
Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Bacillus megaterium LVN01 Using Biogas Digestate

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Study Area and Bacterial Strain
2.3. Carbon Source
2.4. Culture Media
2.5. Activation of Bacterial Strain and Inoculum Preparation
2.5.1. Activation of B. megaterium LVN01
2.5.2. Inoculum Preparation
2.5.3. Characterization of B. megaterium and Visualization of the Accumulated PHA
2.6. Aerobic Fermentative Processes
2.7. Extraction and Purification of PHA
2.8. PHA Characterization
2.8.1. Gas Chromatography–Mass Spectrometry (GC–MS/SIM)
2.8.2. Nuclear Magnetic Resonance Spectroscopy (1H-NMR and 13C-NMR)
2.9. Thermal Properties of PHA
2.9.1. Differential Scanning Calorimetry
2.9.2. Thermogravimetric Analysis
2.10. Analysis and Presentation of Results
3. Results
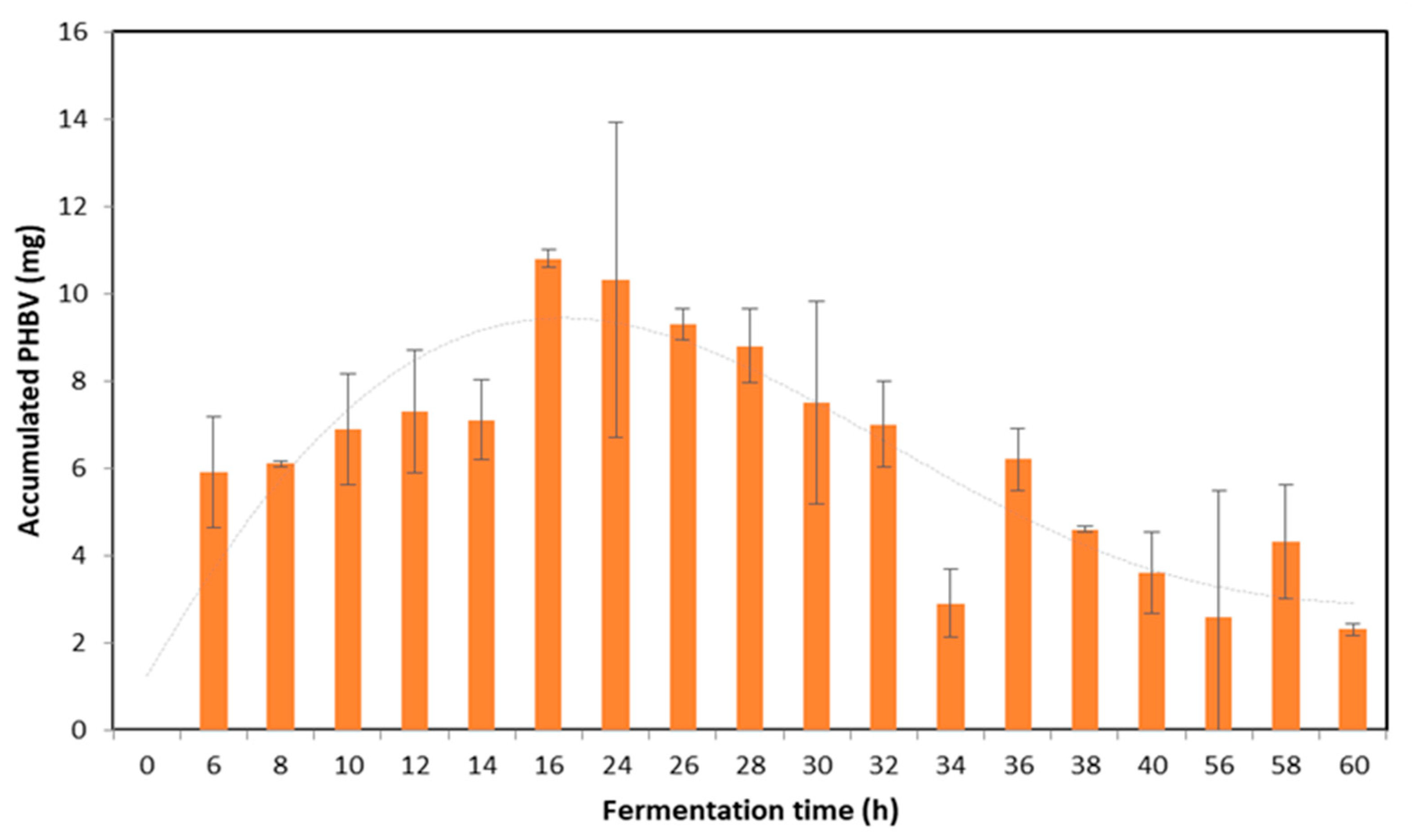
3.1. Fermentation Kinetics
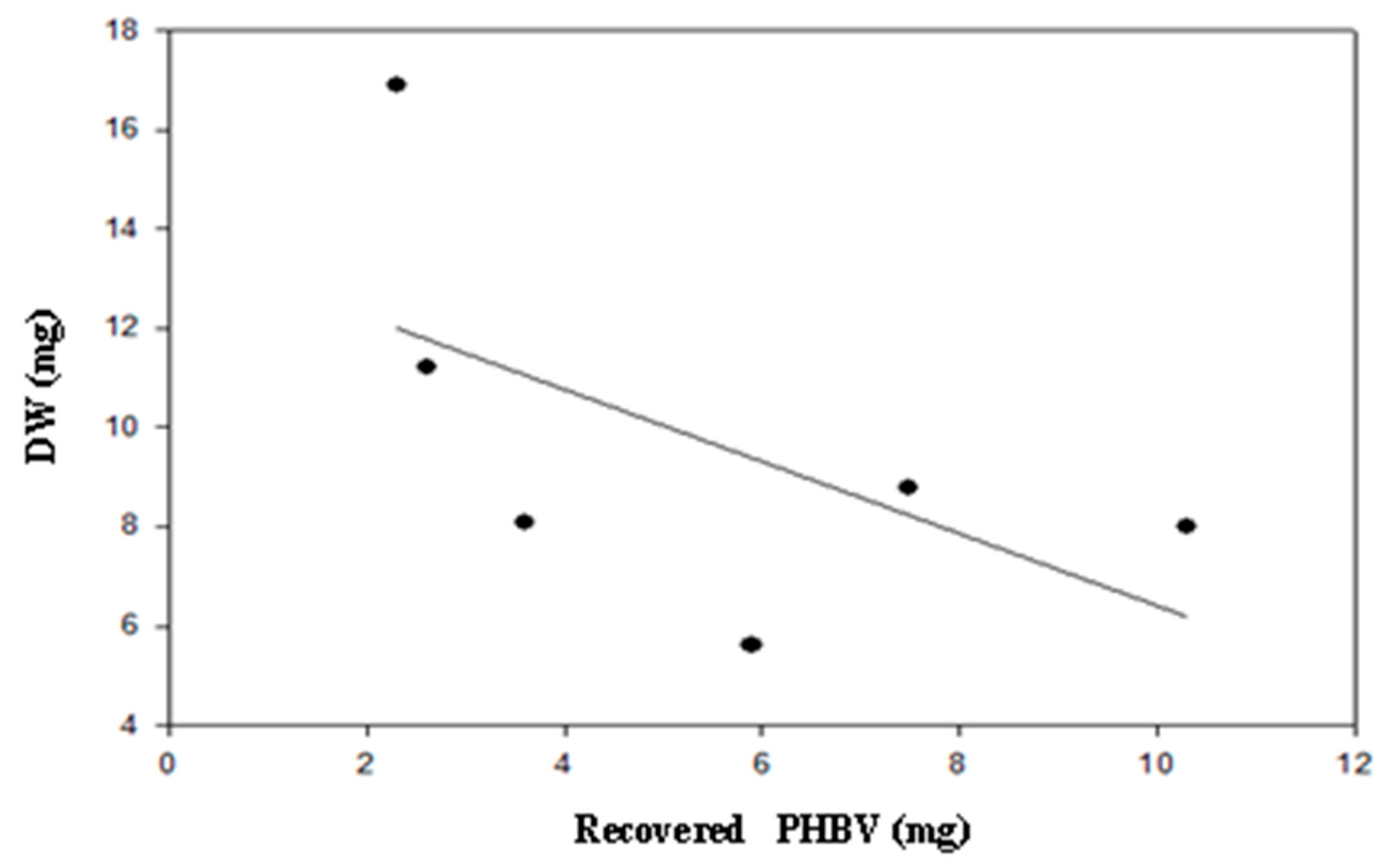
3.2. Poly(3-Hydroxybutyrate)-Co-(3-Hydroxyvalerate) (PHBV) Recovery
3.3. PHBV Characterization
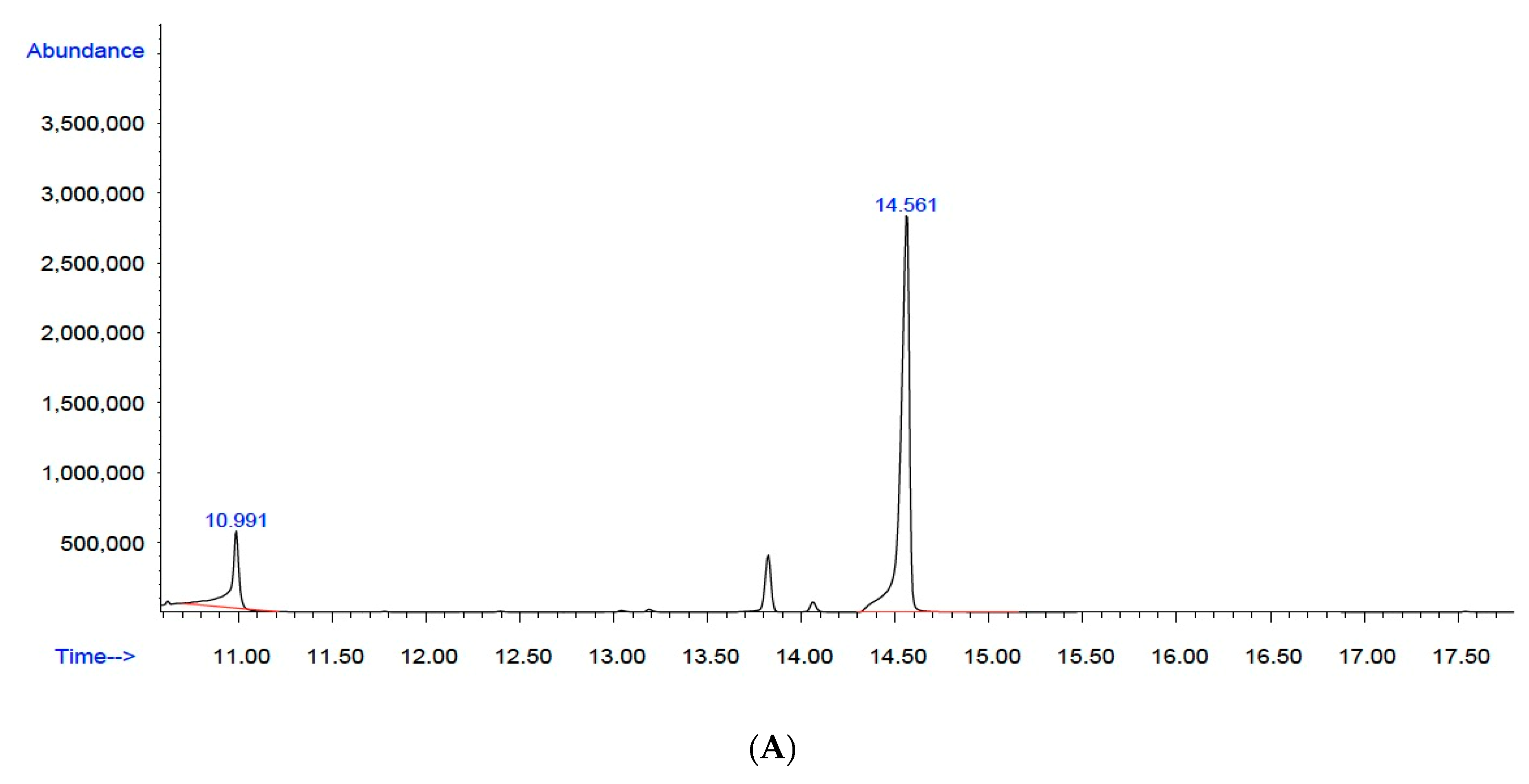
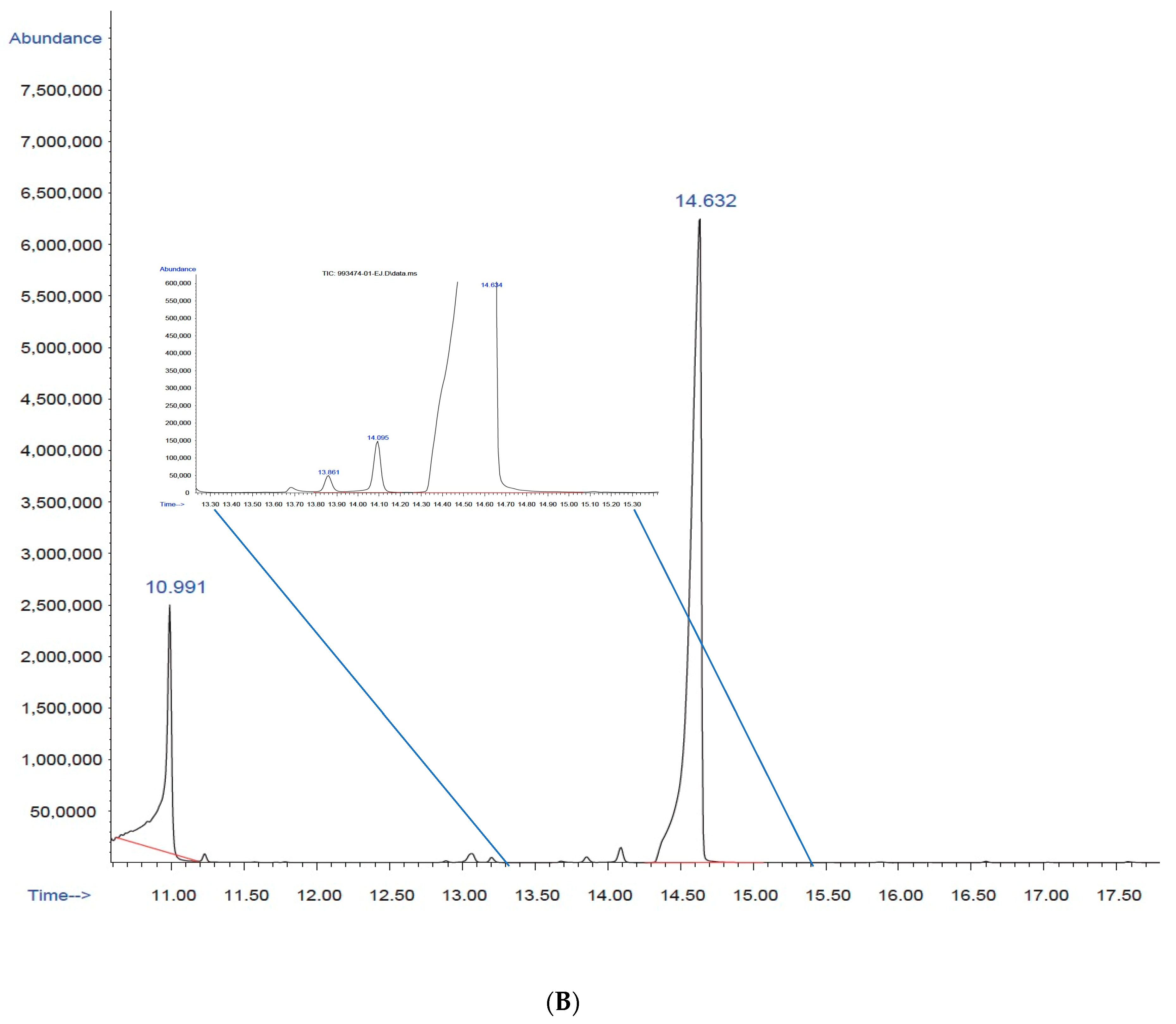
3.3.1. Gas Chromatography with Mass Selective Detector (GC–MS)
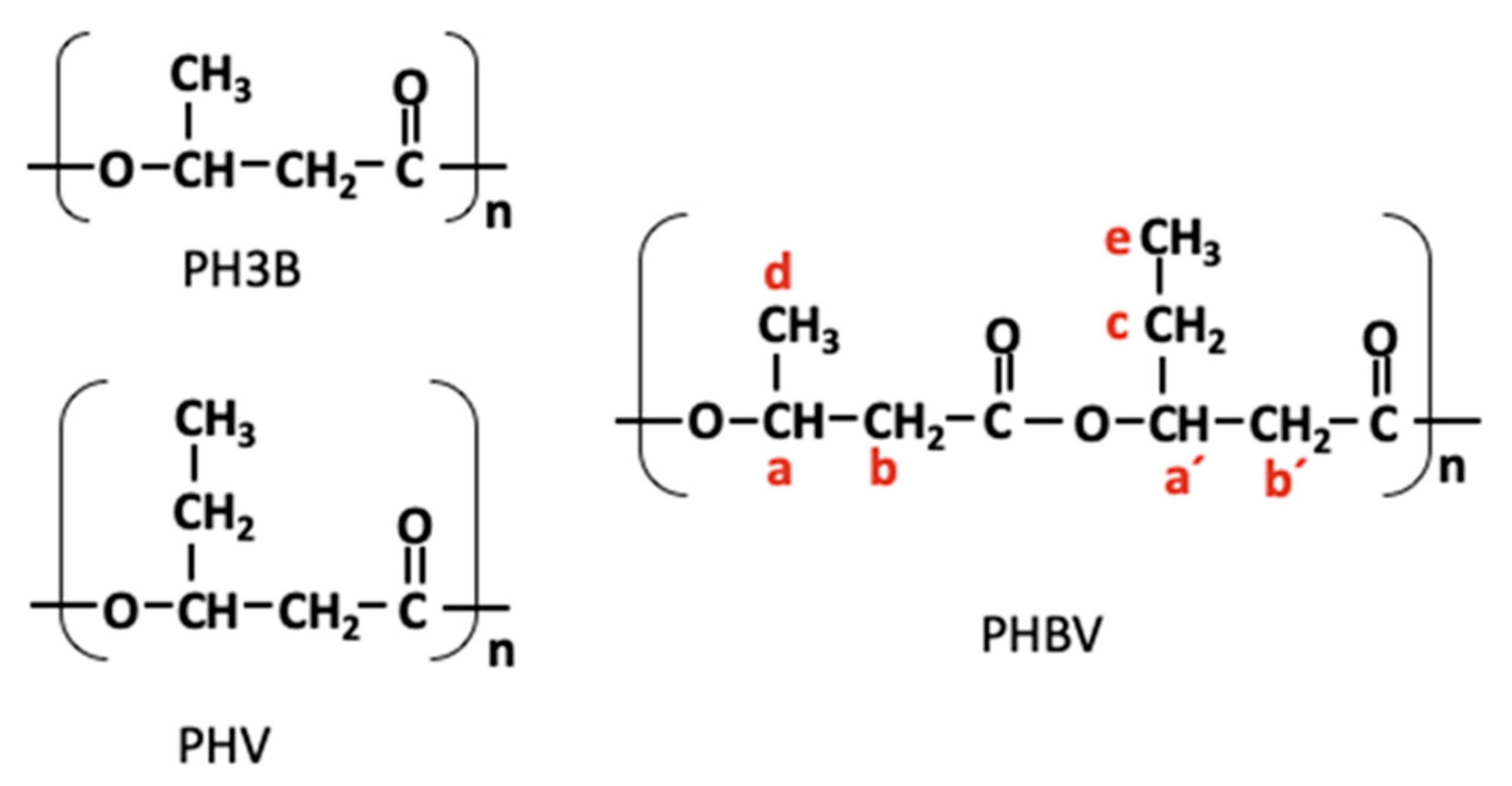
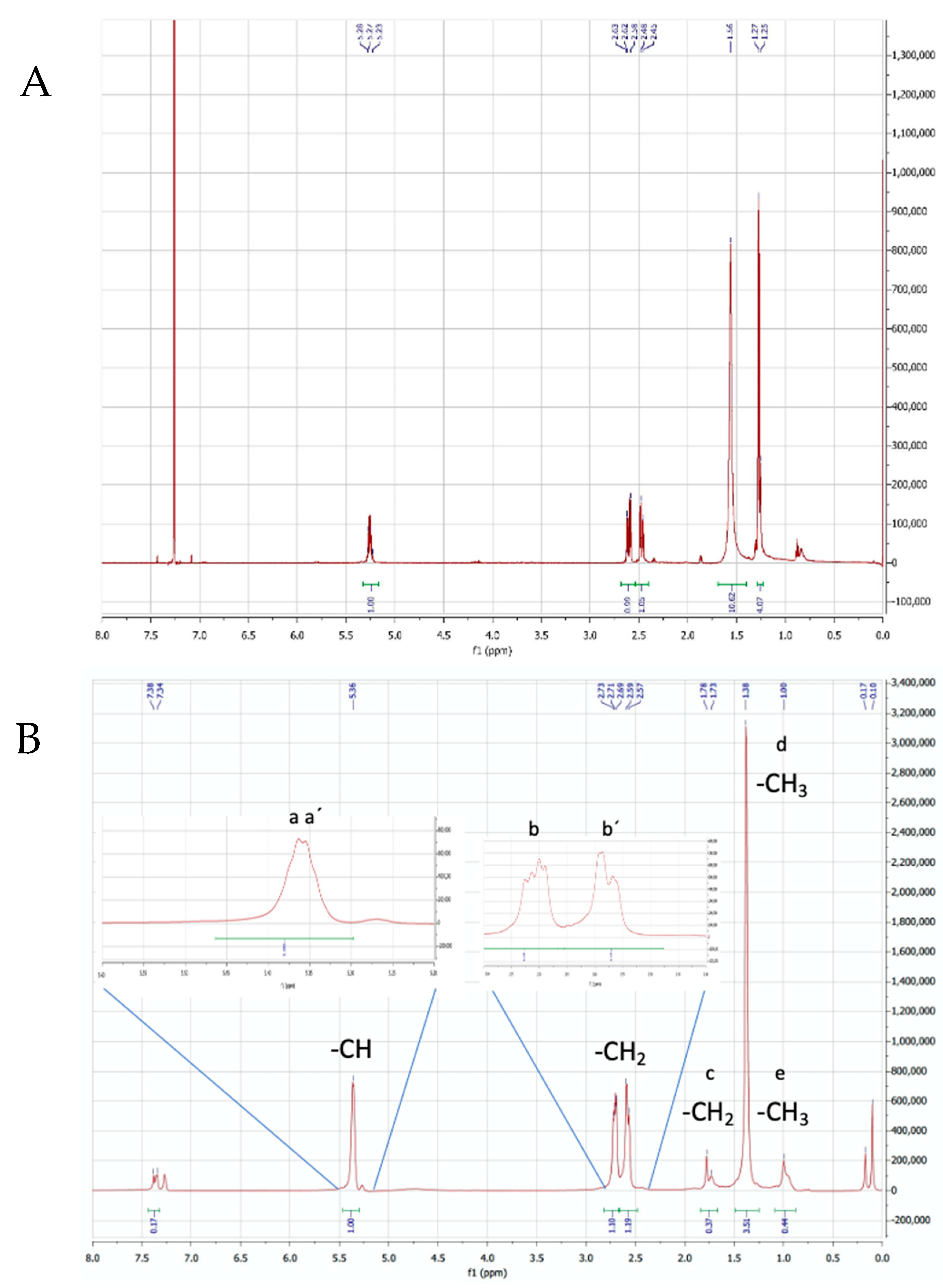
3.3.2. Nuclear Magnetic Resonance (NMR)
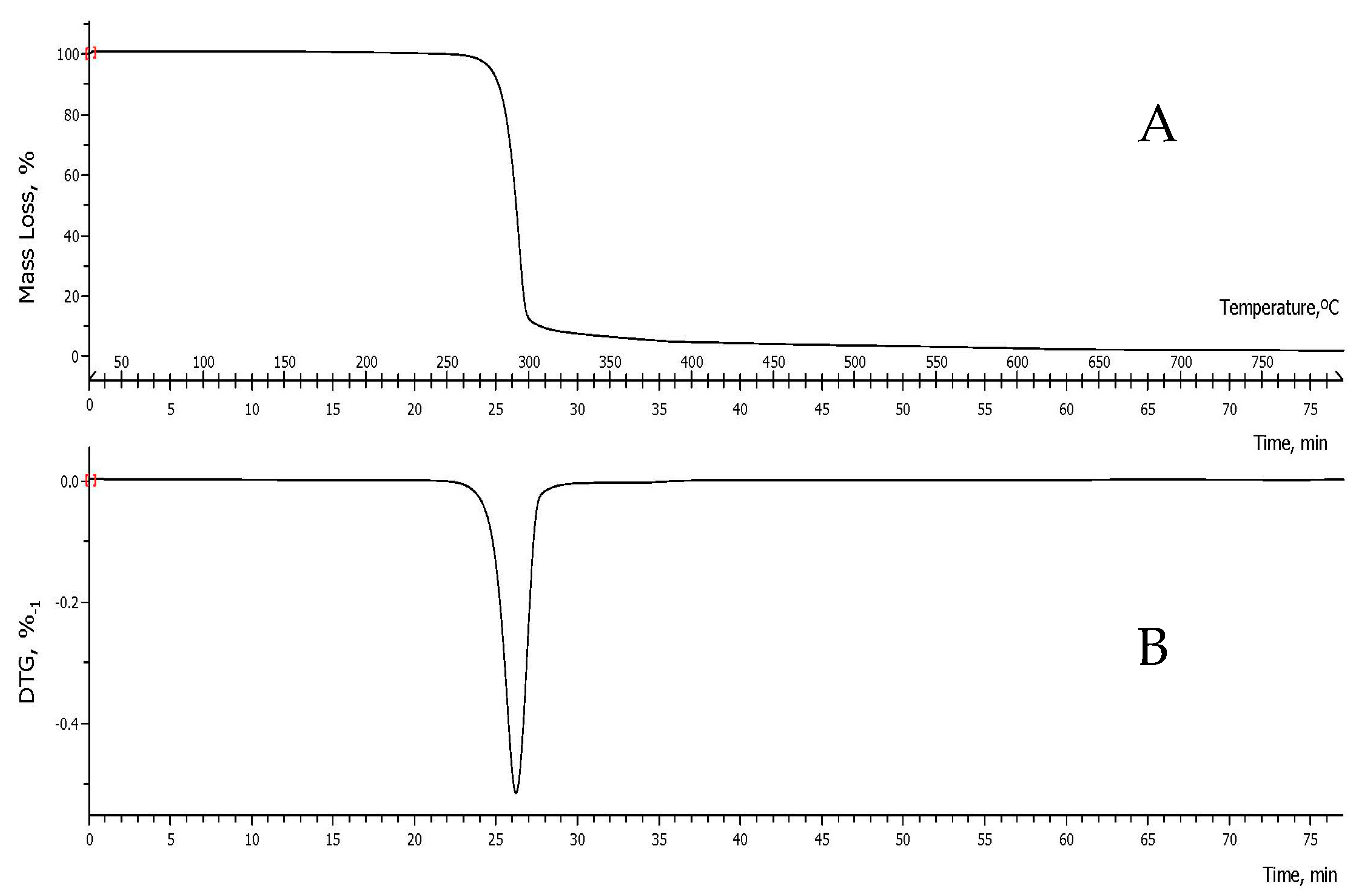
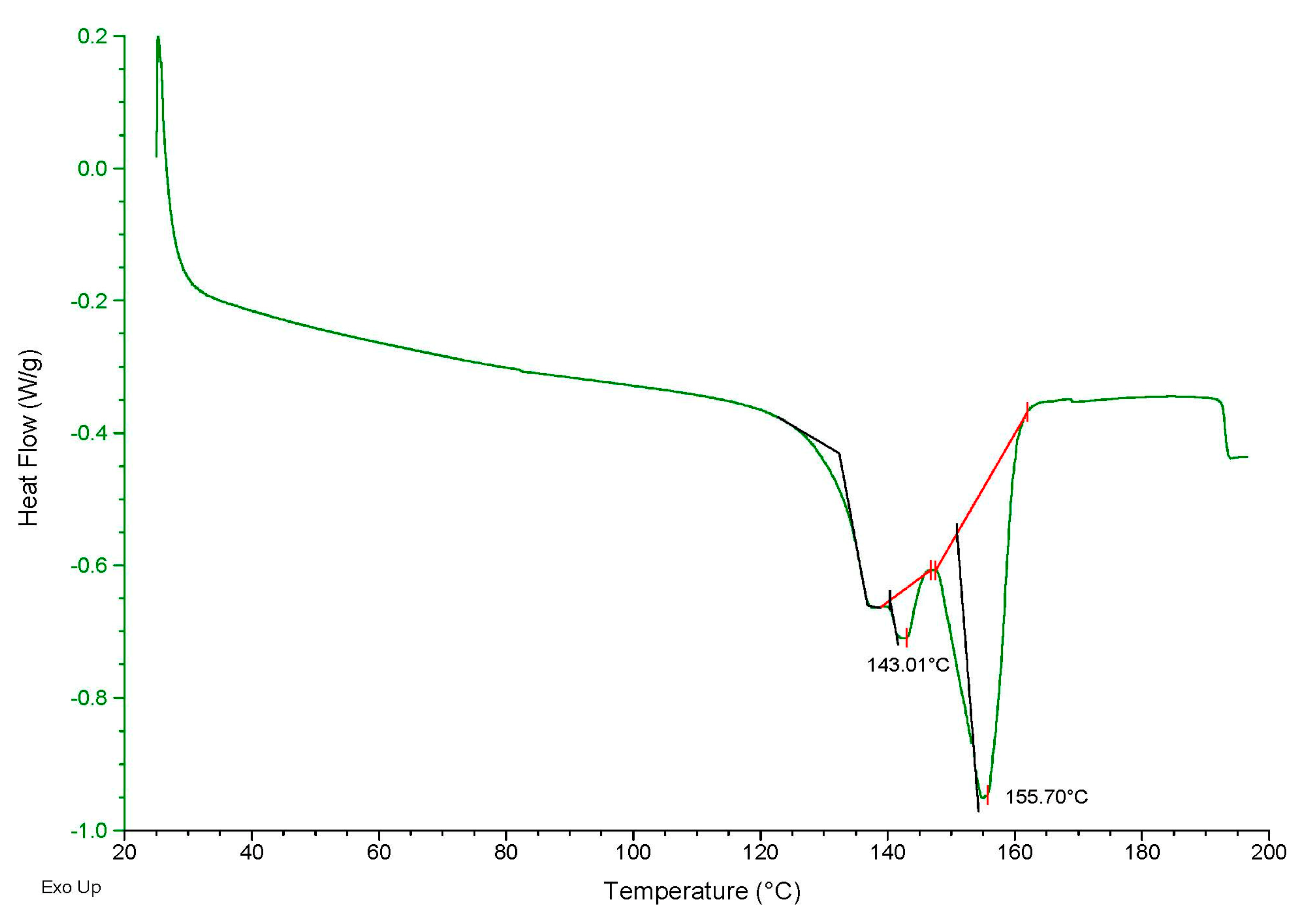
3.4. Determination of Thermal Properties
- ΔHf: Enthalpy of fusion of PHB produced by B. megaterium LVN01;
- ΔHf: Enthalpy of fusion of pure PHB, equivalent to 109 J g−1 [42].
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaso Sánchez, M.A. El surgimiento de los bioplásticos: Un estudio de nichos tecnológicos. Acta Univ. 2020, 30, 1–24. [Google Scholar]
- Plastics Europe, Enabling a Sustainable Future. Plastics—The Fast Facts. Infografía. 2023. Available online: https://plasticseurope.org/es/plastics-europe-publica-plastics-the-fast-facts-2023/ (accessed on 20 March 2024).
- Saratale, R.G.; Cho, S.K.; Saratale, G.D.; Kadam, A.A.; Ghodake, G.S.; Kumar, M.; Bh Bharagava, R.N.; Kumar, G.; Kim, D.S.; Mulla, S.I.; et al. A comprehensive overview and recent advances on polyhydroxyalkanoates (PHA) production using various organic waste streams. Bioresour. Technol. 2021, 325, 124685. [Google Scholar]
- Sohn, Y.J.; Kim, H.T.; Baritugo, K.A.; Jo, S.Y.; Song, H.M.; Park, S.Y.; Park, S.K.; Pyo, J.; Cha, H.G.; Kim, H.; et al. Recent advances in sustainable plastic upcycling and biopolymers. Biotechnol. J. 2020, 15, 1900489. [Google Scholar] [CrossRef] [PubMed]
- Organización de las Naciones Unidas. Todo lo que necesitas saber sobre la contaminación por plásticos. ONU programa para el medio ambiente. Available online: https://www.unep.org/es/noticias-y-reportajes/reportajes/todo-lo-que-necesitas-saber-sobre-la-contaminacion-por-plasticos (accessed on 25 April 2023).
- Grigore, M.E.; Grigorescu, R.M.; Iancu, L.; Ion, R.M.; Zaharia, C.; Andrei, E.R. Methods of synthesis, properties and biomedical applications of polyhydroxyalkanoates: A review. J. Biomater. Sci. Polym. Ed. 2019, 30, 695–712. [Google Scholar] [CrossRef] [PubMed]
- Madhumitha Jaganmohan. Available online: https://www.statista.com/aboutus/our-research-commitment/2234/madhumitha-jaganmohan (accessed on 20 March 2024).
- Anjum, A.; Zuber, M.; Zia, K.M.; Noreen, A.; Anjum, M.N.; Tabasum, S. Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: A review of recent advancements. Int. J. Biol. Macromol. 2016, 89, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Kaniuk, Ł.; Stachewicz, U. Development and advantages of biodegradable PHA polymers based on electrospun PHBV fibers for tissue engineering and other biomedical applications. ACS Biomater. Sci. Eng. 2021, 7, 5339–5362. [Google Scholar] [CrossRef]
- Koller, M. Production of polyhydroxyalkanoate (PHA) biopolyesters by extremophiles. MOJ Polym. Sci. 2017, 1, 1–19. [Google Scholar] [CrossRef]
- Cal, A.J.; Kibblewhite, R.E.; Sikkema, W.D.; Torres, L.F.; Hart-Cooper, W.M.; Orts, W.J.; Lee, C.C. Production of polyhydroxyalkanoate copolymers containing 4-hydroxybutyrate in engineered Bacillus megaterium. Int. J. Biol. Macromol. 2021, 168, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Moreno, S.A.; Marín Montoya, M.A.; Mora Martínez, A.L.; Yepes Pérez, M.D.S. Identification of polyhydroxyalkanoate-producing bacteria in soils contaminated with fique wastes. Rev. Colomb. De Biotecnol. 2012, 14, 89–100. [Google Scholar]
- Dhangdhariya, J.H.; Dubey, S.; Trivedi, H.B.; Pancha, I.; Bhatt, J.K.; Dave, B.P.; Mishra, S. Polyhydroxyalkanoate from marine Bacillus megaterium using CSMCRI’s Dry Sea Mix as a novel growth medium. Int. J. Biol. Macromol. 2015, 76, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Ray, S.; Kalia, V.C. Production of co-polymers of polyhydroxyalkanoates by regulating the hydrolysis of biowastes. Bioresour. Technol. 2016, 200, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Jamil, N. Polyhydroxyalkanoates (PHA) production in bacterial co-culture using glucose and volatile fatty acids as carbon source. J. Basic Microbiol. 2018, 58, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.H.; Sawant, S.S.; Kim, B.S. Production of polyhydroxyalkanoates by Ralstonia eutropha from volatile fatty acids. Korean J. Chem. Eng. 2013, 30, 2223–2227. [Google Scholar] [CrossRef]
- Martinez, G.A.; Bertin, L.; Scoma, A.; Rebecchi, S.; Braunegg, G.; Fava, F. Production of polyhydroxyalkanoates from dephenolised and fermented olive mill wastewaters by employing a pure culture of Cupriavidus necator. Biochem. Eng. J. 2015, 97, 92–100. [Google Scholar] [CrossRef]
- Ferre-Guell, A.; Winterburn, J. Increased production of polyhydroxyalkanoates with controllable composition and consistent material properties by fed-batch fermentation. Biochem. Eng. J. 2019, 141, 35–42. [Google Scholar] [CrossRef]
- Hermann-Krauss, C.; Koller, M.; Muhr, A.; Fasl, H.; Stelzer, F.; Braunegg, G. Archaeal production of polyhydroxyalkanoate (PHA) co-and terpolyesters from biodiesel industry-derived by-products. Archaea 2013, 2013, 129268. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, R.; Pandey, J.P.; Gaur, V.K.; Gnansounou, E.; Sindhu, R. Critical overview of biomass feedstocks as sustainable substrates for the production of polyhydroxybutyrate (PHB). Bioresour. Technol. 2020, 311, 123536. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.H.; Wainaina, S.; Taherzadeh, M.J.; Åkesson, D.; Ferreira, J.A. Production of polyhydroxyalkanoates (PHAs) by Bacillus megaterium using food waste acidogenic fermentation-derived volatile fatty acids. Bioengineered 2021, 12, 2480–2498. [Google Scholar] [CrossRef]
- Kleerebezem, R.; Joosse, B.; Rozendal, R.; Van Loosdrecht, M.C. Anaerobic digestion without biogas? Rev. Environ. Sci. Bio/Technol. 2015, 14, 787–801. [Google Scholar] [CrossRef]
- Cadavid-Rodríguez, L.S.; Castro-López, V.E.; Placido, J. Evaluation of optimal fermentation conditions for volatile fatty acids production from artisanal fish waste. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Gómez-Cardozo, J.R.; Velasco-Bucheli, R.; Marín-Pareja, N.; Ruíz-Villadiego, O.S.; Correa-Londoño, G.A.; Mora-Martínez, A.L. Fed-batch production and characterization of polyhydroxybutyrate by Bacillus megaterium LVN01 from residual glycerol. Dyna 2020, 87, 111–120. [Google Scholar] [CrossRef]
- Salazar, A. Parámetros Operacionales Óptimos Para La Producción de Bioplásticos a Partir de Fuentes Renovables Azucaradas; Universidad Nacional de Colombia: Medellín, Colombia, 2011. [Google Scholar]
- Rai, A.K.; Swapna, H.C.; Bhaskar, N.; Halami, P.M.; Sachindra, N.M. Effect of fermentation ensilaging on recovery of oil from fresh water fish viscera. Enzym. Microb. Technol. 2010, 46, 9–13. [Google Scholar] [CrossRef]
- López-Jácome, L.E.; Hernández-Durán, M.; Colín-Castro, C.A.; Ortega-Peña, S.; Cerón-González, G.; Franco-Cendejas, R. Las tinciones básicas en el laboratorio de microbiología. Investig. En Discapac. 2014, 3, 10–18. [Google Scholar]
- Tinción de Esporas: Fundamento, Técnicas y Usos. Lifeder. Available online: https://www.lifeder.com/tincion-de-esporas/ (accessed on 18 January 2024).
- Pérez, R.; Juárez, M.; Rodríguez, L.P. Manual de Laboratorio de Técnicas Microbiológicas; Departamento de Ciencias Básicas Academia de Microbiología. Instituto Politécnico Nacional: Pereira, Colombia, 2011. [Google Scholar]
- Spiekermann, P.; Rehm, B.H.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A sensitive, viable-colony staining method using Nile red for direct screening of bacteria that accumulate polyhydroxyalkanoic acids and other lipid storage compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Ostle, A.G.; Holt, J.G. Nile blue A as a fluorescent stain for poly-beta-hydroxybutyrate. Appl. Environ. Microbiol. 1982, 44, 238–241. [Google Scholar] [CrossRef]
- ISO 12966-2:2017; Animal and Vegetable Fats and Oils—Gas Chromatography of Fatty Acid Methyl Esters—Part 2: Preparation of methyl Esters of Fatty Acids. International Organization for Standardization: Geneva, Switzerland, 2017.
- Pradhan, S.; Dikshit, P.K.; Moholkar, V.S. Production, characterization, and applications of biodegradable polymer: Polyhydroxyalkanoates. In Advances in Sustainable Polymers: Synthesis, Fabrication and Characterization; Springer: Singapore, 2020; pp. 51–94. [Google Scholar]
- Liu, J.; Zhao, Y.; Diao, M.; Wang, W.; Hua, W.; Wu, S.; Chen, P.; Ruan, R.; Cheng, Y. Poly (3-hydroxybutyrate-co-3-hydroxyvalerate) production by Rhodospirillum rubrum using a two-step culture strategy. J. Chem. 2019, 2019, 8369179. [Google Scholar] [CrossRef]
- Jakić, M.; Vrandečić, N.S.; Erceg, M. Thermal degradation of poly (3-hydroxybutyrate)/poly (ethylene oxide) blends: Thermogravimetric and kinetic analysis. Eur. Polym. J. 2016, 81, 376–385. [Google Scholar] [CrossRef]
- Vahabi, H.; Michely, L.; Moradkhani, G.; Akbari, V.; Cochez, M.; Vagner, C.; Renard, E.; Saeb, M.R.; Langlois, V. Thermal stability and flammability behavior of poly (3-hydroxybutyrate)(PHB) based composites. Materials 2019, 12, 2239. [Google Scholar] [CrossRef]
- Thiré, R.M.; Arruda, L.C.; Barreto, L.S. Morphology and thermal properties of poly (3-hydroxybutyrate-co-3-hydroxyvalerate)/attapulgite nanocomposites. Mater. Res. 2011, 14, 340–344. [Google Scholar] [CrossRef]
- Abbasi, M.; Coats, E.R.; McDonald, A.G. Green solvent extraction and properties characterization of Poly (3-hydroxybutyrate-co-3-hydroxyvalerate) biosynthesized by mixed microbial consortia fed fermented dairy manure. Bioresour. Technol. Rep. 2022, 28, 101065. [Google Scholar] [CrossRef]
- Abbasi, M.; Pokhrel, D.; Coats, E.R.; Guho, N.M.; McDonald, A.G. Effect of 3-hydroxyvalerate content on thermal, mechanical, and rheological properties of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) biopolymers produced from fermented dairy manure. Polymers 2022, 14, 4140. [Google Scholar] [CrossRef]
- Guho, N.M.; Pokhrel, D.; Abbasi, M.; McDonald, A.G.; Alfaro, M.; Brinkman, C.K.; Coats, E.R. Pilot-scale production of poly-3-hydroxybutyrate-co-3-hydroxyvalerate from fermented dairy manure: Process performance, polymer characterization, and scale-up implications. Bioresour. Technol. Rep. 2020, 12, 100588. [Google Scholar] [CrossRef]
- Souza Junior, O.F.; Staffa, L.H.; Costa, L.C.; Chinelatto, M.A. Thermal and Rheological Behavior of Binary Blends of Poly (hydroxybutyrate-co-hydroxyvalerate) and Poly (ethylene-co-vinyl acetate) with Different Vinyl Acetate Content. Macromol. Symp. 2019, 383, 1800020. [Google Scholar] [CrossRef]
- Chan, C.H.; Kummerlöwe, C.; Kammer, H.W. Crystallization and melting behavior of poly (3-hydroxybutyrate)-based blends. Macromol. Chem. Phys. 2004, 205, 664–675. [Google Scholar] [CrossRef]
- Bora, L.; Das, R.; Gohain, D. A novel melt stable and high tensile strength biopolymer (polyhydroxyalkanoates) from Bacillus megaterium (MTCC10086) and its characterization. J. Basic Microbiol. 2014, 54, 1012–1016. [Google Scholar] [CrossRef]
- Castillo, D. Efecto del gen fadH1 en la Producción de PHA Contenido Monómeros Insaturados por Pseudomona Putida; Pontificia Universidad Javeriana: Bogotá, Colombia, 2008. [Google Scholar]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Wang, J.; Liu, S.; Huang, J.; Qu, Z. A review on polyhydroxyalkanoate production from agricultural waste Biomass: Development, Advances, circular Approach, and challenges. Bioresour. Technol. 2021, 342, 126008. [Google Scholar] [CrossRef] [PubMed]
- Kosseva, M.R.; Rusbandi, E. Trends in the biomanufacture of polyhydroxyalkanoates with focus on downstream processing. Int. J. Biol. Macromol. 2018, 107, 762–778. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Murphy, R.; Narayan, R.; Davies, G. Biodegradable and compostable alternatives to conventional plastics. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2127–2139. [Google Scholar] [CrossRef] [PubMed]
- Gómez Cardozo, J.R.; Velasco-Bucheli, R.; Del Cerro, C.; Mata, I.D.L.; Mora Martínez, A.L. Engineering of Bacillus megaterium for improving PHA production from glycerol. Asia Pac. J. Mol. Biol. Biotechnol. 2019, 27, 64–72. [Google Scholar]
- Gómez Cardozo, J.R.; Mora Martínez, A.L.; Yepes Pérez, M.; Correa Londoño, G.A. Production and characterization of polyhydroxyalkanoates and native microorganisms synthesized from fatty waste. Int. J. Polym. Sci. 2016, 2016, 6541718. [Google Scholar] [CrossRef]
- Salazar, A.; Yepes, M.; Correa, G.; Mora, A. Polyhydroxyalkanoate production from unexplored sugar substrates. Dyna 2014, 81, 73–77. [Google Scholar] [CrossRef]
- Cardona, A.C.; Mora, A.L.; Marín, M. Identificación molecular de bacterias productoras de polihidroxialcanoatos en subproductos de lácteos y caña de azúcar. Rev. Fac. Nac. De Agron. Medellín 2013, 66, 7129–7140. [Google Scholar]
- Salgaonkar, B.B.; Mani, K.; Braganca, J.M. Characterization of polyhydroxyalkanoates accumulated by a moderately halophilic salt pan isolate Bacillus megaterium strain H16. J. Appl. Microbiol. 2013, 114, 1347–1356. [Google Scholar] [CrossRef]
- Alkotaini, B.; Sathiyamoorthi, E.; Kim, B.S. Potential of Bacillus megaterium for production of polyhydroxyalkanoates using the red algae Gelidium amansii. Biotechnol. Bioprocess Eng. 2015, 20, 856–860. [Google Scholar] [CrossRef]
- Winnacker, M. Polyhydroxyalkanoates: Recent advances in their synthesis and applications. Eur. J. Lipid Sci. Technol. 2019, 121, 1900101. [Google Scholar] [CrossRef]
- Chen, G.Q.; Jiang, X.R.; Guo, Y. Synthetic biology of microbes synthesizing polyhydroxyalkanoates (PHA). Synth. Syst. Biotechnol. 2016, 1, 236–242. [Google Scholar] [CrossRef]
- Tian, P.; Shang, L.; Ren, H.; Mi, Y.; Fan, D.; Jiang, M. Biosynthesis of polyhydroxyalkanoates: Current research and development. Afr. J. Biotechnol. 2009, 8, 709–714. [Google Scholar]
- Cavalheiro, J.M.; Raposo, R.S.; De Almeida, M.C.M.; Cesário, M.T.; Sevrin, C.; Grandfils, C.; Da Fonseca, M. Effect of cultivation parameters on the production of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) and poly (3-hydroxybutyrate-4-hydroxybutyrate-3-hydroxyvalerate) by Cupriavidus necator using waste glycerol. Bioresour. Technol. 2012, 111, 391–397. [Google Scholar] [CrossRef]
- Polyhydroxyalkanoate (PHA) Market by Type (Short Chain Length, Medium Chain Lenth), Production Methods (Sugar Fermentation, Vegetable Oil Fermentation), Application (Packing & Food Services Biomedical) and Region-Global Forecast to 2028. Available online: https://www.marketsandmarkets.com/Market-Reports/pha-market-395.html?gad_source=1&gclid=Cj0KCQjwhtWvBhD9ARIsAOP0GogViHNhj3GJBVD8HaHD2hI_z9qSSB0dUmUTmNE-rvudbLClu9nmLCIaAgzzEALw_wcB (accessed on 15 May 2024).
- Kumar, M.; Rathour, R.; Singh, R.; Sun, Y.; Pandey, A.; Gnansounou, E.; Lin, K.Y.; Tsang, D.C.; Thakur, I.S. Bacterial polyhydroxyalkanoates: Opportunities, challenges, and prospects. J. Clean. Prod. 2020, 263, 121500. [Google Scholar] [CrossRef]
- Abd El-malek, F.; Khairy, H.; Farag, A.; Omar, S. The sustainability of microbial bioplastics, production and applications. Int. J. Biol. Macromol. 2020, 157, 319–328. [Google Scholar] [CrossRef]
- Loizidou, M.; Moustakas, K.; Rehan, M.; Nizami, A.S.; Tabatabaei, M. New developments in sustainable waste-to-energy systems. Renew. Sustain. Energy Rev. 2021, 151, 111581. [Google Scholar] [CrossRef]
- Lambert, S.; Wagner, M. Environmental performance of bio-based and biodegradable plastics: The road ahead. Chem. Soc. Rev. 2017, 46, 6855–6871. [Google Scholar] [CrossRef]
- Xiang, H.; Wen, X.; Miu, X.; Li, Y.; Zhou, Z.; Zhu, M. Thermal depolymerization mechanisms of poly (3-hydroxybutyrate-co-3-hydroxyvalerate). Prog. Nat. Sci. Mater. Int. 2016, 26, 58–64. [Google Scholar] [CrossRef]
- Miranda, D.A.; Marín, K.; Sundman, O.; Hedenström, M.; Quillaguaman, J.; Gorzsás, A.; Brosrom, M.; Carlborg, M.; Lundvist, J.; Romero-Soto LJonsson, L.J.; et al. Production and characterization of poly (3-hydroxybutyrate) from Halomonas boliviensis LC1 cultivated in hydrolysates of quinoa stalks. Fermentation 2023, 9, 556. [Google Scholar] [CrossRef]
- Sirohi, R.; Pandey, J.P.; Tarafdar, A.; Agarwal, A.; Chaudhuri, S.K.; Sindhu, R. An environmentally sustainable green process for the utilization of damaged wheat grains for poly-3-hydroxybutyrate production. Environ. Technol. Innov. 2021, 21, 101271. [Google Scholar] [CrossRef]
- Zhuikov, V.A.; Zhuikova, Y.V.; Makhina, T.K.; Myshkina, V.L.; Rusakov, A.; Useinov, A.; Voinova, V.V.; Bonartseva, G.A.; Berlin, A.A.; Bonartsev, A.P.; et al. Comparative structure-property characterization of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) s films under hydrolytic and enzymatic degradation: Finding a transition point in 3-hydroxyvalerate content. Polymers 2020, 12, 728. [Google Scholar] [CrossRef]
- Mohapatra, S.; Pattnaik, S.; Maity, S.; Sharma, S.; Akhtar, J.; Pati, S.; Samantaray, D.P.; Varma, A. Comparative analysis of PHAs production by Bacillus megaterium OUAT 016 under submerged and solid-state fermentation. Saudi J. Biol. Sci. 2020, 27, 1242–1250. [Google Scholar] [CrossRef]
- Han, J.; Hou, J.; Zhang, F.; Ai, G.; Li, M.; Cai, S.; Liu, H.; Wang, L.; Wang, Z.; Zhang, S.; et al. Multiple propionyl coenzyme A-supplying pathways for production of the bioplastic poly (3-hydroxybutyrate-co-3-hydroxyvalerate) in Haloferax mediterranei. Appl. Environ. Microbiol. 2013, 79, 2922–2931. [Google Scholar] [CrossRef]
- Cheng, H.N.; Biswas, A.; Vermillion, K.; Melendez-Rodriguez, B.; Lagaron, J.M. NMR analysis and triad sequence distributions of poly (3-hydroxybutyrate-co-3-hydroxyvalerate). Polym. Test. 2020, 90, 106754. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, R.; Cai, J.; Liu, Z.; Zheng, Y.; Wang, H.; Li, Q.; He, N. Biosynthesis and thermal properties of PHBV produced from levulinic acid by Ralstonia eutropha. PLoS ONE 2013, 8, e60318. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Bee, G.; Kreuzer, M.; Wenk, C. Novel biodegradable plastics in sheep nutrition-Effects of NaOH pretreatment of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) on in vivo digestibility and on in vitro disappearance (Rusitec). J. Anim. Phys. Anim. Nutrit. 1999, 81, 41–50. [Google Scholar] [CrossRef]
- Elles Montero, A.E.; García Echeverry, C.A. Mezcla Sinérgica Entre Polihidroxibutirato (PHB) y Caucho Natural (Látex) para Obtener un Copolímero. Bachelor’s Thesis, Universidad de Cartagena, Cartagena, Colombia, 2012. [Google Scholar]
- Barham, P.; Keller, A.; Otun, E.; Holmes, P. Crystallization and morphology of a bacterial thermoplastic: Poly-3-hydroxybutyrate. J. Mater. Sci. 1984, 19, 2781–2794. [Google Scholar] [CrossRef]
- Gutiérrez, G.; Getzabeth, M. Producción de Poli-Hidroxialcanoatos por Bacterias del Género Bacillus de Origen Marino. Master’s Thesis, Centro de Investigaciones Biológicas, Madrid, Spain, 2008. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Proton | Chemical Shift δ (ppm) | |
|---|---|---|
| a′ and a | Methin (-CH-) in PHV and PHB | 5.356 and 5.363 |
| b′ and b | Methylene (-CH2) in PHV and PHB | 2.59–2.57 and 2.69–2.73 |
| c | Methylene (-CH2) in PHV | 1.73–1.78 |
| d | Methyl (-CH3) in PHB | 1.38 |
| e | Methyl (-CH3) in PHV | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, A.L.M.; Yepes-Pérez, M.; Contreras, K.A.C.; Moreno, P.E.Z. Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Bacillus megaterium LVN01 Using Biogas Digestate. Appl. Microbiol. 2024, 4, 1057-1078. https://doi.org/10.3390/applmicrobiol4030072
Martínez ALM, Yepes-Pérez M, Contreras KAC, Moreno PEZ. Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Bacillus megaterium LVN01 Using Biogas Digestate. Applied Microbiology. 2024; 4(3):1057-1078. https://doi.org/10.3390/applmicrobiol4030072
Chicago/Turabian StyleMartínez, Amanda Lucía Mora, María Yepes-Pérez, Karent Alexandra Carrero Contreras, and Paola Eliana Zapata Moreno. 2024. "Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Bacillus megaterium LVN01 Using Biogas Digestate" Applied Microbiology 4, no. 3: 1057-1078. https://doi.org/10.3390/applmicrobiol4030072
APA StyleMartínez, A. L. M., Yepes-Pérez, M., Contreras, K. A. C., & Moreno, P. E. Z. (2024). Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Bacillus megaterium LVN01 Using Biogas Digestate. Applied Microbiology, 4(3), 1057-1078. https://doi.org/10.3390/applmicrobiol4030072