
Climate Change Challenges in Temperate and Sub-Tropical Fruit Tree Cultivation


Abstract
1. Introduction
2. Effects of Climate Change on Tree Phenology—Physiology
2.1. Effect of Climate Change on Bud Dormancy
2.2. Impact of Climate Change on Pollination
2.3. Impact of Climate Change on Fruit Growth and Quality
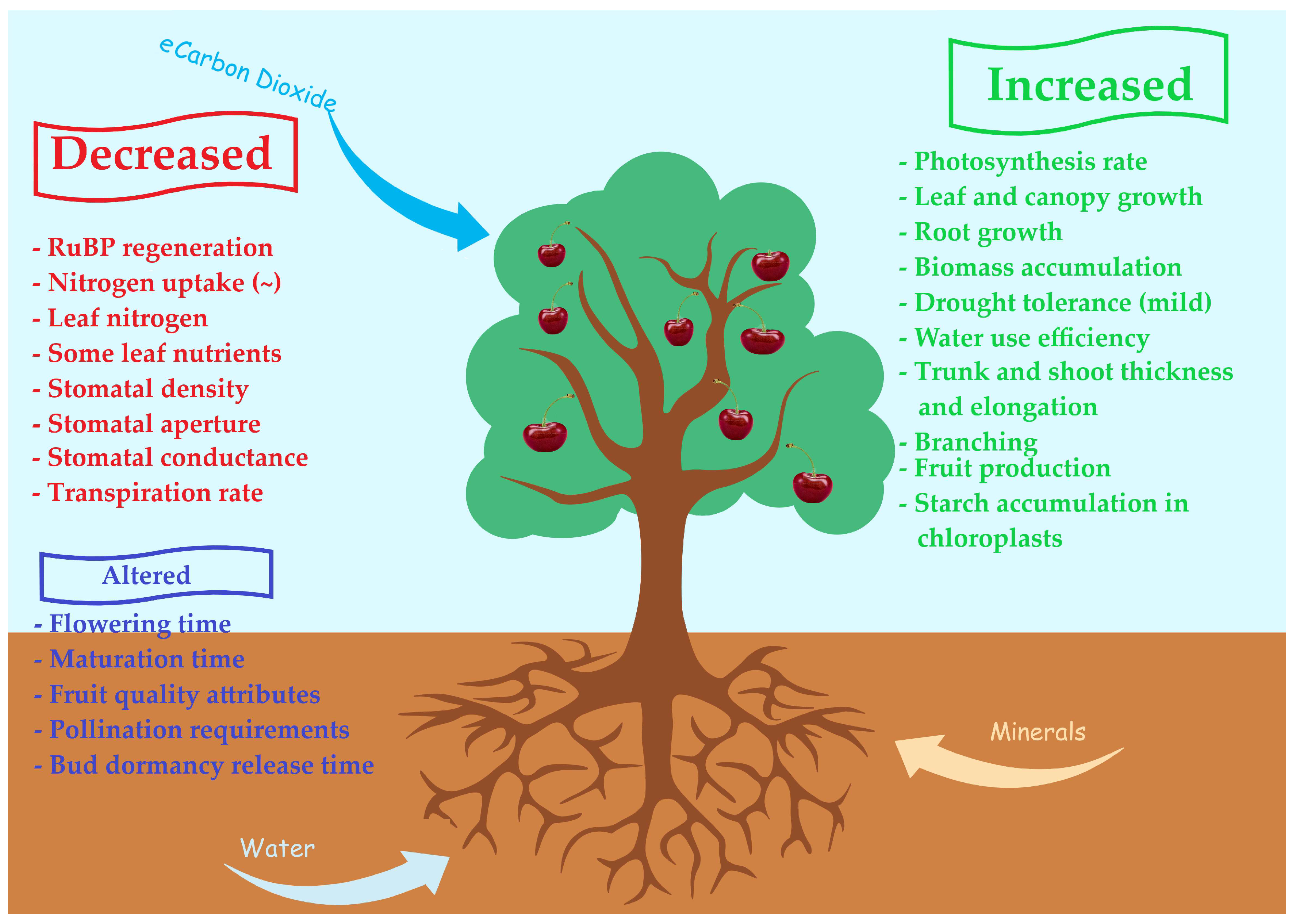
2.4. Impact of Climate Change on Plant Growth
- Altered photosynthesis-related gene expression,
- The feedback inhibition phenomenon described above,
- A limitation in the regeneration of ribulose-1,5 bisphosphate (RuBP) due to a conjugation of inorganic phosphate with the accumulated carbohydrates resulting in lower phosphorus availability,
- The progressive accumulation of starch in the chloroplasts,
- The limited utilization of triose phosphate,
- The decrease in stomatal density along with increased leaf thickness (due to increased mesophyll and vascular tissue cross-sectional area) which leads to reduced CO2 diffusion rates,
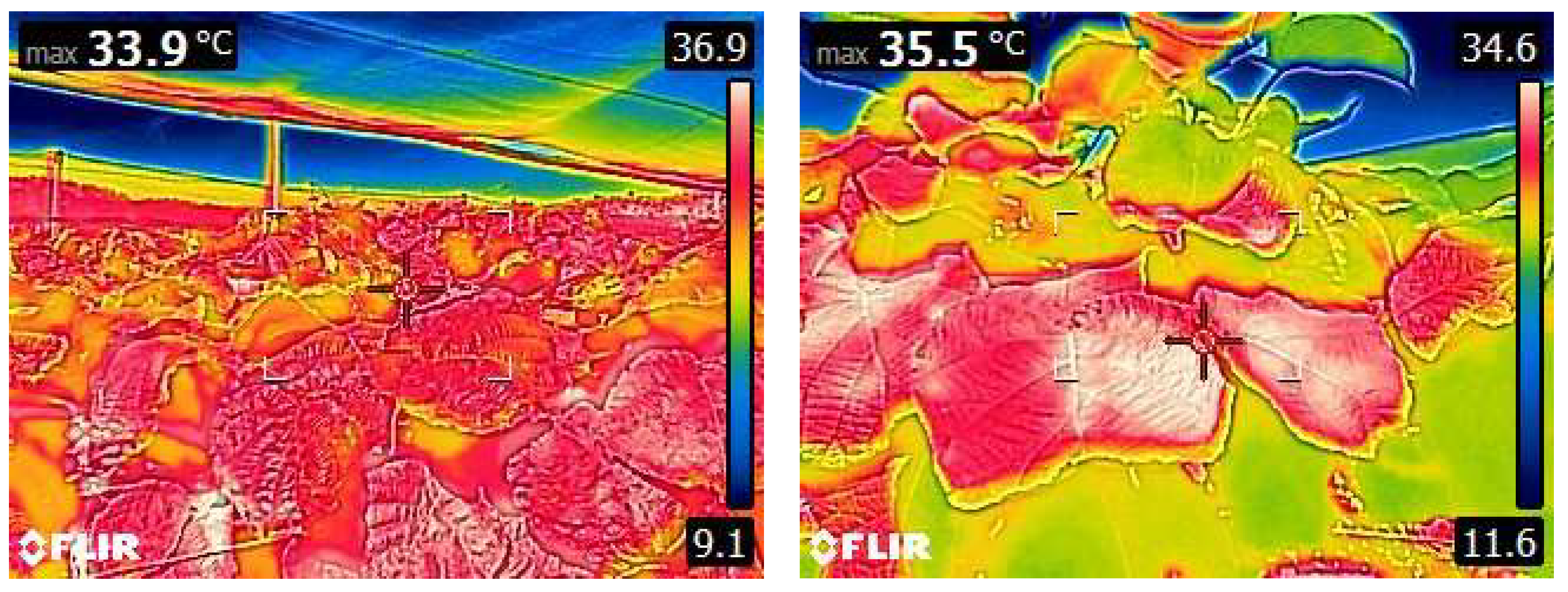
2.5. Effect of Erratic Precipitation or Sudden Rainstorms
3. Impact on Pest, Disease, and Weed Incidence
4. Possible Mitigation Measures
5. Conclusions and Prospects
Funding
Conflicts of Interest
References
- Kunz, A.; Blanke, M.M. Effects of climate change on fruit tree physiology—Based on 55 years of meteorological and phenological data at Klein-Altendorf. Acta Hortic. 2016, 1130, 49–54. [Google Scholar] [CrossRef]
- Roussos, P.A. Climate change impacts on fruit trees and mitigation strategies of adverse effects. AgroLife Sci. J. 2020, 9, 269–276. [Google Scholar]
- Haokip, S.W.; Shankar, K.; Lalrinngheta, J. Climate change and its impact on fruit crops. J. Pharmacogn. Phytochem. 2020, 9, 435–438. [Google Scholar]
- Salama, A.-M.; Ezzat, A.; El-Ramady, H.; Alam-Eldein, S.M.; Okba, S.K.; Elmenofy, H.M.; Hassan, I.F.; Illés, A.; Holb, I.J. Temperate Fruit Trees under Climate Change: Challenges for Dormancy and Chilling Requirements in Warm Winter Regions. Horticulturae 2021, 7, 86. [Google Scholar] [CrossRef]
- Naz, S.; Fatima, Z.; Iqbal, P.; Khan, A.; Zakir, I.; Ullah, H.; Abbas, G.; Ahmed, M.; Mubeen, M.; Hussain, S.; et al. An Introduction to Climate Change Phenomenon. In Building Climate Resilience in Agriculture: Theory, Practice and Future Perspective; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hashmi, M.Z., Eds.; Springer Nature: Cham, Switzerland, 2022; pp. 3–16. [Google Scholar] [CrossRef]
- Rajatiya, J. Climate Change: Impact, Mitigation and Adaptation in Fruit Crops. Intern. J. Pure Appl. Biosci. 2018, 6, 6161. [Google Scholar] [CrossRef]
- Lee, H.; Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.; Trisos, C.; Romero, J.; Aldunce, P.; Barret, K. Climate Change 2023: Synthesis Report, Summary for Policymakers. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Saqib, M.; Anjum, M.A.; Ali, M.; Ahmad, R.; Sohail, M.; Zakir, I.; Ahmad, S.; Hussain, S. Horticultural Crops as Affected by Climate Change. In Building Climate Resilience in Agriculture: Theory, Practice and Future Perspective; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hashmi, M.Z., Eds.; Springer Nature: Cham, Switzerland, 2022; pp. 95–109. [Google Scholar] [CrossRef]
- Sthapit, S.R.; Scherr, S.J. Tropical fruit trees and opportunities for adaptation and mitigation. In Tropical Fruit Tree Species and Climate Change; Sthapit, B.R., Ramanatha Rao, V., Sthapit, S.R., Eds.; Biodiversity International: New Delhi, India, 2012; pp. 129–137. [Google Scholar]
- Inouye, D.W. Climate change and phenology. WIREs Clim. Chang. 2022, 13, e764. [Google Scholar] [CrossRef]
- IPCC. 2021: Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- Medda, S.; Fadda, A.; Mulas, M. Influence of Climate Change on Metabolism and Biological Characteristics in Perennial Woody Fruit Crops in the Mediterranean Environment. Horticulturae 2022, 8, 273. [Google Scholar] [CrossRef]
- Lu, L.-C.; Chiu, S.-Y.; Chiu, Y.-h.; Chang, T.-H. Sustainability efficiency of climate change and global disasters based on greenhouse gas emissions from the parallel production sectors–A modified dynamic parallel three-stage network DEA model. J. Environ. Manag. 2022, 317, 115401. [Google Scholar] [CrossRef]
- Beck, S.; Mahony, M. The IPCC and the new map of science and politics. Wiley Interdiscip. Rev. Clim. Chang. 2018, 9, e547. [Google Scholar] [CrossRef]
- Rodríguez, A.; Pérez-López, D.; Centeno, A.; Ruiz-Ramos, M. Viability of temperate fruit tree varieties in Spain under climate change according to chilling accumulation. Agric. Syst. 2021, 186, 102961. [Google Scholar] [CrossRef]
- Aslam, M.; Ahmed, M.; Hassan, F.-U.; Afzal, O.; Mehmood, M.Z.; Qadir, G.; Asif, M.; Komal, S.; Hussain, T. Impact of Temperature Fluctuations on Plant Morphological and Physiological Traits. In Building Climate Resilience in Agriculture; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hashmi, M.Z., Eds.; Springer Nature: Cham, Switzerland, 2022; pp. 25–52. [Google Scholar] [CrossRef]
- Rodríguez, A.; Pérez-López, D.; Sánchez, E.; Centeno, A.; Gómara, I.; Dosio, A.; Ruiz-Ramos, M. Chilling accumulation in fruit trees in Spain under climate change. Nat. Hazards Earth Syst. Sci. 2019, 19, 1087–1103. [Google Scholar] [CrossRef]
- Cherif, S.; Doblas-Miranda, E.; Lionello, P.; Borrego, C.; Giorgi, F.; Iglesias, A.; Jebari, S.; Mahmoudi, E.; Moriondo, M.; Pringault, O.; et al. Drivers of Change. In Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future. First Mediterranean Assessment Report; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UNEP/MAP: Marseille, France, 2020; pp. 59–180. [Google Scholar]
- Derbile, E.K.; Kanlisi, S.K.; Dapilah, F. Mapping the vulnerability of indigenous fruit trees to environmental change in the fragile savannah ecological zone of Northern Ghana. Heliyon 2022, 8, e09796. [Google Scholar] [CrossRef]
- Abd El-Rahman, M.M.A. Effect of using bio-stimulants and foliar spraying of anti-stressors for counteract the negative effects of climate changes on growth and fruiting of Balady mandarin trees. SVU-Intern. J. Agric. Sci. 2022, 4, 153–167. [Google Scholar] [CrossRef]
- Vanalli, C.; Casagrandi, R.; Gatto, M.; Bevacqua, D. Shifts in the thermal niche of fruit trees under climate change: The case of peach cultivation in France. Agric. For. Meteorol. 2021, 300, 108327. [Google Scholar] [CrossRef]
- Vujadinović Mandić, M.; Vuković Vimić, A.; Ranković-Vasić, Z.; Đurović, D.; Ćosić, M.; Sotonica, D.; Nikolić, D.; Đurđević, V. Observed Changes in Climate Conditions and Weather-Related Risks in Fruit and Grape Production in Serbia. Atmosphere 2022, 13, 948. [Google Scholar] [CrossRef]
- Stewart, A.L.; Ahmed, S. Chapter 7—Effects of climate change on fruit nutrition. In Fruit Crops; Srivastava, A.K., Hu, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 77–93. [Google Scholar] [CrossRef]
- Paul, V.; Pandey, V.; Singh, A. Vulnerability of trees and fruit crops to climate change. In Climate Change: Impacts and Adaptations in Crop Plants; Singh, M.P., Ed.; Today’s and Tomorrow’s Printer and Publishers: New Delhi, India, 2011. [Google Scholar] [CrossRef]
- Stockle, C.; Marsal, J.; Villar, J.M. Impact of climate change on irrigated tree fruit production. Acta Hortic. 2011, 889, 41–52. [Google Scholar] [CrossRef]
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef]
- Hribar, J.; Vidrih, R. Impacts of climate change on fruit physiology and quality. In Proceedings of the 50th Croatian and 10th International Symposium on Agriculture, Opatija, Croatia, 16–20 February 2015; pp. 42–45. [Google Scholar]
- Tominaga, A.; Ito, A.; Sugiura, T.; Yamane, H. How Is Global Warming Affecting Fruit Tree Blooming? “Flowering (Dormancy) Disorder” in Japanese Pear (Pyrus pyrifolia) as a Case Study. Front. Plant Sci. 2022, 12, 787638. [Google Scholar] [CrossRef]
- Fernandez, E.; Mojahid, H.; Fadón, E.; Rodrigo, J.; Ruiz, D.; Egea, J.A.; Ben Mimoun, M.; Kodad, O.; El Yaacoubi, A.; Ghrab, M.; et al. Climate change impacts on winter chill in Mediterranean temperate fruit orchards. Reg. Environ. Chang. 2022, 23, 7. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.; Luedeling, V.; Girvetz, E.H. Sensitivity of winter chill models for fruit and nut trees to climatic changes expected in California’s Central Valley. Agric. Ecosyst. Environ. 2009, 133, 23–31. [Google Scholar] [CrossRef]
- Fraga, H.; Santos, J.A. Assessment of Climate Change Impacts on Chilling and Forcing for the Main Fresh Fruit Regions in Portugal. Front. Plant Sci. 2021, 12, 689121. [Google Scholar] [CrossRef] [PubMed]
- Drogoudi, P.; Cantín, C.M.; Brandi, F.; Butcaru, A.; Cos-Terrer, J.; Cutuli, M.; Foschi, S.; Galindo, A.; García-Brunton, J.; Luedeling, E.; et al. Impact of Chill and Heat Exposures under Diverse Climatic Conditions on Peach and Nectarine Flowering Phenology. Plants 2023, 12, 584. [Google Scholar] [CrossRef] [PubMed]
- Pantelidis, G.; Drogoudi, P. Exploitation of genotypic variation in chilling and heat requirements for flowering in Prunus armeniaca and Prunus persica (L.) Batsch cultivars. Sci. Hortic. 2023, 321, 112287. [Google Scholar] [CrossRef]
- Naseri, S.; Gholami, M.; Baninasab, B. Chilling and heat requirements in the flower and vegetative buds of some local almond cultivars. Theor. Appl. Clim. 2023, 154, 337–347. [Google Scholar] [CrossRef]
- Alzate-Marin, A.L.; Rivas, P.M.S.; Galaschi-Teixeira, J.S.; Bonifácio-Anacleto, F.; Silva, C.C.; Schuster, I.; Nazareno, A.G.; Giuliatti, S.; da Rocha Filho, L.C.; Garófalo, C.A.; et al. Warming and elevated CO2 induces changes in the reproductive dynamics of a tropical plant species. Sci. Total Environ. 2021, 768, 144899. [Google Scholar] [CrossRef] [PubMed]
- Nath, V.; Kumar, G.; Pandey, S.; Pandey, S. Impact of climate change on tropical fruit production systems and its mitigation strategies. In Climate Change and Agriculture in India: Impact and Adaptation; Mahdi, S.S., Ed.; Springer Nature: Cham, Switzerland, 2019; pp. 129–146. [Google Scholar]
- Dias, M.C.; Araújo, M.; Silva, S.; Santos, C. Sustainable Olive Culture under Climate Change: The Potential of Biostimulants. Horticulturae 2022, 8, 1048. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Damvakaris, T.; Roussos, P.A. Antioxidant defense system in young olive plants against drought stress and mitigation of adverse effects through external application of alleviating products. Sci. Hortic. 2020, 259, 108812. [Google Scholar] [CrossRef]
- Chmielewski, F.-M.; Müller, A.; Bruns, E. Climate changes and trends in phenology of fruit trees and field crops in Germany, 1961–2000. Agric. For. Meteorol. 2004, 121, 69–78. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- El Yaacoubi, A.; Malagi, G.; Oukabli, A.; Hafidi, M.; Legave, J.-M. Global warming impact on floral phenology of fruit trees species in Mediterranean region. Sci. Hortic. 2014, 180, 243–253. [Google Scholar] [CrossRef]
- Lopez, G.; Mata, M.; Arbones, A.; Solans, J.R.; Girona, J.; Marsal, J. Mitigation of effects of extreme drought during stage III of peach fruit development by summer pruning and fruit thinning. Tree Physiol. 2006, 26, 469–477. [Google Scholar] [CrossRef]
- Toreti, A.; Deryng, D.; Tubiello, F.N.; Müller, C.; Kimball, B.A.; Moser, G.; Boote, K.; Asseng, S.; Pugh, T.A.M.; Vanuytrecht, E.; et al. Narrowing uncertainties in the effects of elevated CO2 on crops. Nat. Food 2020, 1, 775–782. [Google Scholar] [CrossRef]
- Alae-Carew, C.; Nicoleau, S.; Bird, F.A.; Hawkins, P.; Tuomisto, H.L.; Haines, A.; Dangour, A.D.; Scheelbeek, P.F.D. The impact of environmental changes on the yield and nutritional quality of fruits, nuts and seeds: A systematic review. Environ. Res. Lett. 2020, 15, 023002. [Google Scholar] [CrossRef]
- Fischer, G.; Melgarejo, L.M.; Balaguera-López, H.E. Review on the impact of elevated CO2 concentrations on fruit species in the face of climate change. Cienc. Tecnol. Agropecu. 2022, 23, e2475. [Google Scholar]
- Lauriks, F.; Salomón, R.L.; Steppe, K. Temporal variability in tree responses to elevated atmospheric CO2. Plant Cell Environ. 2021, 44, 1292–1310. [Google Scholar] [CrossRef]
- Zhu, C.; Wolf, J.; Zhang, J.; Anderegg, W.R.L.; Bunce, J.A.; Ziska, L.H. Rising temperatures can negate CO2 fertilization effects on global staple crop yields: A meta-regression analysis. Agric. For. Meteorol. 2023, 342, 109737. [Google Scholar] [CrossRef]
- Ma, Y.; Xie, Y.; Ha, R.; Cao, B.; Song, L. Effects of Elevated CO2 on Photosynthetic Accumulation, Sucrose Metabolism-Related Enzymes, and Genes Identification in Goji Berry (Lycium barbarum L.). Front. Plant Sci. 2021, 12, 643555. [Google Scholar] [CrossRef]
- Kimball, B.A.; Idso, S.B.; Johnson, S.; Rillig, M.C. Seventeen years of carbon dioxide enrichment of sour orange trees: Final results. Glob. Chang. Biol. 2007, 13, 2171–2183. [Google Scholar] [CrossRef]
- Pritchard, S.G.; Rogers, H.H.; Prior, S.A.; Peterson, C.M. Elevated CO2 and plant structure: A review. Glob. Chang. Biol. 1999, 5, 807–837. [Google Scholar] [CrossRef]
- Yong, J.W.; Wong, S.C.; Letham, D.S.; Hocart, C.H.; Farquhar, G.D. Effects of elevated CO2 and nitrogen nutrition on cytokinins in the xylem sap and leaves of cotton. Plant Physiol. 2000, 124, 767–780. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Asensio, J.S.R.; Cousins, A.B. Carbon Dioxide Enrichment Inhibits Nitrate Assimilation in Wheat and ‘Arabidopsis’. Science 2010, 328, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Maschler, J.; Bialic-Murphy, L.; Wan, J.; Andresen, L.C.; Zohner, C.M.; Reich, P.B.; Lüscher, A.; Schneider, M.K.; Müller, C.; Moser, G.; et al. Links across ecological scales: Plant biomass responses to elevated CO2. Glob. Chang. Biol. 2022, 28, 6115–6134. [Google Scholar] [CrossRef]
- Wang, F.; Gao, J.; Yong, J.W.H.; Wang, Q.; Ma, J.; He, X. Higher Atmospheric CO2 Levels Favor C3 Plants Over C4 Plants in Utilizing Ammonium as a Nitrogen Source. Front. Plant Sci. 2020, 11, 537443. [Google Scholar] [CrossRef]
- Ramírez, F.; Kallarackal, J. Response of trees to CO2 Increase. In Responses of Fruit Trees to Global Climate Change; Springer International Publishing: Cham, Switzerland, 2015; pp. 3–7. [Google Scholar] [CrossRef]
- Hao, L.; Chang, Z.; Lu, Y.; Tian, Y.; Zhou, H.; Wang, Y.; Liu, L.; Wang, P.; Zheng, Y.; Wu, J. Drought dampens the positive acclimation responses of leaf photosynthesis to elevated CO2 by altering stomatal traits, leaf anatomy, and Rubisco gene expression in Pyrus. Environ. Exp. Bot. 2023, 211, 105375. [Google Scholar] [CrossRef]
- Yu, M.; Sun, P.; Huang, X.; Zha, Z.; Wang, X.; Mantri, N.; Lou, H.; Jiang, B.; Shen, Z.; Sun, Y.; et al. Interacting Effects of CO2, Temperature, and Nitrogen Supply on Photosynthetic, Root Growth, and Nitrogen Allocation of Strawberry at the Fruiting Stage. Agronomy 2023, 13, 1353. [Google Scholar] [CrossRef]
- Leibar-Porcel, E.; Dodd, I.C. Role of Plant Hormones in Plant Response to Elevated CO2 Concentrations: Above- and Below-ground Interactions. In Plant Hormones and Climate Change; Ahammed, G.J., Yu, J., Eds.; Springer Nature: Singapore, 2023; pp. 55–74. [Google Scholar] [CrossRef]
- Norby, R.J.; O’Neill, E.G.; Luxmoore, R.J. Effects of Atmospheric CO2 Enrichment on the Growth and Mineral Nutrition of Quercus alba Seedlings in Nutrient-Poor Soil. Plant Physiol. 1986, 82, 83–89. [Google Scholar] [CrossRef]
- Conroy, J.P.; Milham, P.J.; Mazur, M.; Barlow, E.W.R. Growth, dry weight partitioning and wood properties of Pinus radiata D. Don after 2 years of CO2 enrichment. Plant Cell Environ. 1990, 13, 329–337. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Wei, Z.; Liu, F. ABA-mediated modulation of elevated CO2 on stomatal response to drought. Curr. Opin. Plant Biol. 2020, 56, 174–180. [Google Scholar] [CrossRef]
- Kiba, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H. Sugar-induced de novo cytokinin biosynthesis contributes to Arabidopsis growth under elevated CO2. Sci. Rep. 2019, 9, 7765. [Google Scholar] [CrossRef]
- Cruz, J.L.; Alves, A.A.C.; LeCain, D.R.; Ellis, D.D.; Morgan, J.A. Interactive effects between nitrogen fertilization and elevated CO2 on growth and gas exchange of papaya seedlings. Sci. Hortic. 2016, 202, 32–40. [Google Scholar] [CrossRef]
- Elbasiouny, H.; El-Ramady, H.; Elbehiry, F.; Rajput, V.D.; Minkina, T.; Mandzhieva, S. Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability 2022, 14, 914. [Google Scholar] [CrossRef]
- Lahive, F.; Handley, L.R.; Hadley, P.; Daymond, A.J. Climate Change Impacts on Cacao: Genotypic Variation in Responses of Mature Cacao to Elevated CO2 and Water Deficit. Agronomy 2021, 11, 818. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Ruan, Y.; Kuzyakov, Y.; Liu, X.; Zhang, X.; Xu, Q.; Guo, J.; Guo, S.; Shen, Q.; Yang, Y.; Ling, N. Elevated temperature and CO2 strongly affect the growth strategies of soil bacteria. Nat. Comm. 2023, 14, 391. [Google Scholar] [CrossRef]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.; Shew, H.D.; Rufty, T.W.; Hu, S. Arbuscular Mycorrhizal Fungi Increase Organic Carbon Decomposition Under Elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef]
- Ammar, A.; Aissa, I.B.; Gouiaa, M.; Mars, M. Fig (Ficus carica L.) vulnerability to climate change: Combined effects of water stress and high temperature on ecophysiological behaviour of different cultivars. S. Afric J. Bot. 2022, 147, 482–492. [Google Scholar] [CrossRef]
- Choudhury, B.U.; Nengzouzam, G.; Islam, A. Evaluation of climate change impact on soil erosion in the integrated farming system based hilly micro-watersheds using Revised Universal Soil Loss Equation. Catena 2022, 214, 106306. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L. Analysis of Invasive Insects: Links to Climate Change. In Invasive Species and Global Climate Change; Ziska, L.H., Dukes, J.S., Eds.; CABI Publishing: Wallingford, UK, 2014; pp. 45–61. ISBN 978-1780641645. [Google Scholar] [CrossRef]
- Jawo, T.O.; Kyereh, D.; Lojka, B. The impact of climate change on coffee production of small farmers and their adaptation strategies: A review. Clim. Dev. 2023, 15, 93–109. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Graniti, A.; Faedda, R.; Cacciola, S.O.; Magnano di San Lio, G. Olive diseases in a changing ecosystem. In Olive Diseases and Disorders; Transworld Research Network: Kerala, India, 2011; pp. 403–433. [Google Scholar]
- Narouei-Khandan, H.A.; Worner, S.P.; Viljanen, S.L.H.; van Bruggen, A.H.C.; Balestra, G.M.; Jones, E. The Potential Global Climate Suitability of Kiwifruit Bacterial Canker Disease (Pseudomonas syringae pv. actinidiae (Psa)) Using Three Modelling Approaches: CLIMEX, Maxent and Multimodel Framework. Climate 2022, 10, 14. [Google Scholar] [CrossRef]
- Nunes, L.J. Creation of Value Chains for the Sustainability of Control and Eradication Actions on Ailanthus altissima (Mill.) Swingle. Environments 2022, 9, 64. [Google Scholar] [CrossRef]
- Chuang, L.; Liu, S.; Biedermann, D.; Franke, J. Identification of early quassinoid biosynthesis in the invasive tree of heaven (Ailanthus altissima) confirms evolutionary origin from protolimonoids. Front. Plant Sci. 2022, 13, 958138. [Google Scholar] [CrossRef]
- Valiño, A.; Pardo-Muras, M.; López-Periago, J.E.; Puig, C.G.; Pedrol, N. Biomass from Allelopathic Agroforestry and Invasive Plant Species as Soil Amendments for Weed Control—A Review. Agronomy 2023, 13, 2880. [Google Scholar] [CrossRef]
- Khamare, Y.; Chen, J.; Marble, S.C. Allelopathy and its application as a weed management tool: A review. Front. Plant Sci. 2022, 13, 1034649. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef]
- Korres, N.E.; Norsworthy, J.K.; Tehranchian, P.; Gitsopoulos, T.K.; Loka, D.A.; Oosterhuis, D.M.; Gealy, D.R.; Moss, S.R.; Burgos, N.R.; Miller, M.R.; et al. Cultivars to face climate change effects on crops and weeds: A review. Agron. Sustain. Dev. 2016, 36, 12. [Google Scholar] [CrossRef]
- Tataridas, A.; Moreira, M.; Frazão, L.; Kanatas, P.; Ota, N.; Travlos, I. Biology of Invasive Plants 5. Solanum elaeagnifolium Cav. Invasive Plant Sci. Manag. 2023, 16, 139–159. [Google Scholar] [CrossRef]
- Ramesh, K.; Matloob, A.; Aslam, F.; Florentine, S.K.; Chauhan, B.S. Weeds in a Changing Climate: Vulnerabilities, Consequences, and Implications for Future Weed Management. Front. Plant Sci. 2017, 8, 95. [Google Scholar] [CrossRef]
- Zeinab, R.-R.; Majid, M.; Ahmad, G.; Taqi, R.; Neal, S.E. Abiotic Stresses Management in Citrus. In Citrus Research; Mateus Perei-ra, G., Júlia Scherer, S., Eds.; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar] [CrossRef]
- Ziogas, V.; Tanou, G.; Morianou, G.; Kourgialas, N. Drought and Salinity in Citriculture: Optimal Practices to Alleviate Salinity and Water Stress. Agronomy 2021, 11, 1283. [Google Scholar] [CrossRef]
- Jalil, S.U.; Ansari, M.I. Biotechnology Strategies to Combat Plant Abiotic Stress. In Nanobiotechnology: Mitigation of Abiotic Stress in Plants; Al-Khayri, J.M., Ansari, M.I., Singh, A.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 61–76. [Google Scholar] [CrossRef]
- Kisku, K.; Naik, U.C. Nanobiotechnology: A Process to Combat Abiotic Stress in Crop Plants. In Nanobiotechnology: Mitigation of Abiotic Stress in Plants; Al-Khayri, J.M., Ansari, M.I., Singh, A.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 139–163. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Hernández, J.A.; Díaz-Vivancos, P. Antioxidant system: The hub of bud dormancy regulation in Prunus sp. Sci. Hortic. 2022, 305, 111396. [Google Scholar] [CrossRef]
- Bound, S.A.; Foo, E.; Gélinas-Marion, A.; Nichols, D.S.; Nissen, R. The impact of dormancy breakers on hormone profiles, Fruit Growth and Quality in Sweet Cherry. Agriculture 2022, 12, 270. [Google Scholar] [CrossRef]
- Yamane, H.; Singh, A.K.; Cooke, J.E.K. Plant dormancy research: From environmental control to molecular regulatory networks. Tree Physiol. 2021, 41, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Quesada-Traver, C.; Ríos, G. Models for a molecular calendar of bud-break in fruit trees. Sci. Hortic. 2022, 297, 110972. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Díaz-Vivancos, P.; Martínez-Sánchez, G.; Alburquerque, N.; Martínez, D.; Barba-Espín, G.; Acosta-Motos, J.R.; Carrera, E.; García-Bruntón, J. Physiological and biochemical characterization of bud dormancy: Evolution of carbohydrate and antioxidant metabolisms and hormonal profile in a low chill peach variety. Sci. Hortic. 2021, 281, 109957. [Google Scholar] [CrossRef]
- Wang, S.Y.; Steffens, G.L.; Faust, M. Breaking bud dormancy in apple with a plant bioregulator, thidiazuron. Phytochemistry 1986, 25, 311–317. [Google Scholar] [CrossRef]
- Bera, K.; Dutta, P.; Sadhukhan, S. Plant Responses Under Abiotic Stress and Mitigation Options Towards Agricultural Sustainability. In Plant Stress: Challenges and Management in the New Decade; Roy, S., Mathur, P., Chakraborty, A.P., Saha, S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 3–28. [Google Scholar] [CrossRef]
- Ntanos, E.; Tsafouros, A.; Denaxa, N.-K.; Kosta, A.; Bouchagier, P.; Roussos, P.A. Mitigation of high solar irradiance and heat stress in kiwifruit during summer via the use of alleviating products with different modes of action—Part 1 Effects on leaf physiology and biochemistry. Agriculture 2022, 12, 2121. [Google Scholar] [CrossRef]
- Khattak, W.; Sun, J.; Abbas, A.; Hameed, R.; Jalal, D.; Niaz, N.; Anwar, S.; Liu, Y.; Wang, Y. Melatonin alleviating drought stress in plants: A review. S. Afr. J. Bot. 2023, 161, 192–201. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Naga, K.C.; Kumar, R.; Chourasia, K.N.; Subhash, S.; Kumar, D.; Sharma, S. Emerging roles of melatonin in mit-igating abiotic and biotic stresses of horticultural crops. Sci. Hortic. 2020, 272, 109592. [Google Scholar] [CrossRef]
- Sajjad, N.; Bhat, E.A.; Hassan, S.; Ali, R.; Shah, D. Role of Melatonin in Amelioration of Abiotic Stress-induced Damages. In Pro-Tective Chemical Agents in the Amelioration of Plant Abiotic Stress. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 306–317. [Google Scholar] [CrossRef]
- Parankusam, S.; Jalli, L.; Bhatnagar-Mathur, P.; Sharma, K. Emerging Roles of Salicylic Acid and Jasmonates in Plant Abiotic Stress Responses. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 342–373. [Google Scholar] [CrossRef]
- Zahid, G.; Iftikhar, S.; Shimira, F.; Ahmad, H.M.; Aka Kaçar, Y. An overview and recent progress of plant growth regulators (PGRs) in the mitigation of abiotic stresses in fruits: A review. Sci. Hortic. 2023, 309, 111621. [Google Scholar] [CrossRef]
- Vardhini, B. Brassinosteroids and Salicylic Acid as Chemical Agents to Ameliorate Diverse Environmental Stresses in Plants. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 389–412. [Google Scholar] [CrossRef]
- Ranjan, A.; Sinha, R.; Bala, M.; Pareek, A.; Singla-Pareek, S.L.; Singh, A.K. Silicon-mediated abiotic and biotic stress mitigation in plants: Underlying mechanisms and potential for stress resilient agriculture. Plant Physiol. Biochem. 2021, 163, 15–25. [Google Scholar] [CrossRef]
- Ahmad, F.; Singh, A.; Kamal, A. Osmoprotective Role of Sugar in Mitigating Abiotic Stress in Plants. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 53–70. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Kundu, A. Sugars and Sugar Polyols in Overcoming Environmental Stresses. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 71–101. [Google Scholar] [CrossRef]
- Graziani, G.; Cirillo, A.; Giannini, P.; Conti, S.; El-Nakhel, C.; Rouphael, Y.; Ritieni, A.; Di Vaio, C. Biostimulants improve plant growth and bioactive compounds of young olive trees under abiotic stress conditions. Agriculture 2022, 12, 227. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Roussos, P.A.; Damvakaris, T.; Stournaras, V. Comparative effects of exogenous glycine betaine, kaolin clay particles and Ambiol on photosynthesis, leaf sclerophylly indexes and heat load of olive cv. Chondrolia Chalkidikis under drought. Sci. Hortic. 2012, 137, 87–94. [Google Scholar] [CrossRef]
- Abobatta, w. Fruit orchards under climate change conditions. J. Appl. Bio. Bioeng. 2021, 8, 99–102. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Tsafouros, A.; Ntanos, E.; Roussos, P.A. Chapter 8—Role of glycine betaine in the protection of plants against environmental stresses. In Plant Stress Mitigators; Ghorbanpour, M., Adnan Shahid, M., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 127–158. [Google Scholar] [CrossRef]
- Bhatt, M.D.; Bhatt, D. Strigolactones in Overcoming Environmental Stresses. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 327–341. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, C.-B.; Ren, R.-M.; Jiang, J.-H. Salicylic acid had the potential to enhance tolerance in horticultural crops against abiotic stress. Front. Plant Sci. 2023, 14, 1141918. [Google Scholar] [CrossRef]
- Khalid, M.F.; Saleem, M.S.; Zakir, I.; Khan, R.I.; Sohail, M.; Ejaz, S.; Anjum, M.A.; Sabir, S.; Ali, S.; Ahmad, S.; et al. Chapter 3—Salicylic acid induced abiotic stress tolerance in plants. In Plant Stress Mitigators; Ghorbanpour, M., Adnan Shahid, M., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 57–67. [Google Scholar] [CrossRef]
- Reddy, K.S.; Wakchaure, G.; Khapte, P.; Changan, S. Plant Bio-stimulants for Mitigating Abiotic Stresses in Agriculture. Indian J. Fertil. 2023, 19, 788–800. [Google Scholar]
- Biondi, S.; Antognoni, F.; Marincich, L.; Mariacaterina, L.; Tejos, R.; Ruiz Carrasco, K. The polyamine “multiverse” and stress mit-igation in crops: A case study with seed priming in quinoa. Sci. Hortic. 2022, 304, 111292. [Google Scholar] [CrossRef]
- Sajjad, N.; Bhat, E.A.; Shah, D.; Wani, A.; Nazir, N.; Ali, R.; Hassan, S. Abscisic Acid Application and Abiotic Stress Ameliora-tion. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 280–290. [Google Scholar] [CrossRef]
- Munir, N.; Hanif, M.; Abideen, Z.; Sohail, M.; El-Keblawy, A.; Radicetti, E.; Mancinelli, R.; Haider, G. Mechanisms and strategies of plant microbiome interactions to mitigate abiotic stresses. Agronomy 2022, 12, 2069. [Google Scholar] [CrossRef]
- Singh, A.; Mazahar, S.; Chapadgaonkar, S.S.; Giri, P.; Shourie, A. Phyto-microbiome to mitigate abiotic stress in crop plants. Front. Microbiol. 2023, 14, 1210890. [Google Scholar] [CrossRef] [PubMed]
- Imran, S.; Sarker, P.; Hoque, M.N.; Paul, N.C.; Mahamud, M.A.; Chakrobortty, J.; Tahjib-Ul-Arif, M.; Latef, A.A.H.A.; Hasanuz-zaman, M.; Rhaman, M.S. Biochar actions for the mitigation of plant abiotic stress. Crop Pasture Sci. 2023, 74, 6–20. [Google Scholar] [CrossRef]
- Huang, L.; Fu, W.; Zhang, Y.; Liu, X.; Wang, Q.; Wang, L.; Tanveer, M. The role of melatonin in regulating horticultural crop production under various abiotic stresses. Sci. Hortic. 2024, 323, 112508. [Google Scholar] [CrossRef]
- Koh, Y.S.; Wong, S.K.; Ismail, N.H.; Zengin, G.; Duangjai, A.; Saokaew, S.; Phisalprapa, P.; Tan, K.W.; Goh, B.H.; Tang, S.Y. Mit-igation of Environmental Stress-Impacts in Plants: Role of Sole and Combinatory Exogenous Application of Glutathione. Front. Plant Sci. 2021, 12, 791205. [Google Scholar] [CrossRef] [PubMed]
- Das, K. Ascorbate and Tocopherols in Mitigating Oxidative Stress. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 102–121. [Google Scholar] [CrossRef]
- Amist, N.; Singh, N.B. Role of Glutathione Application in Overcoming Environmental Stress. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 122–146. [Google Scholar] [CrossRef]
- Machado, R.; Serralheiro, R. Chapter 33—Salt stress alleviation through fertilization in fruit crops. In Fruit Crops; Srivastava, A.K., Hu, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 465–480. [Google Scholar] [CrossRef]
- Ali, R.; Hassan, S.; Shah, D.; Sajjad, N.; Bhat, E.A. Role of Polyamines in Mitigating Abiotic Stress. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, K., Tripathi, D.K., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 291–305. [Google Scholar] [CrossRef]
- Liu, J.; Qiu, G.; Liu, C.; Li, H.; Chen, X.; Fu, Q.; Lin, Y.; Guo, B. Salicylic Acid, a multifaceted hormone, combats abiotic stresses in plants. Life 2022, 12, 886. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weather Event | Impacts on Various Fruit/Nut Developmental Stages |
|---|---|
| Mild winter |
|
| Falsh spring conditions |
|
| Extremely high and long-lasting temperatures in summer |
|
| Low precipitation |
|
| Water stress |
|
| Warm autumn and early winter |
|
| Water logging |
|
| Frost |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roussos, P.A. Climate Change Challenges in Temperate and Sub-Tropical Fruit Tree Cultivation. Encyclopedia 2024, 4, 558-582. https://doi.org/10.3390/encyclopedia4010036
Roussos PA. Climate Change Challenges in Temperate and Sub-Tropical Fruit Tree Cultivation. Encyclopedia. 2024; 4(1):558-582. https://doi.org/10.3390/encyclopedia4010036
Chicago/Turabian StyleRoussos, Petros A. 2024. "Climate Change Challenges in Temperate and Sub-Tropical Fruit Tree Cultivation" Encyclopedia 4, no. 1: 558-582. https://doi.org/10.3390/encyclopedia4010036
APA StyleRoussos, P. A. (2024). Climate Change Challenges in Temperate and Sub-Tropical Fruit Tree Cultivation. Encyclopedia, 4(1), 558-582. https://doi.org/10.3390/encyclopedia4010036