Spectrom. 2003, 9, 509–524. [Google Scholar] [CrossRef]
- Cohen, A.M.; Banoub, J.H. Application of Mass Spectrometry for the Analysis of Vitellogenin, a Unique Biomarker for Xenobiotic Compounds. NATO Sci. Peace Secur. Ser. A: Chem. Biol. 2010, 301–318. [Google Scholar] [CrossRef]
- Katahira, J. Nuclear export of messenger RNA. Genes 2015, 6, 163–184. [Google Scholar] [CrossRef]
- Zhao, H.; Chiaro, C.R.; Zhang, L.; Smith, P.B.; Chan, C.Y.; Pedley, A.M.; Pugh, R.J.; French, J.B.; Patterson, A.D.; Benkovic, S.J. Quantitative analysis of purine nucleotides indicates that purinosomes increase de novo purine biosynthesis. J. Biol. Chem. 2015, 290, 6705–6713. [Google Scholar] [CrossRef]
- Tanaka, K. The proteasome: Overview of structure and functions. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 12–36. [Google Scholar] [CrossRef]
- Gregory, B.; Rahman, N.; Bommakanti, A.; Shamsuzzaman, M.; Thapa, M.; Lescure, A.; Zengel, J.M.; Lindahl, L. The small and large ribosomal subunits depend on each other for stability and accumulation. Life Sci. Alliance 2019, 2, e201800150. [Google Scholar] [CrossRef]
- Dreyfuss, G.; Philipson, L.; Mattaj, I.W. Ribonucleoprotein particles in cellular processes. J. Cell Biol. 1988, 106, 1419–1425. [Google Scholar] [CrossRef]
- Moreno, A.; Taleb, V.; Sebastian, M.; Anoz-Carbonell, E.; Martinez-Julvez, M.; Medina, M. Cofactors and pathogens: Flavin mononucleotide and flavin adenine dinucleotide (FAD) biosynthesis by the FAD synthase from Brucella ovis. IUBMB Life 2022, 74, 655–671. [Google Scholar] [CrossRef]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef]
- Moore, M.N.; Koehler, A.; Lowe, D.; Viarengo, A. Chapter thirty-three lysosomes and autophagy in aquatic animals. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2008; pp. 581–620. [Google Scholar]
- Bonam, S.R.; Wang, F.; Muller, S. Lysosomes as a therapeutic target. Nat. Rev. Drug Discov. 2019, 18, 923–948. [Google Scholar] [CrossRef]
- Hu, G.; Shu, Y.; Luan, P.; Zhang, T.; Chen, F.; Zheng, X. Genomic analysis of the proteasome subunit gene family and their response to high density and saline-alkali stresses in grass carp. Fishes 2022, 7, 350. [Google Scholar] [CrossRef]
- Sharkey, T.D. Pentose phosphate pathway reactions in photosynthesizing cells. Cells 2021, 10, 1547. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; Li, B.; Ding, L.; Wei, X.; Wang, P.; Chen, Z.; Han, S.; Huang, T.; Wang, B.; et al. Physiological responses to heat stress in the liver of rainbow trout (Oncorhynchus mykiss) revealed by UPLC-QTOF-MS metabolomics and biochemical assays. Ecotoxicol. Environ. Saf. 2022, 242, 113949. [Google Scholar] [CrossRef]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef]
- Wang, W.; Nag, S.; Zhang, X.; Wang, M.H.; Wang, H.; Zhou, J.; Zhang, R. Ribosomal proteins and human diseases: Pathogenesis, molecular mechanisms, and therapeutic implications. Med. Res. Rev. 2015, 35, 225–285. [Google Scholar] [CrossRef]
- Pattin, K.A.; Moore, J.H. Role for protein-protein interaction databases in human genetics. Expert. Rev. Proteom. 2009, 6, 647–659. [Google Scholar] [CrossRef]
- Wimalagunasekara, S.S.; Weeraman, J.; Tirimanne, S.; Fernando, P.C. Protein-protein interaction (PPI) network analysis reveals important hub proteins and sub-network modules for root development in rice (Oryza sativa). J. Genet. Eng. Biotechnol. 2023, 21, 69. [Google Scholar] [CrossRef]
- Cyr, F.; Snook, S.; Bishop, C.; Galbraith, P.S.; Pye, B.; Chen, N.; Han, G. Physical oceanographic conditions on the Newfoundland and Labrador shelf during 2019. DFO Can. Sci. Advis. Sec. Res. Doc. 2021, iv + 54. Available online: https://waves-vagues.dfo-mpo.gc.ca/library-bibliotheque/40960754.pdf (accessed on 14 May 2024).
- Cyr, F.; Colbourne, E.; Holden, J.; Snook, S.; Han, G.; Chen, N.; Bailey, W.; Higdon, J.; Lewis, S.; Pye, B.; et al. Physical oceanographic conditions on the Newfoundland and Labrador shelf during 2017. DFO Can. Sci. Advis. Sec. Res. 2019, iv + 58. Available online: https://waves-vagues.dfo-mpo.gc.ca/library-bibliotheque/40888113.pdf (accessed on 14 May 2024).
- Cyr, F.; Galbraith, P.S. A climate index for the Newfoundland and Labrador shelf. Earth Syst. Sci. Data 2021, 13, 1807–1828. [Google Scholar] [CrossRef]
- Alexander, M.A.; Shin, S.I.; Scott, J.D.; Curchitser, E.; Stock, C. The response of the Northwest Atlantic ocean to climate change. J. Clim. 2020, 33, 405–428. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Accession | Female Count a | Male Count b | Abundances (Average): Male c |
|---|---|---|---|---|
| Charged multivesicular body protein 2Ba OS = Gadus morhua OX = 8049 GN = chmp2ba PE = 3 SV = 1 | A0A8C4YUD6 | 0 | 5 | 261,448.36432 |
| DNA damage-binding protein 1 OS = Gadus morhua OX = 8049 GN = ddb1 PE = 3 SV = 1 | A0A8C5A9Z6 | 0 | 5 | 98,004.482948 |
| Fibroblast growth factor OS = Gadus morhua OX = 8049 PE = 3 SV = 1 | A0A8C4ZEA7 | 0 | 5 | 24,497.65508 |
| Microtubule-associated protein 1Ab OS = Gadus morhua OX = 8049 GN = map1ab PE = 4 SV = 1 | A0A8C5CU29 | 0 | 5 | 3,458,736.85 |
| Uncharacterized protein OS = Gadus morhua OX = 8049 PE = 3 SV = 1 | A0A8C5FA14 | 0 | 5 | 778,547.8001 |
| Description | Accession | Female Count a | Male Count b | Abundances (Average): Female c |
|---|---|---|---|---|
| Dihydrolipoamide acetyltransferase component of pyruvate dehydrogenase complex OS = Gadus morhua OX = 8049 GN = dbt PE = 3 SV = 1 | A0A8C5C1K4 | 5 | 0 | 108,232.639474 |
| Galectin OS = Gadus morhua OX = 8049 GN = LOC115545324 PE = 4 SV = 1 | A0A8C5ACJ8 | 5 | 0 | 489,972.337868 |
| Heterogeneous nuclear ribonucleoprotein A/Ba OS = Gadus morhua OX = 8049 GN = hnrnpaba PE = 4 SV = 1 | A0A8C4YVY0 | 5 | 0 | 1,301,640.954998 |
| Myosin light chain, phosphorylatable, fast skeletal muscle a OS = Gadus morhua OX = 8049 GN = mylpfa PE = 4 SV = 1 | A0A8C5B8E2 | 5 | 0 | 495,851.125408 |
| Nidogen 2a (osteonidogen) OS = Gadus morhua OX = 8049 PE = 4 SV = 1 | A0A8C5D3Y0 | 5 | 0 | 1,349,773.1182679997 |
| Polypeptide N-acetylgalactosaminyltransferase OS = Gadus morhua OX = 8049 GN = galnt6 PE = 3 SV = 1 | A0A8C5BP60 | 5 | 0 | 42,686.3808994 |
| SRSF protein kinase 2 OS = Gadus morhua OX = 8049 GN = srpk2 PE = 4 SV = 1 | A0A8C5CES3 | 5 | 0 | 20,084.879869999997 |
| Uncharacterized protein OS = Gadus morhua OX = 8049 PE = 4 SV = 1 | A0A8C5CCX7 | 5 | 0 | 211,271.979502 |
| Vinculin OS = Gadus morhua OX = 8049 PE = 3 SV = 1 | A0A8C5A430 | 5 | 0 | 411,121.875006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, T.N.; Christian, S.L.; Rideout, R.; Adamack, A.; Thibault, P.; Bonneil, E.; Fridgen, T.D.; Banoub, J. Proteomic Blueprint of Atlantic Cod (Gadus morhua) Otoliths Revealing Environmental Stress Insights through Label-Free Quantitative Shotgun Proteomics. BioChem 2024, 4, 144-165. https://doi.org/10.3390/biochem4020008
Youssef TN, Christian SL, Rideout R, Adamack A, Thibault P, Bonneil E, Fridgen TD, Banoub J. Proteomic Blueprint of Atlantic Cod (Gadus morhua) Otoliths Revealing Environmental Stress Insights through Label-Free Quantitative Shotgun Proteomics. BioChem. 2024; 4(2):144-165. https://doi.org/10.3390/biochem4020008
Chicago/Turabian StyleYoussef, Trevena N., Sherri L. Christian, Rick Rideout, Aaron Adamack, Pierre Thibault, Eric Bonneil, Travis D. Fridgen, and Joseph Banoub. 2024. "Proteomic Blueprint of Atlantic Cod (Gadus morhua) Otoliths Revealing Environmental Stress Insights through Label-Free Quantitative Shotgun Proteomics" BioChem 4, no. 2: 144-165. https://doi.org/10.3390/biochem4020008
APA StyleYoussef, T. N., Christian, S. L., Rideout, R., Adamack, A., Thibault, P., Bonneil, E., Fridgen, T. D., & Banoub, J. (2024). Proteomic Blueprint of Atlantic Cod (Gadus morhua) Otoliths Revealing Environmental Stress Insights through Label-Free Quantitative Shotgun Proteomics. BioChem, 4(2), 144-165. https://doi.org/10.3390/biochem4020008