
Role of microRNA in Oxidative Stress


Department of Biotechnology, Institute of Applied Sciences & Humanities, GLA University, Mathura 281406, Uttar Pradesh, India
Stresses 2024, 4(2), 269-281; https://doi.org/10.3390/stresses4020016
Submission received: 1 March 2024
/
Revised: 29 March 2024
/
Accepted: 7 April 2024
/
Published: 9 April 2024
(This article belongs to the Section Animal and Human Stresses)
{kind=link}
{kind=link}
Abstract
:An imbalance between the formation of reactive oxygen species (ROS) and the reaction of antioxidant proteins is referred to as oxidative stress. NFE2L2/Nrf2, also known as nuclear factor erythroid-derived 2-related factor 2, is a critical enabler of cytoprotective responses to oxidative and electrophilic insults. When Nrf2 is activated, it triggers the transcription of numerous cytoprotective genes, whose promoter regions contain antioxidant response elements (AREs). In recent times, the regulation of Nrf2 by miRNAs has garnered significant attention, among the various mechanisms that govern Nrf2 signaling. It has been reported that a number of miRNAs directly suppress the expression of Nrf2s, which in turn negatively regulates the Nrf2-dependent cellular cytoprotective response. Furthermore, it has been shown that Nrf2 itself regulates miRs, which carry out some of Nrf2’s unique metabolic regulation functions. Here, we provide an overview of the functions and mechanisms of action of miRs as downstream effectors of Nrf2, as well as in their regulation of its activity.
1. Introduction
An imbalance between the formation of reactive oxygen species (ROS) and the reaction of antioxidant proteins is referred to as oxidative stress. Cells rely on a range of biochemical, metabolic, and genetic pathways to sustain this balance between oxidants and antioxidants; when it is disrupted, pathophysiological consequences may ensue [1]. Reactive oxygen (ROS), reactive nitrogen (RNS), and reactive sulfur (RSS) species are thought to be triggers for a variety of pathogenic illnesses, such as cancer, inflammatory disorders, cardiovascular disease, neurological diseases, and persistent infections [2]. Reactive oxygen species (ROS) play essential roles in living organisms and are mainly produced under physiological conditions. A moderate rise in ROS can usually promote cell differentiation and proliferation, even though an excess of ROS can cause oxidative damage to proteins, lipids, and DNA [3]. The bulk of clinical trials employing general antioxidant therapy have failed, despite tremendous efforts over the past few decades to bring antioxidant therapy into clinical practice. This is most likely due to a lack of knowledge regarding the roles of redox signaling pathways in health and disease [4]. With a greater understanding of the mechanisms through which oxidants work, as well as the potential drawbacks and benefits of antioxidant therapy, a rational plan for improving therapeutic intervention may be developed.
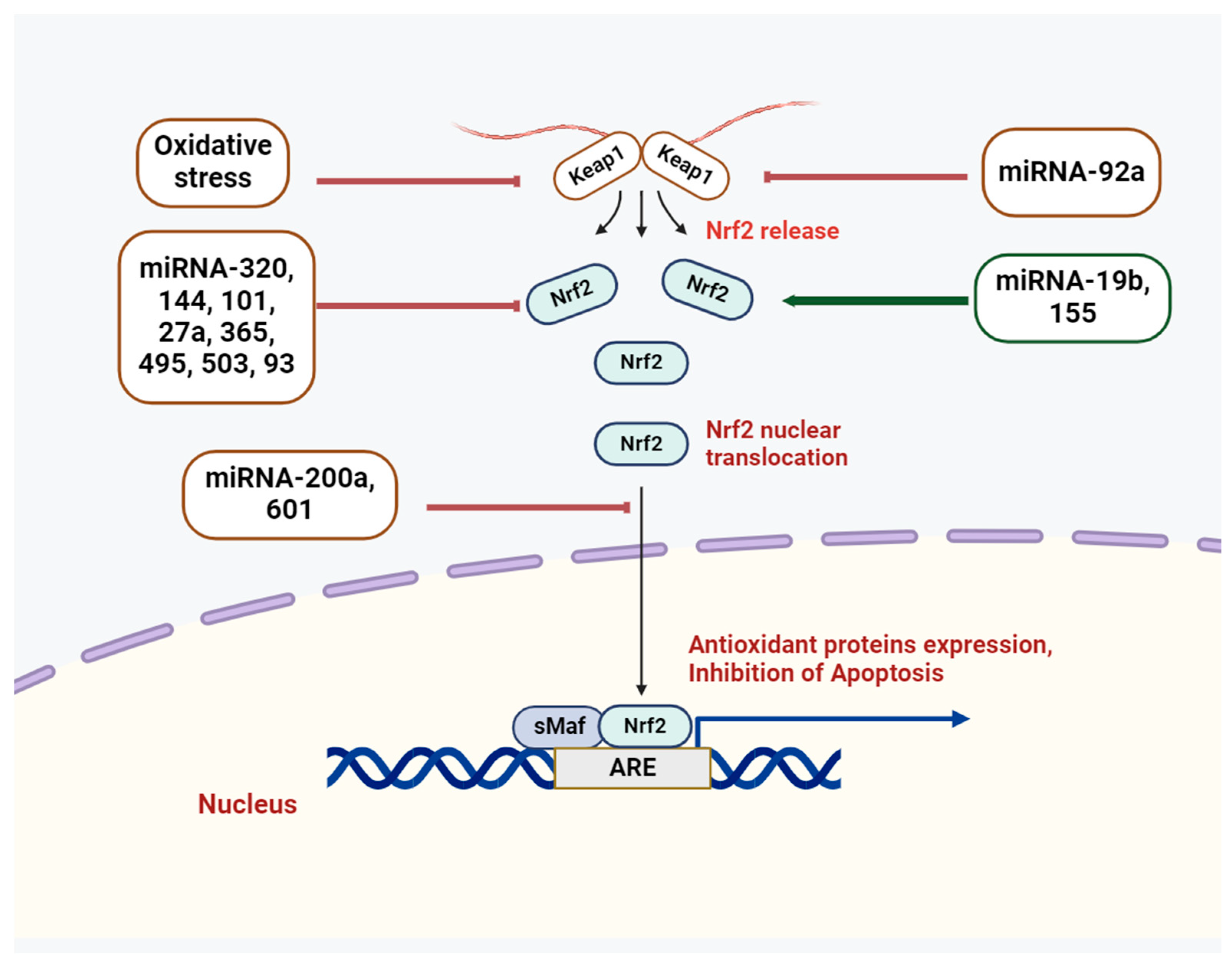
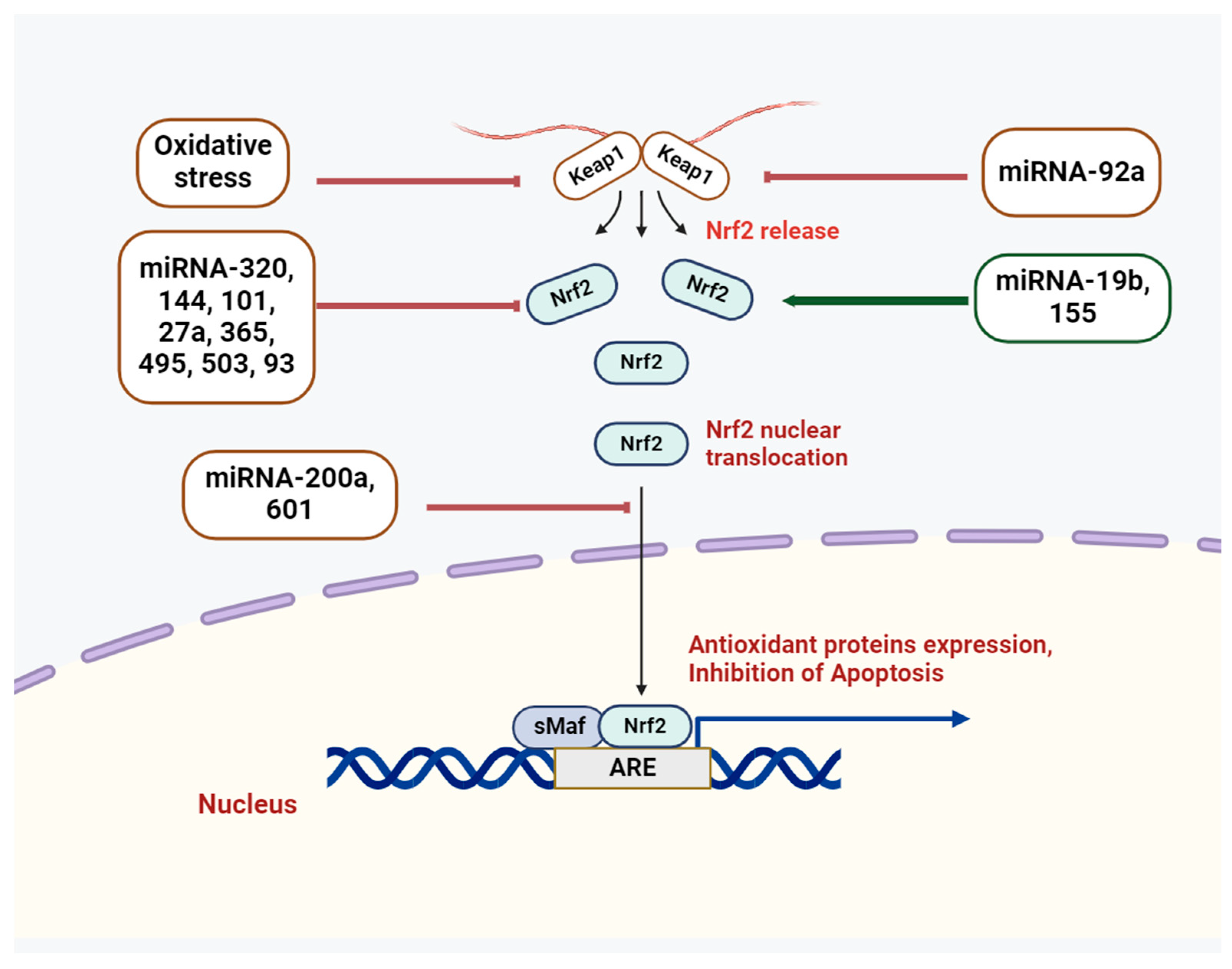
In this context, the Nrf2/Keap1 signaling systems have demonstrated critical roles among the major antioxidant systems [5]. Both Nrf2 and its antagonistic regulator, the E3 ligase adaptor Kelch-like ECH-associated protein 1 (Keap1), are critical to sustaining the redox state and the synthesis of antioxidant genes [6]. Normally, Keap1 attaches to the Neh2 domain of Nrf2 through the ETGE and DLG motifs, which results in the cytoplasmic localization of Nrf2 [7]. Nevertheless, oxidative stress causes a conformational change in the E3-ligase complex that stops Nrf2 from interacting with the ubiquitin-conjugating mechanism [7]. The outcome is the release of Nrf2 from the complex and its entry into the nucleus, where it joins forces with the sMaf protein to form a heterodimer. Subsequently, the ARE is recognized by Nrf2, further regulating the synthesis of antioxidant proteins and cell defense systems [7]. Genes like TXN, G6PD, GSTA2, NQO1, and HO1 (HMOX1), which are involved in redox detoxification and NADH regeneration, are stimulated to express themselves by the Nrf2 pathway [8]. Nrf2 signaling is connected to a wide range of oxidative stress-related illnesses, including pulmonary disorders, cardiovascular abnormalities, and neurodegenerative diseases [9,10,11]. Because of its important functions, Nrf2’s activity needs to be tightly controlled, and its targets in different tissues and organs need to be known.
On the other hand, growing evidence from earlier research suggests that ROS accumulation changed the expression of microRNA [12]. MicroRNAs are 19–25 nt long noncoding RNA molecules that are crucial for posttranscriptional regulation because they bind to the 3′-untranslated regions (UTRs) of target genes and control a number of biological processes, including cell division, apoptosis, and stress response [13]. Previous research has revealed that the regulation of microRNAs varies depending on the type of cell and tissue. While a single mRNA may be targeted by multiple microRNAs, multiple microRNAs may also influence a single mRNA. As a result, different groups of mRNAs involved in a variety of pathological conditions and the pathogenesis of different human diseases, including immune system disorders and cancers, can be regulated by microRNAs [14]. However, due to its relative stability, microRNA may be able to serve as both a novel biomarker for diagnosis and a possible target for treatment for a variety of cancers [15]. Understanding the interaction between ROS production and microRNAs is crucial because it has been linked to the development of cancer and several other pathological conditions. Remarkably, OS controls the expression levels of certain miRNAs, known as ROS-miRs or redoximiRs, which change target gene expression in response to ROS [16]. In MELAS cells, the OS/NF-κB axis induced the expression of miR-9/9*, while inhibiting the expression of GTPBP3, MTO1, and TRMU [17]. However, in human umbilical vein endothelial cells (HUVECs), a prior study found that miR-21 regulated ROS homeostasis and inhibited the antioxidant response [18]. These results imply that ROS may function as miRNAs’ downstream effectors or upstream regulators. This review focuses on the ways in which the crosslink between miRNAs and the Nrf2 signaling pathway regulate ROS-mediated function and how ROS influence biological phenotypes through this interconnection.
As outlined in the following paragraphs, recent research has demonstrated the significant roles that miRNAs (miRs) play in both the regulation of Nrf2 and its downstream effects, in addition to a variety of mRNA targets. The RNA-induced silencing complex (RISC) is brought in to either cleave or repress the translation of the targeted mRNAs when miRs bind correspondingly to their mRNA targets, which are most frequently found within the 3′-UTR [13]. Numerous miRs may target a single mRNA because miRs regulate a large number of targets in a combinatorial manner [19]. Furthermore, oxidative stress triggers the expression of miR-200 in a variety of cells and tissues. In addition to targeting chemotherapy resistance, members of the miR-200 family may regulate the homeostasis of reactive oxygen species during oxidative stress [20].
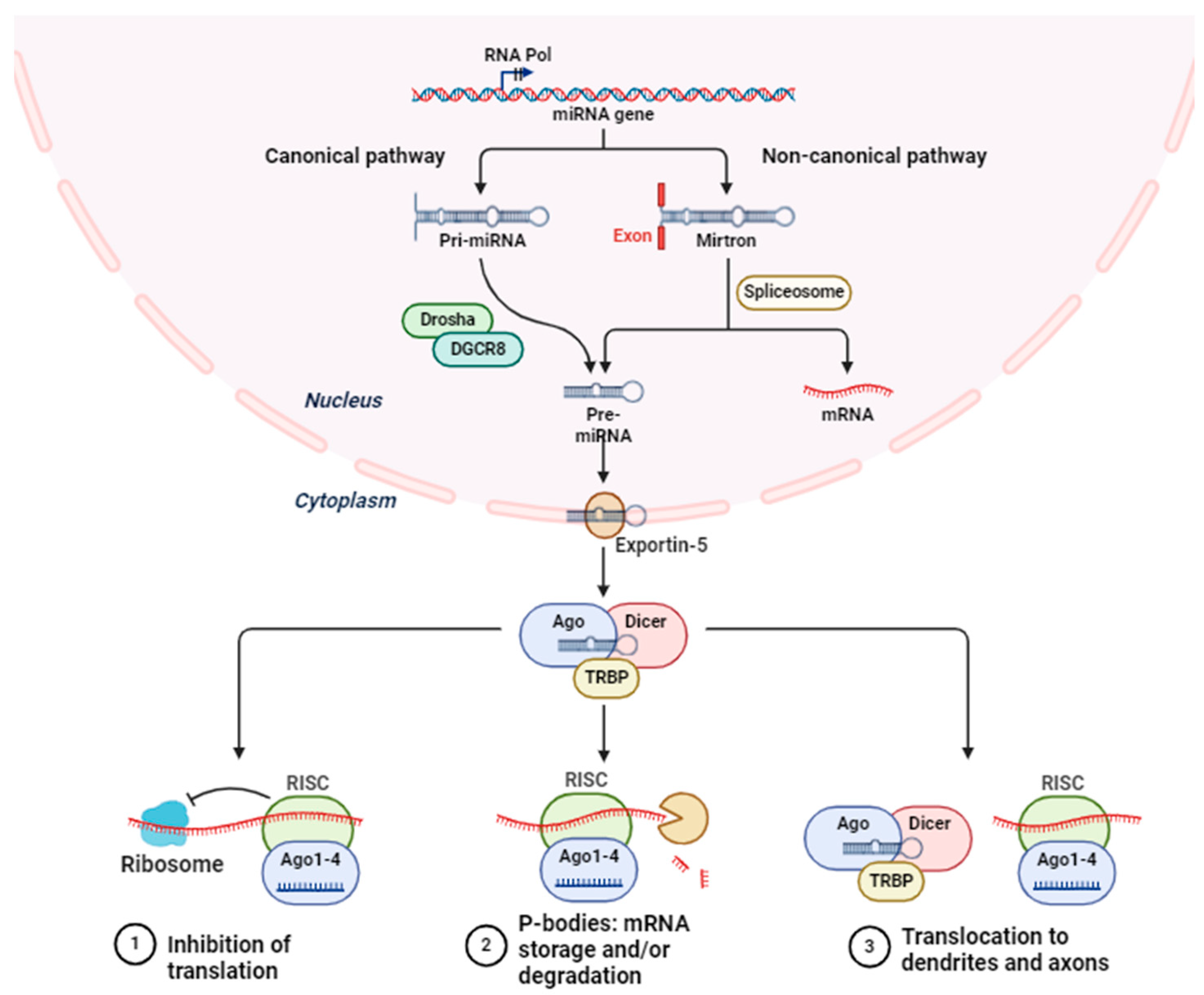
2. miRNA Biology: Biogenesis and Action
Short regulatory RNAs, known as microRNAs (miRNAs), function as post-transcriptional repressors of gene expression in a variety of biological contexts. Molecular biology has undergone a revolutionary shift since the Ambros and Ruvkun groups discovered the first microRNA (miRNA), lin-4, in Caenorhabditis elegans in 1993 [21]. All animal model systems contain miRNAs, some of which have been found to be highly conserved across species [22,23,24]. The roles that miRNAs play in regulating gene expression are widely known, and new ones are constantly being found [25]. Small non-coding RNAs, or miRNAs, have an average length of 22 nucleotides. The majority of miRNAs are produced by converting DNA sequences into primary miRNAs, or pri-miRNAs, which are then processed into mature miRNAs and precursor miRNAs, or pre-miRNAs. miRNAs typically suppress expression by interacting with the 3′ UTR of target RNAs [26]. Nevertheless, there have also been reports of miRNA interaction with other regions, such as the 5′ UTR, coding sequence, and gene promoters [27]. Recent studies have suggested that miRNAs are shuttled between different subcellular compartments to control the rate of translation, and even transcription [28].
miRNA biogenesis is initiated by post- or co-transcriptionally processing RNA polymerase II/III transcripts (Figure 1) [26]. Approximately 50% of all miRNAs that have been discovered thus far are intragenic, meaning they are primarily processed from introns and a small number of exons of genes that code for proteins. The other 50% are intergenic, meaning they are transcribed without the help of a host gene and are controlled by their own promoters [29]. When miRNAs are transcribed as clusters, they are regarded as a family because they may share seed regions and are, therefore, considered to be a single long transcript [30]. Under the canonical pathway, pri-miRNAs are generated from their genes and subsequently processed into premiRNAs by the microprocessor complex, which is made up of the ribonuclease III enzyme Drosha and the RNA binding protein DiGeorge Syndrome Critical Region 8 (DGCR8) [31]. Although Drosha cleaves the pri-miRNA duplex at the base of the pri-miRNA’s distinctive hairpin structure, DGCR8 recognizes an N6-methyladenylated GGAC motif [28]. As a consequence, pre-miRNA develops a 2 nt 3′ overhang [32]. An exportin 5 (XPO5)/RanGTP complex transports them to the cytoplasm, where they are subsequently degraded by the RNase III endonuclease Dicer [33]. By removing the terminal loop during this processing, a mature miRNA duplex is produced [34]. The mature miRNA form’s name is determined by the directionality of the miRNA strand. The pre-miRNA hairpin’s 5′ end gives rise to the 5p strand, while the 3′ end is where the 3p strand begins. It is possible to load both strands from the mature miRNA duplex into the ATP-dependent Argonaute (AGO) family of proteins [35].
On the other hand, numerous non-canonical miRNA biogenesis pathways have been identified thus far. These pathways primarily use combinations of Drosha, Dicer, exportin 5, and AGO2, which are proteins involved in the canonical pathway. Non-canonical miRNA biogenesis can be broadly classified into pathways that are either Dicer-independent or Drosha/DGCR8-independent. Pre-miRNAs produced by the Drosha/DGCR8-independent pathway resemble Dicer substrates. Without requiring Drosha cleavage, exportin 1 transports these developing RNAs straight to the cytoplasm. The m7G cap’s ability to inhibit 5p strand loading into Argonaute is most likely the cause of the strong 3p strand bias [36]. Conversely, Drosha processes endogenous short hairpin RNA (shRNA) transcripts to produce Dicer-independent miRNAs [37]. Because these pre-miRNAs are too short to be Dicer substrates, they need AGO2 to finish maturing within the cytoplasm [38] Consequently, this encourages the full pre-miRNA to be loaded into AGO2 and AGO2-dependent 3p strand slicing. Their maturation is finished by the 5p strand’s 3′–5′ trimming [38].
3. Cross-Talk of miRNAs and Nrf2
Raising the level of ROS influences the expression of miRs, which alters the cellular profile of miR content [39]. By regulating their target genes, such notable alterations in the cellular miR profile have subsequent biological effects [40]. In order to alter the cellular redox balance and potentially create an environment that supports the growth and metastasis of cancer cells, miRs may target genes that produce reactive oxygen species (ROS), or elements that are responsive to antioxidants. Consequently, there may be a reciprocal relationship between ROS signaling and miR pathways, which could have a variety of biological effects.
To put it briefly, miRs have the ability to modulate the Nrf2 signaling pathway upstream and have a high degree of control over the pathway at different stages. Overall, miRs control the Nrf2 pathway through the following four major mechanisms: (a) altering Nrf2’s nuclear translocation; (b) affecting Nrf2’s expression; (c) controlling the Nrf2 upstream mediators; and (d) modulating Keap1.
Although Nrf2 is known to act as a transcription factor for traditional cytoprotective genes, there is little research on its potential to regulate miRNA transcripts. In order to identify Nrf2 target transcripts, including miRNAs, one important study used chromatin immunoprecipitation sequencing (ChIP-Seq) on lymphoid cells treated with the Nrf2 activator, sulforaphane [41]. Near nine putative miRNAs, high confidence ChIP-Seq peaks were found, suggesting that Nrf2 regulates these miRNAs. Only miR-365, miR-193b, miR-181c, and miR-29b were found to be expressed in the lymphoblast cell line, according to conformational studies, and only miR-29b demonstrated a significant change in expression in response to Nrf2 activation. Through its direct control of HDAC4 expression, Nrf2 can indirectly downregulate miR-1 and miR-206, according to a different recent study [42]. Nrf2 binding increased the expression levels of miR-29a and miR-29b in primary human keratinocytes and the mouse epidermis [43]. Transgenic mice expressing a dominant negative Nrf2 mutant in keratinocytes—a mutant that retains DNA binding ability but lacks the transactivation domain—did not exhibit upregulation of miR-29a/b1 expression. This result indicates that the activation of the Mir29ab1 gene requires Nrf2’s transcriptional activity. Treatment with HDACi boosted Nrf2 transcriptional activity and raised Nrf2 mRNA and protein levels [44]. On the other hand, autophagy induced by HDACi was inhibited or blocked by Nrf2 knockdown. Furthermore, an array of microRNAs (miRNAs) revealed that Nrf2 overexpression was associated with the upregulation of miR-129-3p. miR-129-3p was shown to be a direct target of Nrf2 by chromatin immunoprecipitation technologies.
Mutant expression of the miR-365/193b cluster has been associated with cancers, including those of the colon and lungs. It is interesting to note that miR-365 and miR-193b can both be linked to tumors independently of one another in the cluster. Reduced expression of thyroid transcription factor 1, critical for lung development and frequently upregulated in lung tumors, has been linked to downregulation of miR-365 in lung cancer [45]. On the other hand, it is believed that over-expression of miR-365 in colon cancer inhibits the development and growth of tumors by focusing on the anti-apoptotic genes BCL-2 and cyclin D1. Nrf2 has the ability to control the miR-365/193b cluster, according to ChIP-Seq data [41]. Front-line chemotherapy drugs such as cytarbine/daunorubicin, bortezomib (myeloma), and 5-fluorouracil (colon cancer) cause Nrf2 to become upregulated. It was found that miR-200a targets keap1 mRNA and, thus, releases Nrf2 [46]. This leads to the induction of cytoprotective genes, which, in turn, reduces the cell’s sensitivity to these drugs [46].
miR-181c is primarily linked to the differentiation of hematopoietic cells and the pathogenesis of cancer. It has been demonstrated that miR-181c represses a number of genes linked to cancer and is highly expressed in the thymus, brain, liver, and bone marrow [47]. It seems that miR-181c has a tumor-type-specific role in cancer. Most gastric cancer samples tested had downregulated miR-181c, but a small percentage had significantly higher expression, suggesting that this miR can function as both a tumor suppressor and an oncomiR [48].
In human colorectal adenocarcinoma HT29 cells, silencing Nrf2 reduced ATP levels, which, in turn, triggered AMPK activation [49]. The regulation of AMPK signaling was linked to increased miR-181c in Nrf2-knockdown cancer cells as the underlying mechanism. AMP-activated protein kinase-α (AMPKα) and acetyl CoA carboxylase were phosphorylated more when a miR-181c mimic was introduced into control cells. On the other hand, AMPKα and ACC phosphorylation were decreased in Nrf2-knockdown cancer cells upon transfection of a miR-181c inhibitor. Interestingly, these modifications in metabolic signaling caused Nrf2-silenced cancer cells to become more sensitive [49]. Another study found that Nrf2 silencing increased the expression of miR-181c via NF-κB signaling [50]. This increase was linked to a decrease in the catalytic core subunit of complex IV of the electron transport chain, mitochondria-encoded cytochrome c oxidase subunit-1. Interestingly, these modifications triggered the activation of AMPKα, and consequent signaling for metabolic adaptation, which included the inhibition of enzymes involved in the biosynthesis of sterol and fatty acids. Thus, the aforementioned study indicates the importance of Nrf2 in cancer mitochondria by clarifying the role of underlying molecular events related to miR-181c/MT-CO1 [50].
The ability of miR-200c to inhibit the epithelial to mesenchymal transition (EMT) makes it a tumor suppressor miR that is implicated in several cancers [51,52]. Additionally, miR-200c has been connected to drug resistance in a number of cancers. For example, because paclitaxel targets the microtubule component TUBB3, breast and ovarian cancer cells with higher expression levels of miR-200c are more susceptible to paclitaxel [52]. Research has also revealed that breast cancer cells that express low levels of miR-200c are more resistant to radiation, whereas overexpression of this miRNA results in a higher rate of apoptosis [53]. The fact that miR-200c targets TBK1, which, in cancers, represses apoptosis and activates other oncogenic pathways like AKT, contributes to this increased sensitivity. On the other hand, overexpression of miR-200c has been associated with chemoresistance and poor prognosis in esophageal cancers. This is because miR-200c targets the AKT phosphorylation repressor PPP2R1B, increasing AKT activation, which indicates their tissue-specific roles [54].
A recent study revealed apoptosis of enterocytes, colonic dysfunction, downregulation of miR-200a, inflammatory response, and inactivation of the Nrf2/Keap1 pathway in a colitis model (Figure 2) [55]. In addition, in dextran sulfate sodium-induced enterocytes, there was an increase in Keap1 expression and a decrease in the protein expression levels of Nrf2 and NQO1, which were restored by miR-200a. Following miR-200a overexpression, the mRNA expression levels of HO-1, NQO-1, and SOD were also restored, indicating that miR-200a mitigates these detrimental changes by activating the Nrf2 antioxidant pathway and repressing Keap1. As demonstrated by luciferase assays, miR-200a was able to target Keap1’s 3′-UTR. Thus, overexpression of miR-200a may inhibit intestinal inflammation and oxidative stress by degrading Keap1 and activating Nrf2.
Another study found that miR-200a can target Keap1 mRNA, encouraging Keap1 degradation and activating Nrf2, which protects mice against the cardiotoxic effects of doxorubicin [56]. Furthermore, by focusing on Keap1/Nrf2, miR-200a can ameliorate diabetic endothelial dysfunction [57]. Furthermore, polydatin-induced enhancement of miR-200a to regulate the Keap1/Nrf2 pathway can avert fructose-induced hepatic inflammation [58]. Another study revealed that doxorubicin significantly increased the expression level of miR-128-3p, decreased the expression level of Sirt1, and changed the levels of Nrf2 protein [59]. On the other hand, miR-128-3p antagomir reduced liver damage by increasing Sirt1, Nrf2, Sirt3, NQO1, and HO-1 protein levels. In another report, miR-140-5p targeted Nrf2 and Sirt2 to promote myocardial oxidative stress, which is a key factor in doxorubicin-induced cardiotoxicity. By altering the expression levels of HO-1, NQO1, Gst, GCLM, Keap1, and FOXO3a, miR-140-5p directly targeted Nrf2 and Sirt2 [60]. Furthermore, miR-140-5p/Nrf2 and miR-140-5p/Sirt2 might be new targets for treating cardiotoxicity brought on by doxorubicin.
In renal mesangial cells, aldose reductase has the ability to negatively regulate the expression of miR-200a-3p/141-3p [61]. Under high-glucose conditions, miR-200a-3p/141-3p directly targets Keap1, Tgfβ2, fibronectin, and Zeb2, and it indirectly regulates Tgfβ1 and Nrf2. Significant upregulation of Nrf2 and reduced Keap1 expression were observed in the renal cortex upon aldose reductase deficiency-induced elevations in miR-200a-3p/141-3p.
Furthermore, pertinent research indicates that a novel way to activate Nrf2 is through miRNA targeting and Keap1 silencing [62,63]. Through Keap1 targeting and silencing, to trigger Nrf2 signaling, miR-152 protects the heart [64]. By focusing on the Keap1-Nrf2-HO-1 axis, miR-141 suppresses the migration and proliferation of vascular smooth muscle cells [65]. In order to shield retinal pigment epithelial cells from oxidative damage, miR-626 activated Nrf2 and targeted Keap1 [66].
Sangokoya et al. found that erythrocyte samples from patients with homozygous sickle cell disease (HbSS) who were experiencing worsened anemia had elevated expression of miR-144 [67]. Luciferase experiments verified that miR-144 has a direct regulatory effect on Nrf2 expression, since the latter carries two target sequences for miR-144 on the 3′-UTR for Nrf2. A similar investigation demonstrated the direct regulatory impact of miR-28 on Nrf2 expression and the ensuing cellular functions in breast cancer cell lines [68]. Through binding on the Nrf2 3′-UTR, miR-28 successfully regulated Nrf2 expression, as demonstrated by luciferase reporter assays. It was also made clear, however, that the regulatory function of miR-28 is independent of Keap-1 because overexpression of miR-28 neither altered Keap-1 and Nrf2 interactions, nor caused dysregulated expression of Keap-1. Another study demonstrated a decrease in Nrf2 protein levels after miR-93 was induced to express itself, as well as a decrease in phenotypes linked to carcinogenesis after miR-93-knockdown experiments [69]. The miR-93 “seed” sequence did not perfectly match the 3′-UTR of Nrf2, but it still reduced the levels of Nrf2 mRNA and protein in breast cancer cells [70]. Nrf2 protein levels were reduced as a result of ectopic expression of miR-93. One of the known direct regulators of Nrf2 in breast cancer cells is miR-28, which has been shown to bind the Nrf2 3′-UTR and control Nrf2 expression without the need for KEAP1 [71]. Therefore, one potential tactic to restore Nrf2 functions could be to counteract the repression of this transcription factor, mediated by miR-28 and miR-93, in order to rescue Nrf2 activity.
In the SH-SY5Y neuroblastoma cell line, one study found that miR-153, miR-27-a, miR-142-5p, and miR-144 are regulatory miRNAs for Nrf2 [72]. These regulatory connections were confirmed by individual miRNA mimic co-transfection experiments, as well as by the observation of decreased Nrf2 transcription and protein levels after the miRNAs were expressed ectopically. When hydrogen peroxide was applied to neuroblastoma cells, miR-153 bound to the Nrf2 3′-UTR and inhibited the translation of a luciferase reporter [72]. miR-153 and miR-144 were significantly upregulated in rat hippocampi following traumatic brain injury, resulting in diminished mRNA and protein levels of Nrf2 [73]. Remarkably, earlier research had discovered Nrf2-bound enhancer/promoter elements upstream of gene clusters encoding for miR-29s [43], suggesting that Nrf2-dependent regulation of miR-29s in the brain may exist. Reduced expression levels of microsomal glutathione S-transferase 1 (Mgst1) and sirtuin 1 (Sirt1) are the result of miR-34a and miR-93 targeting Nrf2 in aged rat liver [74]. Nrf2 target gene expression was downregulated and Nrf2 mRNA and protein levels were markedly reduced by overexpression of miR-34a. More significantly, the Nrf2-dependent antioxidant pathway was restored and the effects of ischemia–reperfusion-induced hepatic injury were lessened when miR-34a expression was inhibited [75]. In another report, TNF-α treatment causes all endothelial cell lines to express less miR-27b, activates inflammatory pathways, changes the mitochondria, and produces an increase in reactive oxygen species, all of which aid in the induction of intrinsic apoptosis [76]. By targeting the 3′untranslated regions of FOXO1 mRNA, hsa-miR-27b-3p reduces the expression of this gene and lessens the activation of the Akt/FOXO1 pathway.
Following Nrf2 knockdown in acute myeloid leukemia cells, the results of miRNA microarray screenings demonstrated Nrf2-directed upregulated expression of miR-125-b1 and concurrent decrease in miR-29-b1 [77]. The research conducted by Joo and colleagues [78] provided more proof that Nrf2 controls the transcription of miR-125b1 expression, demonstrating how the activation of Nrf2 in the kidney tissues of mice given oltipraz increased the expression of miR-125-b1 [78]. In hepatocellular carcinoma (HCC) chemoresistance models, Shi and colleagues found an increase in miR-141 expression that correlated with drug resistance to 5-fluorouracil [79]. Using luciferase reporter assays and RT-qPCR for additional analysis, it was discovered that miR-141 directly regulated downstream Keap1 transcription [79]. The direct regulatory effect of miR-340 on Nrf2 within HCC cell lines exhibiting a chemoresistance phenotype for cisplatin was also demonstrated in another study [80]. Two miRNAs, miR-1 and miR-206, were downregulated in tandem with a decrease in cell line growth in A549 tumor cell lines that showed knockdown of Nrf2 [81]. Numerous studies demonstrated that the miR-103/107 family (which includes miR-103.1, miR-103.2, and miR-107) targets Dicer by directly binding to its 3′-UTR, thus attenuating miRNA biosynthesis [82].
The cells of the retinal pigment epithelium are vulnerable to oxidative stress. Increased nuclear translocation of Nrf2 and inhibition of Cul3 are caused by increased expression of miR-601 [83]. Consequently, elevated expression levels of ARE-dependent genes, like HO-1 and NQO1, strengthen the antioxidant defense system. Hepatotoxicity is caused by acetaminophen (APAP) overdose through control of the miR-19b [84]. Research has demonstrated that the administration of APAP causes miR-19b to be downregulated. This stage is essential for SIRT1 stimulation, which sets off the Nrf2 cascade and a number of downstream mediators, including antioxidant enzymes. Following I/R injury, a decrease in miR-320 expression has been observed [85]. Activation and expression of the Nrf2 signaling pathway and its downstream mediator, HO-1, are markedly accelerated by downregulating miR-320. I/R injury was attenuated as a result of the reduction in ROS concentrations brought about by Nrf2 stimulation. High oxidative stress sensitivity and cell pyroptosis are linked to chlorpyrifos exposure. Chlorpyrifos partially mediates these deleterious effects on SH-SY5Y cells by upregulating miR-181 [86]. Consequently, the SIRT1/PGC-1α/Nrf2 signaling pathway, which is implicated in cell protection and the inhibition of ROS generation, is suppressed by miR-181.
PRXL2A, a protein that may resemble the scavenger DJ-1/PARK7, is regulated by miR-125b [77]. By favorably influencing the Nrf2 signaling pathway, the upregulation of PRXL2A by miR-125b prevents oxidative damage in cells. I/R injury can affect cardiomyocytes [87]. Molecular pathways involved in the antioxidant defense system can be targeted to lessen this vulnerability [88]. Cardiomyocytes are protected from I/R damage by miR-24-3p, which efficiently initiates the Nrf2 signaling pathway by inhibiting Keap1 [88]. After ischemic stroke, attempts were made to lessen the cerebral I/R injury by miR-34b [89]. The results demonstrated that, while miR-34b increases the activities of SOD and manganese SOD (MnSOD), it significantly reduces the infarction volume, neurological severity scores, and the level of nitric oxide (NO). It was discovered that the inhibition of Keap1 mediates the protective effects of miR-34b. This inhibition of Keap1 then triggers the Nrf2 signaling pathway, which, in turn, activates the antioxidant enzymes. It has been demonstrated that, by blocking the Nrf2/ARE pathway, miR-101 suppresses cell division and promotes apoptosis in gastric mucosal epithelial cells. By attaching itself to the UTR of Nrf2, miR-101 inhibits the activity of Nrf2 [90].
NRAL, a long non-coding RNA, has been linked to cisplatin resistance, according to a 2019 study by Wu and colleagues [91]. The relationship between NRAL and miR-340-5p was also inverse, with NRAL having a negative effect on miR-340-5p to initiate Nrf2-dependent antioxidant enzymes. This indicates that the NRAL/miR-340-5p/Nrf2 axis plays a crucial role in hepatocellular carcinoma cells’ resistance to cisplatin [91]. It seems that the Nrf2 signaling pathway is downregulated by miR-495 activity during epilepsy and seizures, which results in apoptosis. Research has demonstrated that UCA1, a long noncoding RNA, inhibits the expression of miR-495, which, in turn, stimulates the Nrf2 pathway and prevents apoptosis in neurons and epileptiform hippocampal tissues [92].
Oxidative stress has a detrimental effect on bone metabolism, and miR-455-3p has the ability to modulate oxidative damage [93]. Apoptosis and a high degree of oxidative stress are linked to exposure to ferric ammonium citrate (FAC) and a reduction in cell proliferation [94]. The detrimental effects of FAC were counteracted by increasing the expression of miR-455-3p, which led to the restoration of bone metabolism and the inhibition of apoptosis [93]. Oxidative damage is reduced when miR-455-3p stimulates Nrf2’s activation through its upstream mediator, HDAC2 [93]. H2O2 has the ability to increase ROS and oxidative stress. Research has demonstrated the connection between exposure to H2O2 and the development of osteoporosis and osteonecrosis [95]. By degrading cul3, which, in turn, activates ARE-related genes, like NQO1, HO-1, and GCLC, miR-455 stabilizes and increases the level of Nrf2. By strengthening the antioxidant defense system, osteoblasts are shielded from the damaging effects of H2O2 [96].
As a tumor suppressor, miR-148b creates an environment in which ROS levels are high, which inhibits the growth of tumor cells [97]. The expression of endoplasmic reticulum metalloprotease 1 (ERMP1) is inhibited by miR-148b. Consequently, human endometrial cancer RL95-2 cells experience a reduction in cell proliferation due to the downregulation of Nrf2, which raises ROS levels [97]. Another recent study showed that miR-21 directed attenuating SOD3, or via an indirect mechanism that limited TNFα production, thereby reducing SOD2 levels, to inhibit the metabolism of superoxide to hydrogen peroxide, which is produced either by endogenous basal activities or exposure to ionizing radiation (IR) [98]. Therefore, their results demonstrate that miR-21 regulates cellular ROS levels, which, in turn, significantly promotes tumorigenesis. Reduction in ROS and oxidative stress-induced cell death were also observed upon overexpression of miR-210 [99]. Furthermore, following hypoxia–reoxygenation, downregulation of miR-210 increased ROS. Based on microRNA expression profiling, it was found that HIF-1α upregulates the levels of miR-210 and downregulates those of miR-383 [99].
4. Conclusions
Essentially, the purpose of this review is to draw attention to the clinical significance of miRNA effects within the Nrf2/KEAP1 pathway, as well as other important pathways that are impacted by Nrf2 activity. It is crucial to understand these novel molecular interactions at the miRnomic level, as dysregulated miRNAs that are thought to be responsible for exacerbating particular disease conditions with oxidative stress and clinical progression may be novel targets for medication. It was determined that elevated expression levels of the mir-103/107 family during the metastasis process will cause Dicer, mir-200b, and mir-200c to be downregulated or targeted, which will, in turn, promote the expression of sirt1 (the repressor of Nrf2). As a result, Nrf2’s expression level, and the downstream-regulated antioxidant-protective genes it regulates, will be reduced. By affecting Keap1, controlling Nrf2 expression, changing Nrf2’s nuclear translocation, and affecting upstream mediators of the Nrf2 pathway, like SIRT1/PGC1a, PRXL2A, Notch, BRMP1, HDAC2, and MEG3, miRs can influence the Nrf2 signaling pathway. It may be extremely beneficial for clinical practice to learn more about the extent of redox-sensitive miRNA-induced epigenetic gene regulatory control on crucial components of the Nrf2 signaling pathway and its interactions, especially in the context of oxidative stress, since, once discovered and verified, these miRNAs may be used as novel therapeutic targets for soon-to-be-released translational medicine-based treatments in clinical settings. This review article, as a whole, affirms and supports the significance of applying microRNA-based therapies in clinical settings following research trials.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Profumo, E.; Tucci, P.; Saso, L. A Perspective on Nrf2 Signaling Pathway for Neuroinflammation: A Potential Therapeutic Target in Alzheimer’s and Parkinson’s Diseases. Front. Cell. Neurosci. 2022, 15, 787258. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Saha, S.; Buttari, B.; Profumo, E.; Saso, L. Activation of Nrf2 signaling pathway by natural and synthetic chalcones: A therapeutic road map for oxidative stress. Expert Rev. Clin. Pharmacol. 2021, 14, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A. The Keap1-Nrf2pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Pinho, S.A.; Afonso, G.J.M.; Oliveira, P.J.; Cunha-Oliveira, T.; Saso, L. Nrf2 and Mitochondrial Function in Cancer and Cancer Stem Cells. Cells 2022, 11, 2401. [Google Scholar] [CrossRef] [PubMed]
- Boutten, A.; Goven, D.; Artaud-Macari, E.; Bonay, M. Protective role of Nrf2 in the lungs against oxidative airway diseases. Med. Sci. 2011, 27, 966–972. [Google Scholar]
- Gan, L.; Johnson, J.A. Oxidative damage and the Nrf2-ARE pathway in neurodegenerative diseases. Biochim. Biophys. Acta 2014, 1842, 1208–1218. [Google Scholar] [CrossRef]
- Howden, R. Nrf2 and cardiovascular defense. Oxidative Med. Cell. Longev. 2013, 2013, 104308. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jiang, B.H. Interplay between reactive oxygen species and microRNAs in cancer. Curr. Pharmacol. Rep. 2016, 2, 82–90. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Wilczynska, A.; Bushell, M. The complexity of miRNA mediated repression. Cell Death Diff. 2015, 22, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.A.; Arora, S.; Prakasam, G.; Calin, G.A.; Syed, M.A. MicroRNA in lung cancer: Role, mechanisms, pathways and therapeutic relevance. Mol. Asp. Med. 2019, 70, 3–20. [Google Scholar] [CrossRef]
- Liu, Y.; Qiang, W.; Xu, X.; Dong, R.; Karst, A.M.; Liu, Z.; Kong, B.; Drapkin, R.I.; Wei, J.J. Role of miR-182 in response to oxidative stress in the cell fate of human fallopian tube epithelial cells. Oncotarget 2015, 6, 38983–38998. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, S.; Martinez-Zamora, A.; Garcia-Arumi, E.; Andreu, A.L.; Armengod, M.E. The ROS-sensitive microRNA-9/9* controls the expression of mitochondrial tRNA-modifying enzymes and is involved in the molecular mechanism of MELAS syndrome. Hum. Mol. Genet. 2015, 24, 167–184. [Google Scholar] [CrossRef] [PubMed]
- La Sala, L.; Mrakic-Sposta, S.; Micheloni, S.; Prattichizzo, F.; Ceriello, A. Glucose-sensing microRNA-21 disrupts ROS homeostasis and impairs antioxidant responses in cellular glucose variability. Cardiovasc. Diabetol. 2018, 17, 105. [Google Scholar] [CrossRef]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Kozak, J.; Jonak, K.; Maciejewski, R. The function of miR-200 family in oxidative stress response evoked in cancer chemotherapy and radiotherapy. Biomed. Pharmacother. 2020, 125, 110037. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Davis-Dusenbery, B.N.; Hata, A. Mechanisms of control of microRNA biogenesis. J. Biochem. 2010, 148, 381–392. [Google Scholar]
- Li, S.C.; Chan, W.C.; Hu, L.Y.; Lai, C.H.; Hsu, C.N.; Lin, W.C. Identification of homologous microRNAs in 56 animal genomes. Genomics 2010, 96, 1–9. [Google Scholar] [CrossRef]
- Friedlander, M.R.; Lizano, E.; Houben, A.J.; Bezdan, D.; Banez-Coronel, M.; Kudla, G.; Mateu-Huertas, E.; Kagerbauer, B.; González, J.; Chen, K.C.; et al. Evidence for the biogenesis of more than 1000 novel human microRNAs. Genome Biol. 2014, 15, R57. [Google Scholar] [CrossRef] [PubMed]
- de Rie, D.; Abugessaisa, I.; Alam, T.; Arner, E.; Arner, P.; Ashoor, H.; Åström, G.; Babina, M.; Bertin, N.; Burroughs, A.M.; et al. An integrated expression atlas of miRNAs and their promoters in human and mouse. Nat. Biotechnol. 2017, 35, 872–878. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell 2016, 64, 320–333. [Google Scholar] [CrossRef]
- Makarova, J.A.; Shkurnikov, M.U.; Wicklein, D.; Lange, T.; Samatov, T.R.; Turchinovich, A.A.; Tonevitsky, A.G. Intracellular and extracellular microRNA: An update on localization and biological role. Prog. Histochem. Cytochem. 2016, 51, 33–49. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kim, V.N. Processing of intronic microRNAs. EMBO J. 2007, 26, 775–783. [Google Scholar] [CrossRef]
- Tanzer, A.; Stadler, P.F. Molecular evolution of a microRNA cluster. J. Mol. Biol. 2004, 339, 327–335. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.; Plasterk, R.H.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kolb, F.A.; Jaskiewicz, L.; Westhof, E.; Filipowicz, W. Single processing center models for human Dicer and bacterial RNase III. Cell 2004, 118, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Yoda, M.; Kawamata, T.; Paroo, Z.; Ye, X.; Iwasaki, S.; Liu, Q.; Tomari, Y. ATP-dependent human RISC assembly pathways. Nat. Struct. Mol. Biol. 2010, 17, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Li, M.; Vilborg, A.; Lee, N.; Shu, M.D.; Yartseva, V.; Šestan, N.; Steitz, J.A. Mammalian 5′-capped microRNA precursors that generate a single microRNA. Cell 2013, 155, 1568–1580. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Maurin, T.; Robine, N.; Rasmussen, K.D.; Jeffrey, K.L.; Chandwani, R.; Papapetrou, E.P.; Sadelain, M.; O’Carroll, D.; Lai, E.C. Conserved vertebrate mir-451 provides a platform for Dicer-independent, Ago2-mediated microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 15163–15168. [Google Scholar] [CrossRef] [PubMed]
- Cheloufi, S.; Dos Santos, C.O.; Chong, M.M.W.; Hannon, G.J. A dicer-independent miRNA biogenesis pathway that requires Ago catalysis. Nature 2010, 465, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Liu, X.; Cheng, Y.; Yang, J.; Huo, Y.; Zhang, C. Involvement of MicroRNAs in hydrogen peroxide-mediated gene regulation and cellular injury response in vascular smooth muscle cells. J. Biol. Chem. 2009, 284, 7903–7913. [Google Scholar] [CrossRef]
- Seo, J.; Jin, D.; Choi, C.H.; Lee, H. Integration of MicroRNA, mRNA, and protein expression data for the identification of cancer-related MicroRNAs. PLoS ONE 2017, 12, e0168412. [Google Scholar] [CrossRef]
- Chorley, B.N.; Campbell, M.R.; Wang, X.; Karaca, M.; Sambandan, D.; Bangura, F.; Xue, P.; Pi, J.; Kleeberger, S.R.; Bell, D.A. Identification of novel NRF2-regulated genes by ChIP-Seq: Influence on retinoid X receptor alpha. Nucleic Acids Res. 2012, 40, 7416–7429. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Happel, C.; Manna, S.K.; Acquaah-Mensah, G.; Carrerero, J.; Kumar, S.; Nasipuri, P.; Krausz, K.W.; Wakabayashi, N.; Dewi, R.; et al. Transcription factor NRF2 regulates miR-1 and miR-206 to drive tumorigenesis. J. Clin. Investig. 2013, 123, 2921–2934. [Google Scholar] [CrossRef] [PubMed]
- Kurinna, S.; Schafer, M.; Ostano, P.; Karouzakis, E.; Chiorino, G.; Bloch, W.; Bachmann, A.; Gay, S.; Garrod, D.; Lefort, K.; et al. A novel Nrf2-miR-29-desmocollin-2 axis regulates desmosome function in keratinocytes. Nat. Commun. 2014, 5, 5099. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yi, Y.; Xia, G.; Zhao, Y.; Yu, Y.; Li, L.; Hua, C.; He, B.; Yang, B.; Yu, C.; et al. Nrf2-miR-129-3p-mTOR Axis Controls an miRNA Regulatory Network Involved in HDACi-Induced Autophagy. Mol. Ther. 2019, 27, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Rice, S.J.; Salzberg, A.C.; Runkle, E.A.; Liao, J.; Zander, D.S.; Mu, D. MiR-365 regulates lung cancer and developmental gene thyroid transcription factor 1. Cell Cycle 2012, 11, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Zimta, A.A.; Cenariu, D.; Irimie, A.; Magdo, L.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. The role of Nrf2 activity in cancer development and progression. Cancers 2019, 11, 1755. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Akiyama, Y.; Otsubo, T.; Shimada, S.; Yuasa, Y. Involvement of epigenetically silenced microRNA-181c in gastric carcinogenesis. Carcinogenesis 2010, 31, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Kyeong-Ah, J.; Dayoung, R.; Geon, K.; Mi-Kyoung, K. 281—Role of NRF2 in MiR-181c/AMP-Activated Protein Kinase Signaling in Cancer. Free Radic. Biol. Med. 2016, 100, S124. [Google Scholar]
- Jung, K.A.; Lee, S.; Kwak, M.K. NFE2L2/NRF2 Activity Is Linked to Mitochondria and AMP-Activated Protein Kinase Signaling in Cancers through miR-181c/Mitochondria-Encoded Cytochrome c Oxidase Regulation. Antioxid. Redox Signal. 2017, 27, 945–961. [Google Scholar] [CrossRef]
- Chen, M.L.; Liang, L.S.; Wang, X.K. miR-200c inhibits invasion and migration in human colon cancer cells SW480/620 by targeting ZEB1. Clin. Exp. Metastasis 2012, 29, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ohuchida, K.; Mizumoto, K.; Sato, N.; Kayashima, T.; Fujita, H.; Nakata, K.; Tanaka, M. MicroRNA, hsa-miR-200c, is an independent prognostic factor in pancreatic cancer and its upregulation inhibits pancreatic cancer invasion but increases cell proliferation. Mol. Cancer 2010, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Liu, C.; Gao, F.; Mitchel, R.E.; Zhao, L.; Yang, Y.; Lei, J.; Cai, J. MiR-200c enhances radiosensitivity of human breast cancer cells. J. Cell. Biochem. 2012, 114, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Hamano, R.; Miyata, H.; Yamasaki, M.; Kurokawa, Y.; Hara, J.; ho Moon, J.; Nakajima, K.; Takiguchi, S.; Fujiwara, Y.; Mori, M.; et al. Overexpression of miR-200c induces chemoresistance in esophageal cancers mediated through activation of the Akt signaling pathway. Clin. Cancer Res. 2011, 17, 3029–3038. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Shen, L.; Yu, X.; Wu, J.; Zha, L.; Xia, Y.; Luo, H. miR-200a attenuated oxidative stress, inflammation, and apoptosis in dextran sulfate sodium-induced colitis through activation of Nrf2. Front. Immunol. 2023, 14, 1196065. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, H.; Wang, Z.; Hu, Z.; Li, L. miR-200a attenuated doxorubicin-induced cardiotoxicity through upregulation of Nrf2 in mice. Oxidative Med. Cell. Longev. 2019, 2019, 1512326. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Wu, J.; Ma, F.; Jiang, J.; Xu, L.; Du, L.; Huang, W.; Wang, Z.; Jia, Y.; Lu, L.; et al. MicroRNA-200a improves diabetic endothelial dysfunction by targeting KEAP1/NRF2. J. Endocrinol. 2020, 24, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.J.; Yu, H.W.; Yang, Y.Z.; Wu, W.Y.; Chen, T.Y.; Jia, K.K.; Kang, L.-L.; Jiao, R.-Q.; Kong, L.-D. Polydatin prevents fructose-induced liver inflammation and lipid deposition through increasing miR-200a to regulate Keap1/Nrf2 pathway. Redox Biol. 2018, 18, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jin, Y.; Li, L.; Xu, L.; Tang, Z.; Qi, Y.; Yin, L.; Peng, J. MicroRNA-128-3p aggravates doxorubicin-induced liver injury by promoting oxidative stress via targeting Sirtuin-1. Pharmacol. Res. 2019, 146, 104276. [Google Scholar] [CrossRef]
- Zhao, L.; Qi, Y.; Xu, L.; Tao, X.; Han, X.; Yin, L.; Peng, J. MicroRNA-140-5p aggravates doxorubicin-induced cardiotoxicity by promoting myocardial oxidative stress via targeting Nrf2 and Sirt2. Redox Biol. 2018, 15, 284–296. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Luo, Y.; Wang, Z.; Bi, S.; Song, D.; Dai, Y.; Wang, T.; Qiu, L.; Wen, L.; et al. Aldose reductase regulates miR-200a-3p/141-3p to coordinate Keap1-Nrf2, Tgfβ1/2, and Zeb1/2 signaling in renal mesangial cells and the renal cortex of diabetic mice. Free Radic. Biol. Med. 2014, 67, 91–102. [Google Scholar] [CrossRef]
- Akdemir, B.; Nakajima, Y.; Inazawa, J.; Inoue, J. miR-432 induces NRF2 stabilization by directly targeting KEAP1. Mol. Cancer Res. 2017, 15, 1570–1578. [Google Scholar] [CrossRef]
- Silva-Palacios, A.; Ostolga-Chavarria, M.; Zazueta, C.; Konigsberg, M. Nrf2: Molecular and epigenetic regulation during aging. Ageing Res. Rev. 2018, 47, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.B.; Lai, X.; Guo, X.F. Activation of Nrf2 by miR-152 Inhibits Doxorubicin-Induced Cardiotoxicity via Attenuation of Oxidative Stress, Inflammation, and Apoptosis. Oxidative Med. Cell. Longev. 2021, 2021, 8860883. [Google Scholar] [CrossRef]
- Zhang, C.; Kong, X.; Ma, D. miR-141-3p inhibits vascular smooth muscle cell proliferation and migration via regulating Keap1/Nrf2/HO-1 pathway. IUBMB Life 2020, 72, 2167–2179. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Z.; Tang, Y.; Cheng, L.B.; Yao, J.; Jiang, Q.; Li, K.R.; Zhen, Y.-F. Targeting Keap1 by miR-626 protects retinal pigment epithelium cells from oxidative injury by activating Nrf2 signaling. Free Radic. Biol. Med. 2019, 143, 387–396. [Google Scholar] [CrossRef]
- Sangokoya, C.; Telen, M.J.; Chi, J.-T. microRNA miR-144 modulates oxidative stress tolerance and associates with anemia severity in sickle cell disease. Blood 2010, 116, 4338–4348. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yao, Y.; Eades, G.; Zhang, Y.; Zhou, Q. MiR-28 regulates Nrf2 expression through a Keap1-independent mechanism. Breast Cancer Res. Treat. 2011, 129, 983–991. [Google Scholar] [CrossRef]
- Singh, B.; Ronghe, A.M.; Chatterjee, A.; Bhat, N.K.; Bhat, H.K. MicroRNA-93 regulates NRF2 expression and is associated with breast carcinogenesis. Carcinogenesis 2013, 34, 1165–1172. [Google Scholar] [CrossRef]
- Hussen, B.M.; Abdullah, S.R.; Rasul, M.F.; Jawhar, Z.H.; Faraj, G.S.H.; Kiani, A.; Taheri, M. MiRNA-93: A novel signature in human disorders and drug resistance. Cell Commun. Signal. 2023, 21, 79. [Google Scholar] [CrossRef]
- Zimmerman, A.L.; Wu, S. MicroRNAs, cancer and cancer stem cells. Cancer Lett. 2011, 300, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, M.; Riar, A.K.; Rathinam, M.L.; Vedpathak, D.; Henderson, G.; Mahimainathan, L. Hydrogen peroxide responsive miR153 targets Nrf2/ARE cytoprotection in paraquat induced dopaminergic neurotoxicity. Toxicol. Lett. 2014, 228, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sun, T.; Liu, Z.; Chen, X.; Zhao, L.; Qu, G.; Li, Q. Traumatic brain injury dysregulates microRNAs to modulate cell signaling in rat hippocampus. PLoS ONE 2014, 9, e103948. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Muthusamy, S.; Liang, R.; Sarojini, H.; Wang, E. Increased expression of miR-34a and miR-93 in rat liver during aging, and their impact on the expression of Mgst1 and Sirt1. Mech. Ageing Dev. 2011, 132, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, Y.; Qin, J.; Lu, S. The role of miR-34a in the hepatoprotective effect of hydrogen sulfide on ischemia/reperfusion injury in young and old rats. PLoS ONE 2014, 9, e113305. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, N.; Prattichizzo, F.; Martino, E.; Anastasio, C.; Mele, L.; La Grotta, R.; Sardu, C.; Ceriello, A.; Marfella, R.; Paolisso, G.; et al. MiR-27b attenuates mitochondrial oxidative stress and inflammation in endothelial cells. Redox Biol. 2023, 62, 102681. [Google Scholar] [CrossRef]
- Shah, N.M.; Zaitseva, L.; Bowles, K.M.; MacEwan, D.J.; Rushworth, S.A. NRF2-driven miR-125B1 and miR-29B1 transcriptional regulation controls a novel anti-apoptotic miRNA regulatory network for AML survival. Cell Death Differ. 2015, 22, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Joo, M.S.; Lee, C.G.; Koo, J.H.; Kim, S.G. miR-125b transcriptionally increased by Nrf2 inhibits AhR repressor, which protects kidney from cisplatin-induced injury. Cell Death Dis. 2013, 4, e899. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wu, L.; Chen, Z.; Yang, J.; Chen, X.; Yu, F.; Zheng, F.; Lin, X. MiR-141 activates Nrf2-dependent antioxidant pathway via down-regulating the expression of keap1 conferring the resistance of hepatocellular carcinoma cells to 5-fluorouracil. Cell. Physiol. Biochem. 2015, 35, 2333–2348. [Google Scholar] [CrossRef]
- Shi, L.; Chen, Z.-G.; Wu, L.-L.; Zheng, J.-J.; Yang, J.-R.; Chen, X.-F.; Chen, Z.-Q.; Liu, C.-L.; Chi, S.-Y.; Zheng, J.-Y.; et al. miR-340 reverses cisplatin resistance of hepatocellular carcinoma cell lines by targeting Nrf2-dependent antioxidant pathway. Asian Pac. J. Cancer Prev. 2014, 15, 10439–10444. [Google Scholar] [CrossRef]
- Solich, J.; Kolasa, M.; Faron-Górecka, A.; Pabian, P.; Latocha, K.; Korlatowicz, A.; Dziedzicka-Wasylewska, M. Modulating Stress Susceptibility and Resilience: Insights from miRNA Manipulation and Neural Mechanisms in Mice. Mol. Neurobiol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Hsin-Yi, C.; Yu-Min, L.; Hsiang-Ching, C.; Yaw-Dong, L.; Ching-Jung, L.; John, H.; Wei-Chi, W.; Feng-Mao, L.; Zhen, C.; Hsien-Da, H.; et al. miR-103/107 promote metastasis of colorectal cancer by targeting the metastasis suppressors DAPK and KLF4. Canc. Res. 2012, 72, 3631–3641. [Google Scholar]
- Wang, X.; Yuan, B.; Cheng, B.; Liu, Y.; Zhang, B.; Wang, X.; Lin, X.; Yang, B.; Gong, G. Crocin alleviates myocardial ischemia/reperfusion-induced endoplasmic reticulum stress via regulation of miR-34a/Sirt1/Nrf2 pathway. Shock 2019, 51, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, H.; Luo, C.; Du, D.; Huang, J.; Ming, Q.; Jin, F.; Wang, D.; Huang, W. Acetaminophen responsive miR-19b modulates SIRT1/Nrf2 signaling pathway in drug-induced hepatotoxicity. Toxicol. Sci. 2019, 170, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.A.; Gao, L.F.; Zhang, Z.G.; Xiang, D.K. Down-regulation of miR-320 exerts protective effects on myocardial I-R injury via facilitating Nrf2 expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1730–1741. [Google Scholar] [PubMed]
- Cookson, M.R. Parkinson’s Disease, Disease-Modifying Targets in Neurodegenerative Disorders; Elsevier: Amsterdam, The Netherlands, 2017; pp. 157–174. [Google Scholar]
- Liao, Y.H.; Xia, N.; Zhou, S.F.; Tang, T.T.; Yan, X.X.; Lv, B.J.; Nie, S.-F.; Wang, J.; Iwakura, Y.; Xiao, H.; et al. Interleukin-17A contributes to myocardial ischemia/reperfusion injury by regulating cardiomyocyte apoptosis and neutrophil infiltration. J. Am. Coll. Cardiol. 2012, 59, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Lu, Z.; Lin, V.; May, A.; Shaw, D.H.; Wang, Z.; Che, B.; Tran, K.; Du, H.; Shaw, P.X. MicroRNA miR-24-3p reduces apoptosis and regulates Keap1-Nrf2 pathway in mouse cardiomyocytes responding to ischemia/reperfusion injury. Oxidative Med. Cell. Longev. 2018, 2018, 7042105. [Google Scholar] [CrossRef]
- Huang, R.; Ma, J.; Niu, B.; Li, J.; Chang, J.; Zhang, Y.; Liu, P.; Luan, X. MiR-34b protects against focal cerebral ischemia-reperfusion (I/R) injury in rat by targeting Keap1. J. Stroke Cerebrovasc. Dis. 2019, 28, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Q.; Zhang, Y.H.; Shang, X.Q.; Zeng, Y.J. Effects of miR-101 on the proliferation and apoptosis of gastric mucosal epithelial cells via Nrf2/ARE signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5187–5194. [Google Scholar]
- Wu, L.L.; Cai, W.P.; Lei, X.; Shi, K.Q.; Lin, X.Y.; Shi, L. NRAL mediates cisplatin resistance in hepatocellular carcinoma via miR-340-5p/Nrf2 axis. J. Cell Commun. Signal. 2019, 13, 99–112. [Google Scholar] [CrossRef]
- Geng, J.F.; Liu, X.; Zhao, H.B.; Fan, W.F.; Geng, J.J.; Liu, X.Z. LncRNA UCA1 inhibits epilepsy and seizure-induced brain injury by regulating miR-495/Nrf2-ARE signal pathway. Int. J. Biochem. Cell Biol. 2018, 99, 133–139. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, W.; Jiao, G.; Li, C.; Liu, H. MiR-455-3p activates Nrf2/ARE signaling via HDAC2 and protects osteoblasts from oxidative stress. Int. J. Biol. Macromol. 2018, 107, 2094–2101. [Google Scholar] [CrossRef]
- Kang, J.S.; Cho, D.; Kim, Y.I.; Hahm, E.; Kim, Y.S.; Jin, S.N.; Na Kim, H.; Kim, D.; Hur, D.; Park, H.; et al. Sodium ascorbate (vitamin C) induces apoptosis in melanoma cells via the down-regulation of transferrin receptor dependent iron uptake. J. Cell. Physiol. 2005, 204, 192–197. [Google Scholar] [CrossRef]
- Talasila, K.M.; Soentgerath, A.; Euskirchen, P.; Rosland, G.V.; Wang, J.; Huszthy, P.C.; Prestegarden, L.; Skaftnesmo, K.O.; Sakariassen, P.; Eskilsson, E.; et al. EGFR wild-type amplification and activation promote invasion and development of glioblastoma independent of angiogenesis. Acta Neuropathol. 2013, 125, 683–698. [Google Scholar] [CrossRef]
- Xu, D.; Zhu, H.; Wang, C.; Zhu, X.; Liu, G.; Chen, C.; Cui, Z. microRNA-455 targets cullin 3 to activate Nrf2 signaling and protect human osteoblasts from hydrogen peroxide. Oncotarget 2017, 8, 59225. [Google Scholar] [CrossRef]
- Qu, J.; Zhang, L.; Li, L.; Su, Y. miR-148b functions as a tumor suppressor by targeting endoplasmic reticulum metallo protease 1 in human endometrial cancer cells. Oncol. Res. 2018, 27, 81–88. [Google Scholar] [CrossRef]
- Zhang, X.; Ng, W.L.; Wang, P.; Tian, L.; Werner, E.; Wang, H.; Doetsch, P.; Wang, Y. MicroRNA-21 modulates the levels of reactive oxygen species by targeting SOD3 and TNFα. Cancer Res. 2012, 72, 4707–4713. [Google Scholar] [CrossRef]
- Karshovska, E.; Wei, Y.; Subramanian, P.; Mohibullah, R.; Geißler, C.; Baatsch, I.; Popal, A.; Campos, J.C.; Exner, N.; Schober, A. HIF-1α (Hypoxia-Inducible Factor-1α) Promotes Macrophage Necroptosis by Regulating miR-210 and miR-383. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 583–596. [Google Scholar] [CrossRef]
Figure 1.
The process of microRNA biogenesis and the control of gene expression.
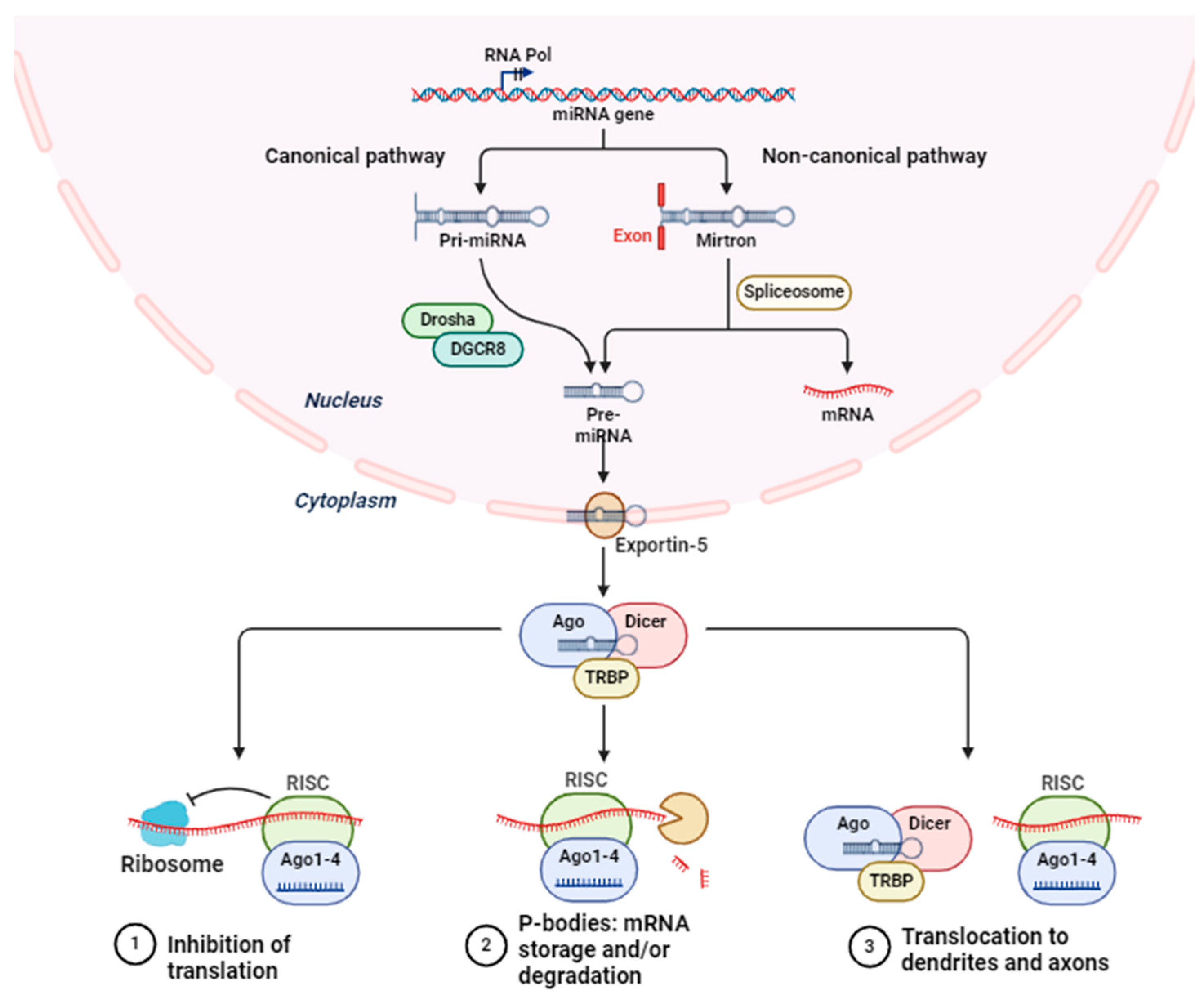
Figure 2.
miR-mediated control of Nrf2 signaling pathway.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Saha, S. Role of microRNA in Oxidative Stress. Stresses 2024, 4, 269-281. https://doi.org/10.3390/stresses4020016
AMA Style
Saha S. Role of microRNA in Oxidative Stress. Stresses. 2024; 4(2):269-281. https://doi.org/10.3390/stresses4020016
Chicago/Turabian StyleSaha, Sarmistha. 2024. "Role of microRNA in Oxidative Stress" Stresses 4, no. 2: 269-281. https://doi.org/10.3390/stresses4020016