
Global Chemical Characterization of Sargassum spp. Seaweeds from Different Locations on Caribbean Islands: A Screening of Organic Compounds and Heavy Metals Contents

 , , , ,
, , , ,  , , , and
, , , and
Abstract
1. Introduction
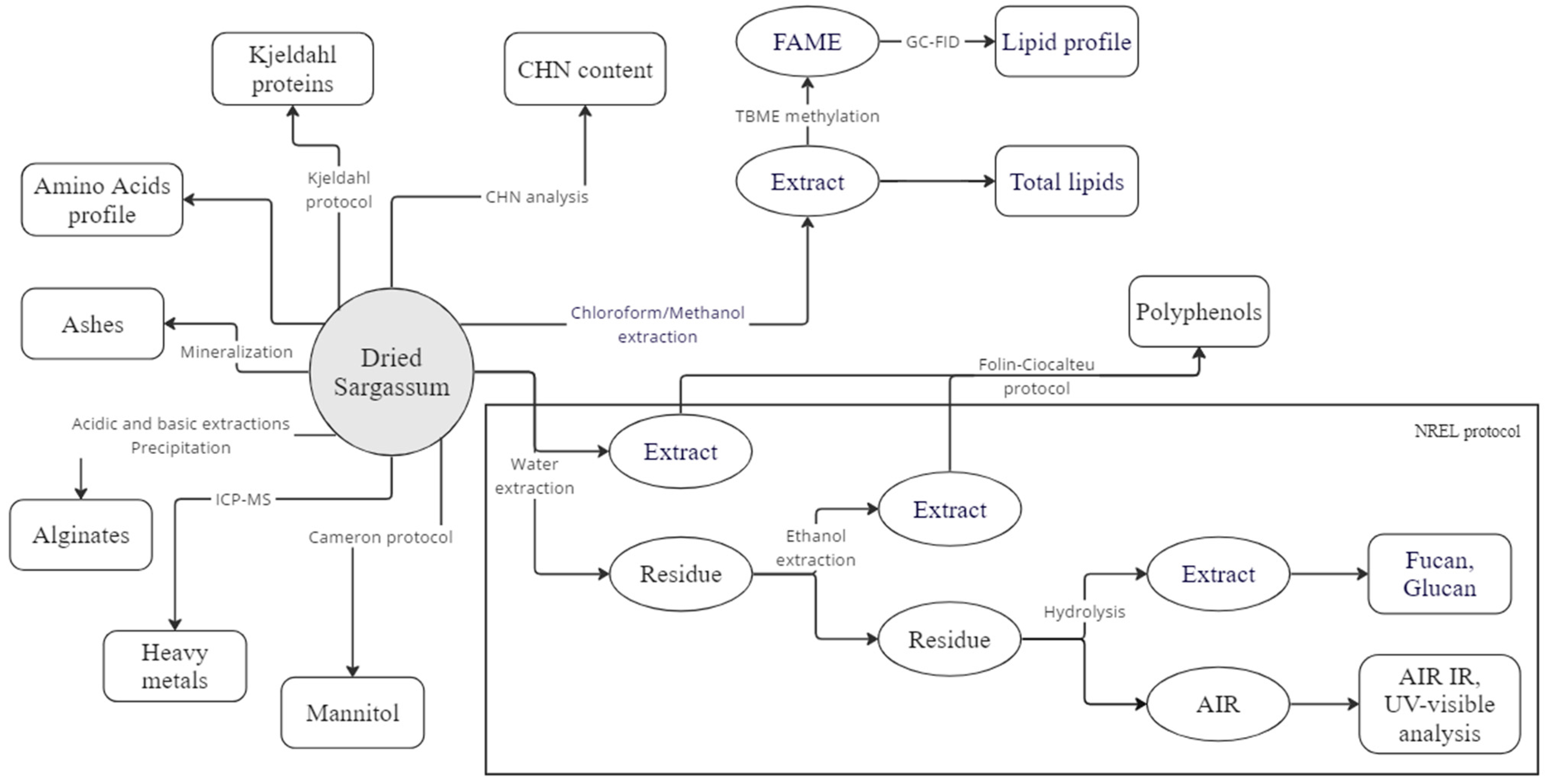
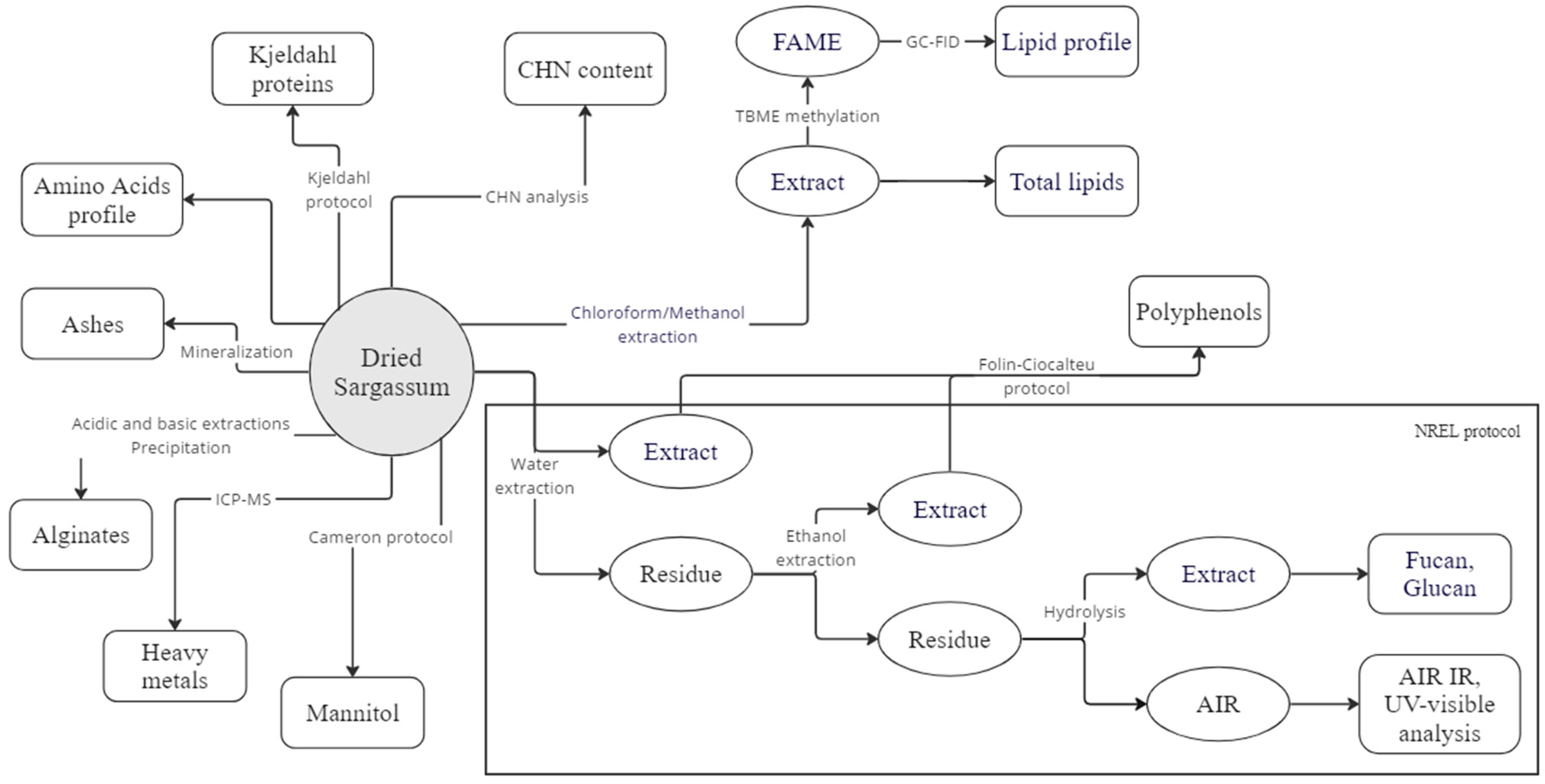
2. Materials and Methods
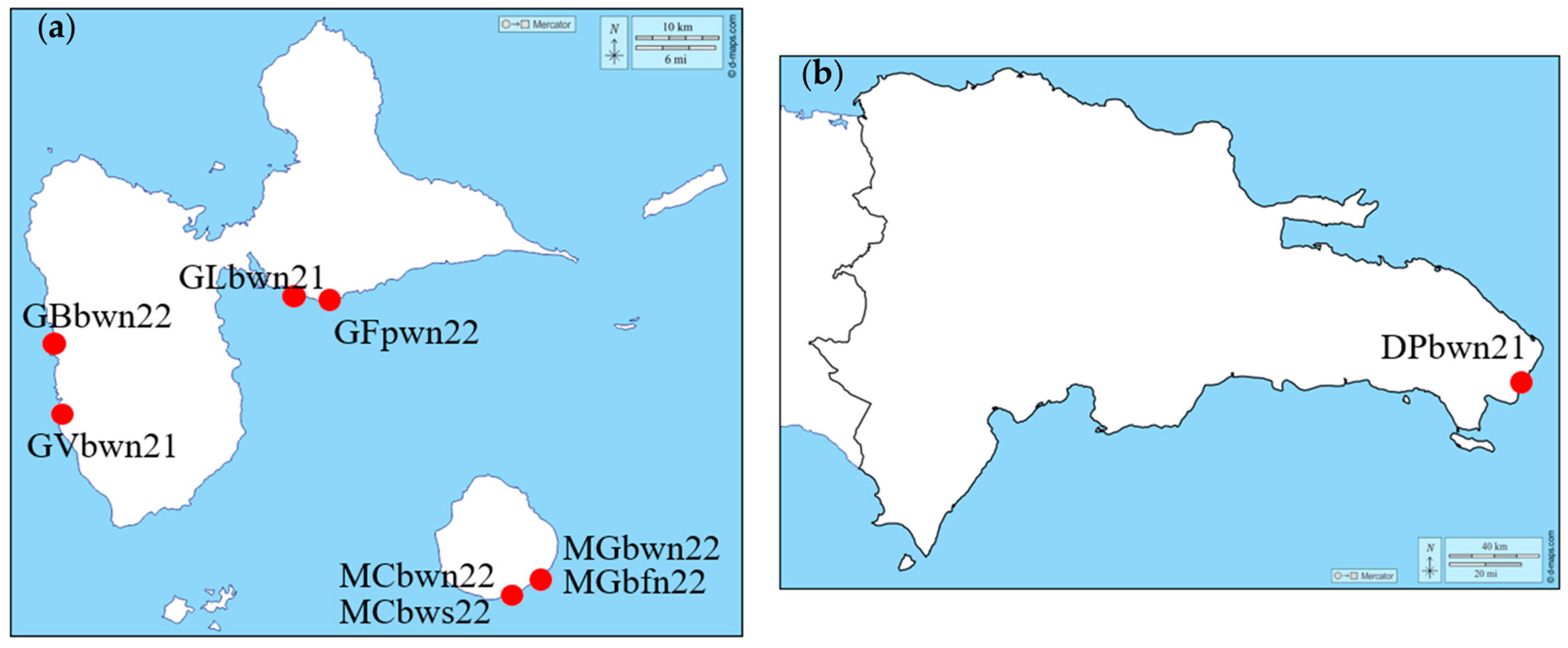
2.1. Sargassum Sampling
2.2. Grinding/Screening
2.3. Moisture
2.4. Ash
2.5. Carbon, Hydrogen and Nitrogen (CHN)Analysis
2.6. Proteins Analysis
2.6.1. Total Protein Content
2.6.2. Amino Acids
2.7. Lipids Analysis
2.7.1. Total Lipid Content
2.7.2. Fatty Acids Profile
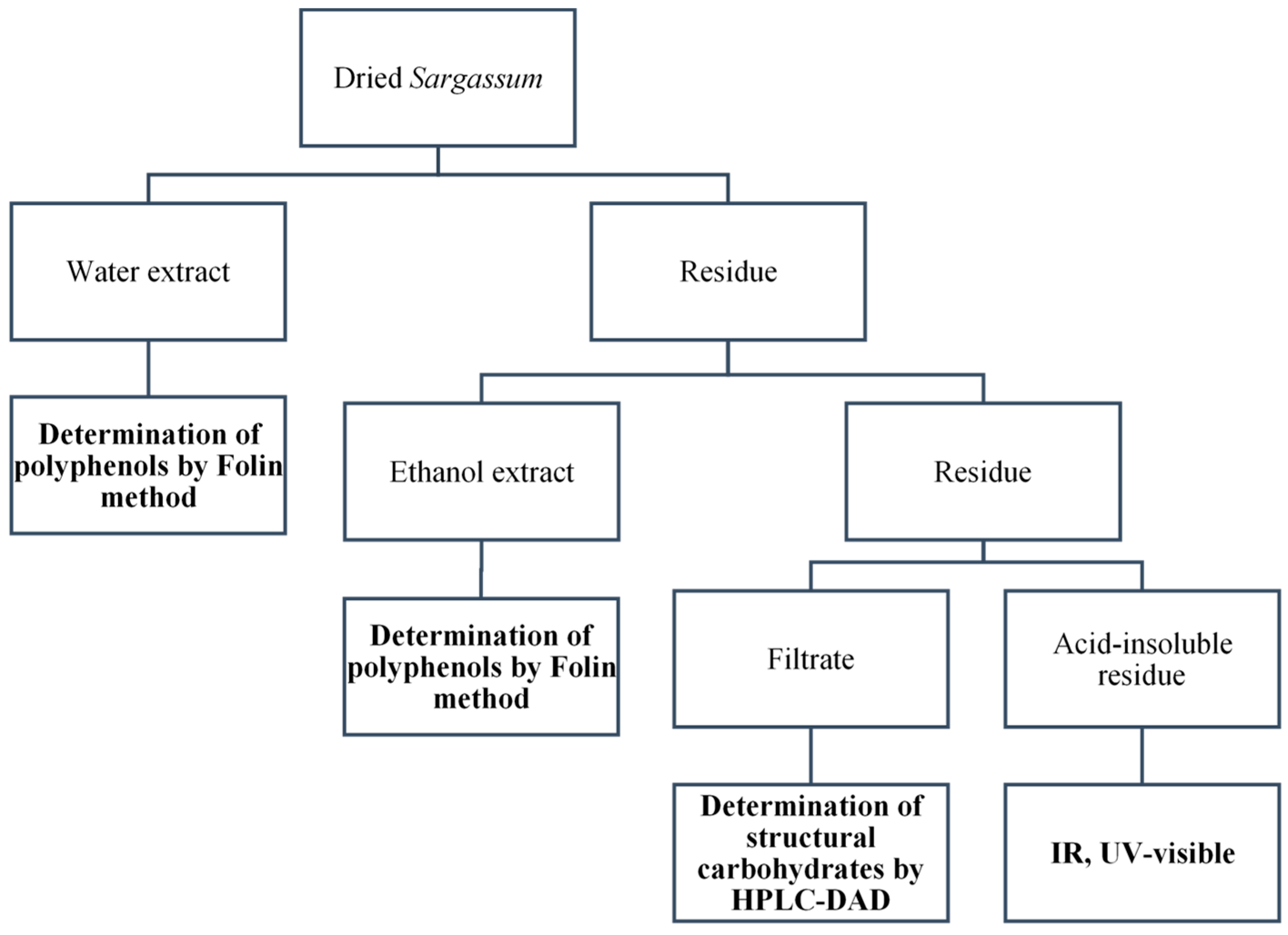
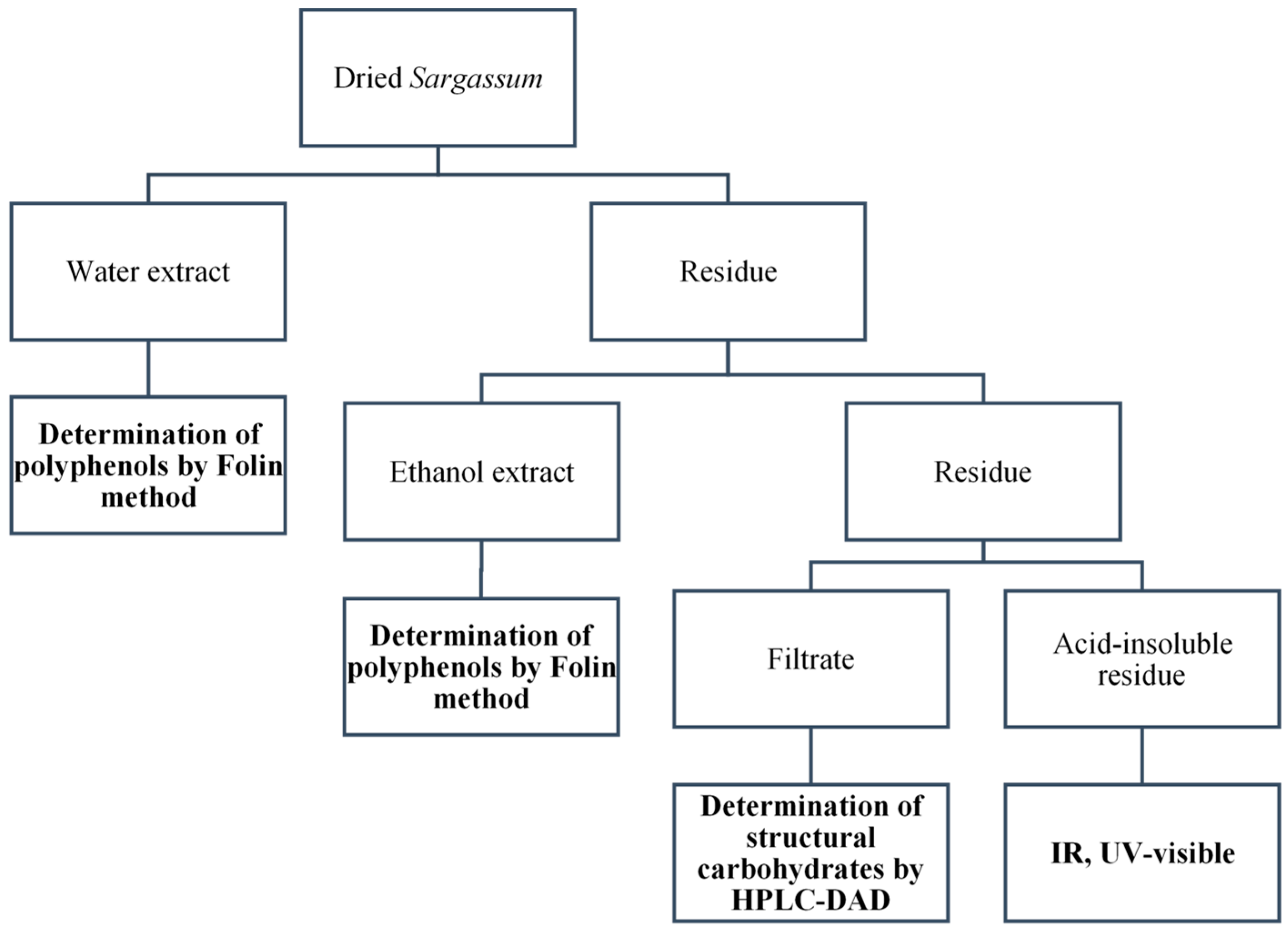
2.8. Structural Carbohydrates
2.8.1. Glucan, Fucan
2.8.2. Spectroscopic Analysis of the Acid-Insoluble Residue
2.8.3. Alginates
2.9. Mannitol
2.10. Polyphenols
2.11. Elemental Analysis
2.12. Statistical Analysis
3. Results
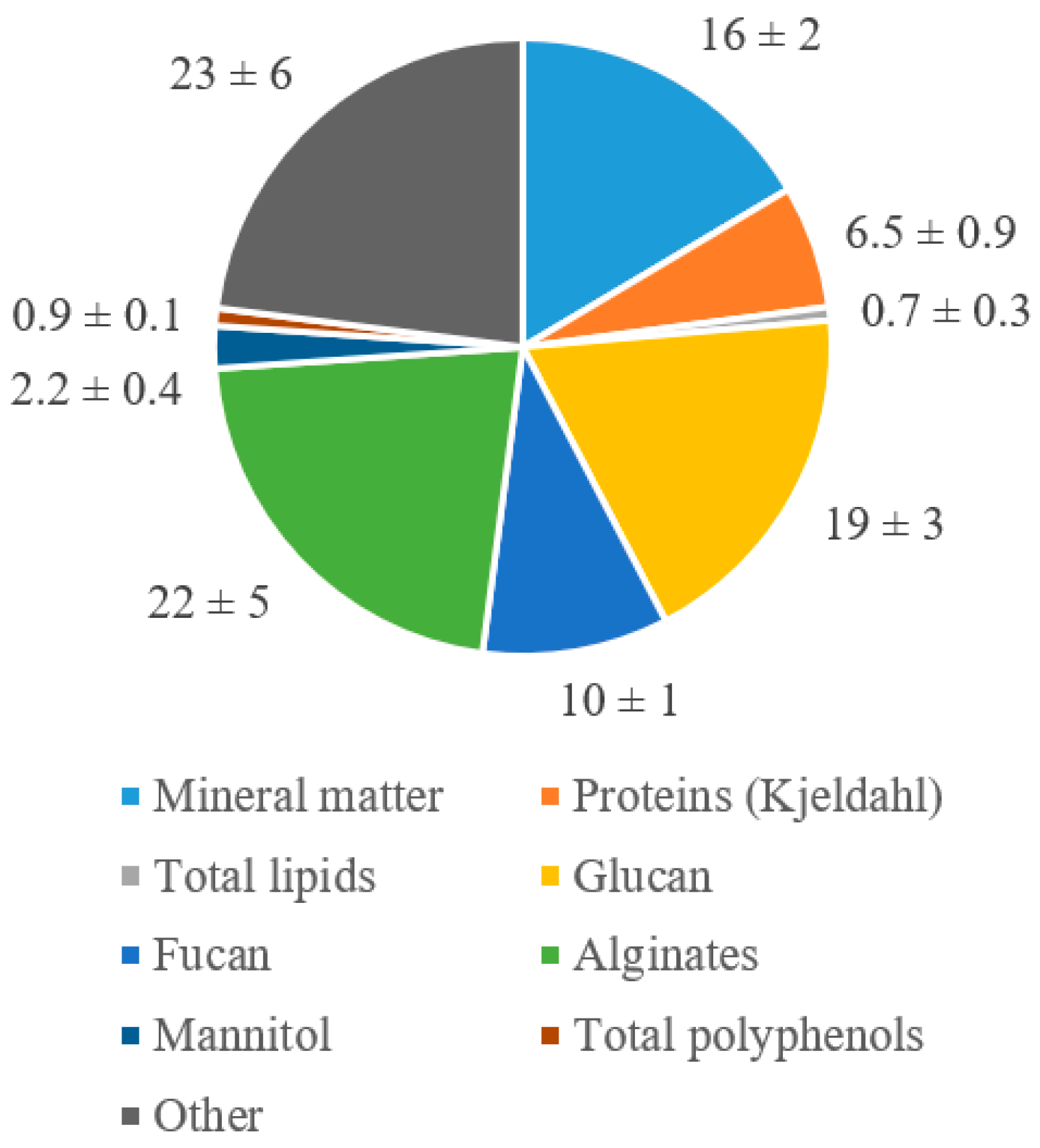
3.1. Proximate Analysis
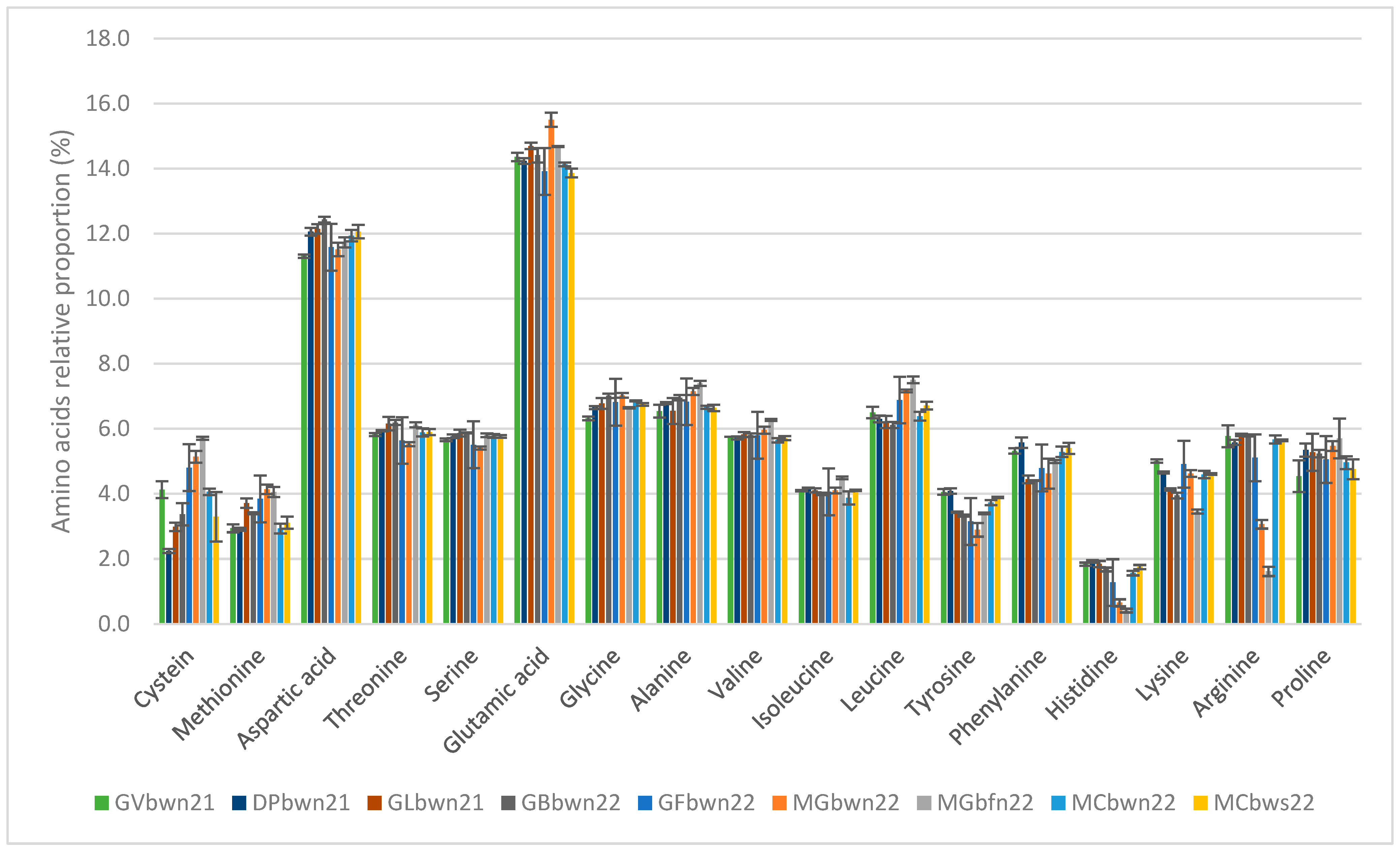
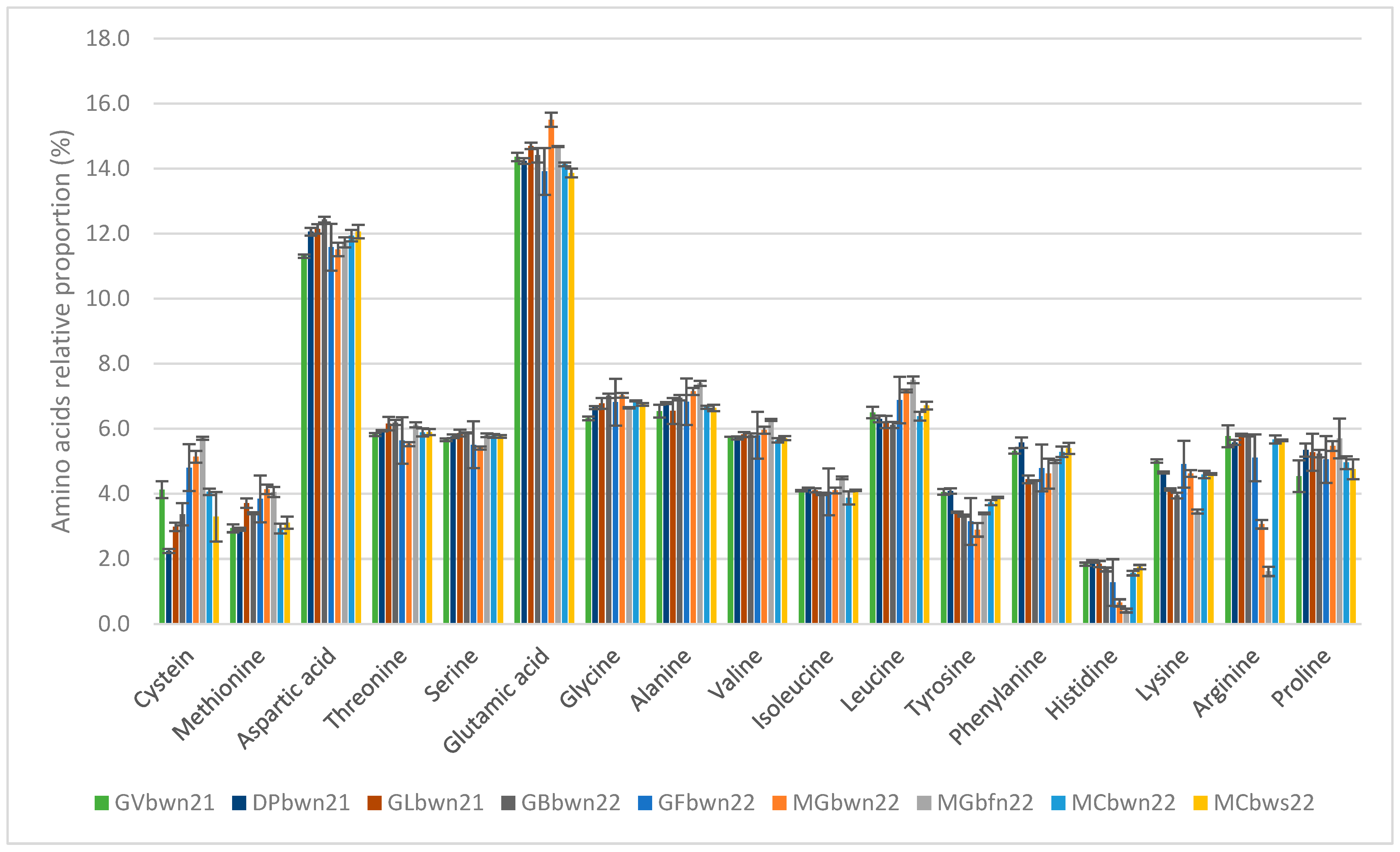
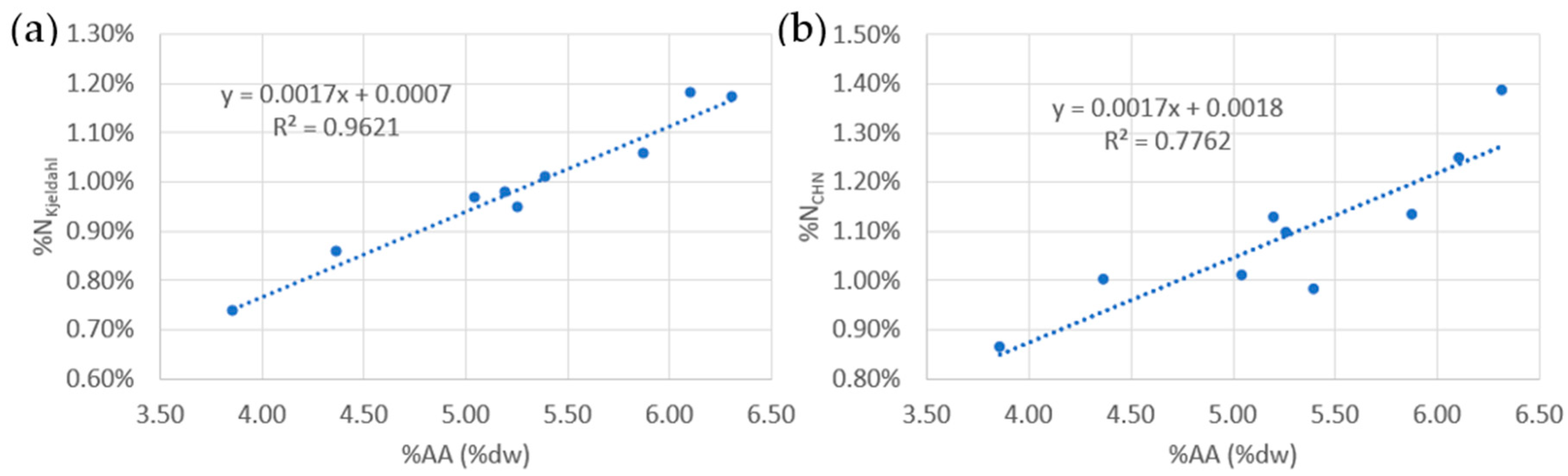
3.2. Proteins
3.3. Lipids
3.4. Structural Carbohydrates
3.5. Mannitol
3.6. Polyphenols
3.7. Elemental Analysis
4. Discussion
4.1. Proximate Analysis
4.2. Protein
4.3. Lipids
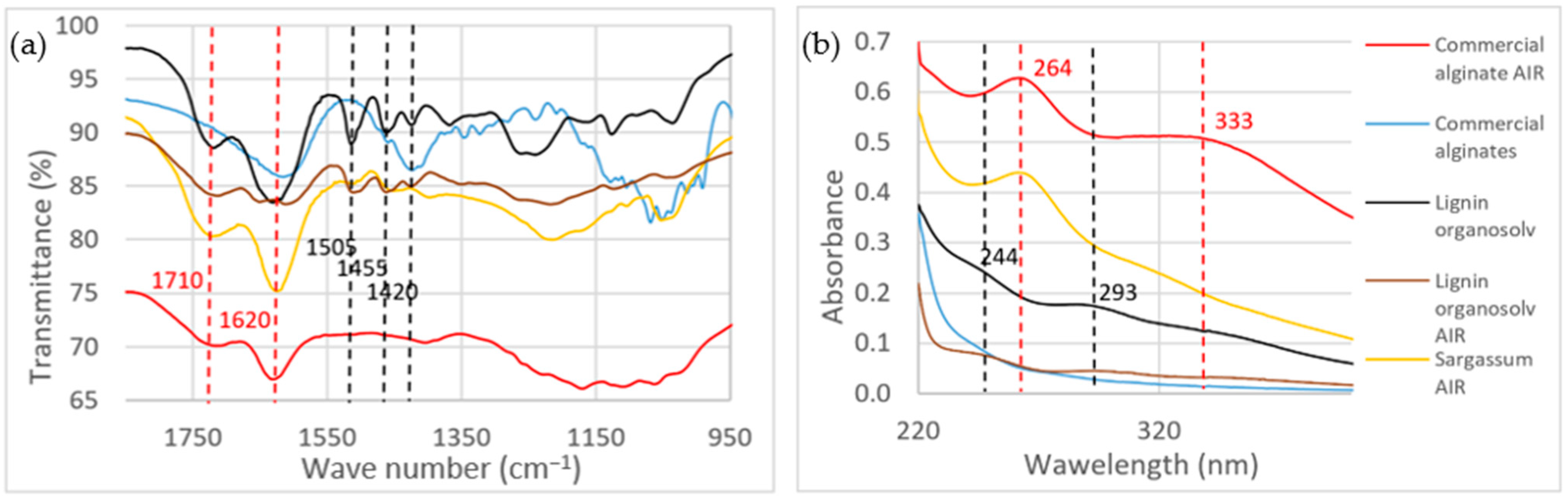
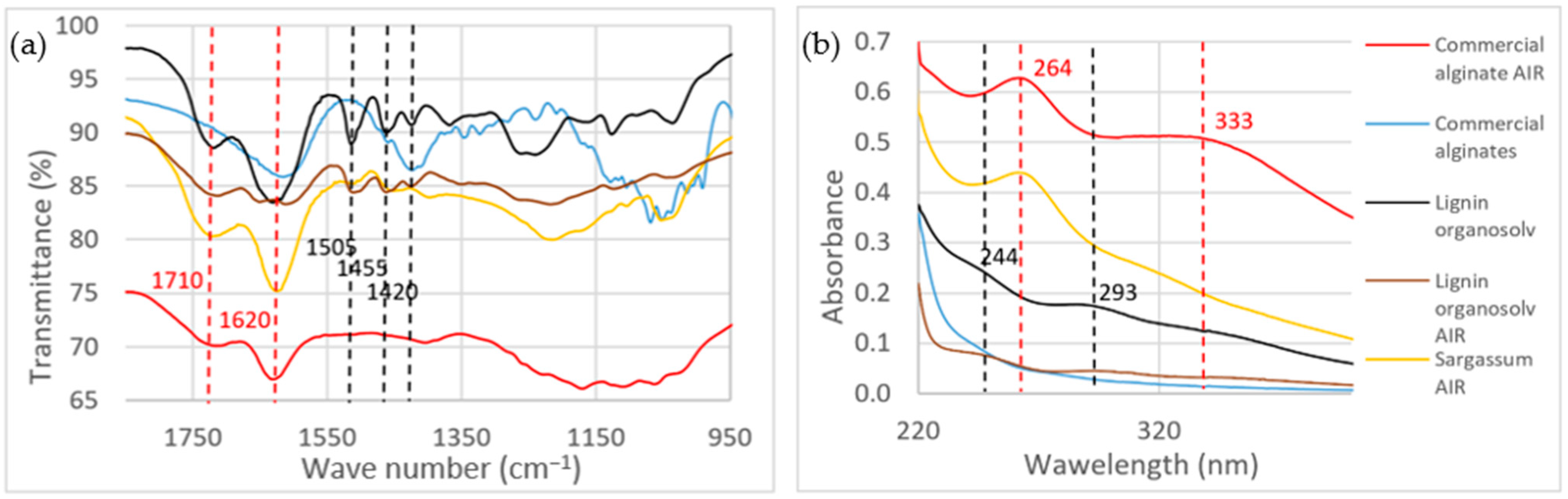
4.4. Structural Carbohydrates
4.5. Acid-Insoluble Residue
4.6. Mannitol
4.7. Polyphenols
4.8. Elemental Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kingsbury, J.M. Christopher Columbus as a Botanist. Arnoldia 1992, 52, 11–28. [Google Scholar]
- Wang, M.; Hu, C.; Barnes, B.B.; Mitchum, G.; Lapointe, B.; Montoya, J.P. The Great Atlantic Sargassum Belt. Science 2019, 365, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Gavio, B.; Rincón-Díaz, M.N.; Santos-Martínez, A. Massive quantities of pelagic Sargassum on the shores of San Andres island, southwestern Caribbean. Acta Biol. Colomb. 2014, 20, 239–241. [Google Scholar] [CrossRef]
- Oviatt, C.A.; Huizenga, K.; Rogers, C.S.; Miller, W.J. What Nutrient Sources Support Anomalous Growth and the Recent Sargassum Mass Stranding on Caribbean Beaches? A Review. Mar. Pollut. Bull. 2019, 145, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.; Harvey, P. Golden Tides: Problem or Golden Opportunity? The Valorisation of Sargassum from Beach Inundations. J. Mar. Sci. Eng. 2016, 4, 60. [Google Scholar] [CrossRef]
- Kumar, A.; Buia, M.C.; Palumbo, A.; Mohany, M.; Wadaan, M.A.M.; Hozzein, W.N.; Beemster, G.T.S.; AbdElgawad, H. Ocean Acidification Affects Biological Activities of Seaweeds: A Case Study of Sargassum vulgare from Ischia Volcanic CO2 Vents. Environ. Pollut. 2020, 259, 113765. [Google Scholar] [CrossRef] [PubMed]
- Louime, C.; Fortune, J.; Gervais, G. Sargassum Invasion of Coastal Environments: A Growing Concern. Am. J. Environ. Sci. 2017, 13, 58–64. [Google Scholar] [CrossRef]
- Johnson, D.R.; Ko, D.S.; Franks, J.S.; Moreno, P.; Sanchez-Rubio, G. The Sargassum Invasion of the Eastern Caribbean and Dynamics of the Equatorial North Atlantic, Conference Paper. 2012. Available online: https://www.researchgate.net/publication/324418761_The_Sargassum_Invasion_of_the_Eastern_Caribbean_and_Dynamics_of_the_Equatorial_North_Atlantic (accessed on 10 April 2024).
- Franks, J.S.; Johnson, D.R.; Ko, D.S.; Sanchez-Rubio, G.; Hendon, J.R.; Lay, M. Unprecedented Influx of Pelagic Sargassum along Caribbean Island Coastlines during Summer 2011; ResearchGate: Puerto Morelos, Mexico, 2011; pp. 6–8. [Google Scholar]
- García-Sánchez, M.; Graham, C.; Vera, E.; Escalante-Mancera, E.; Álvarez-Filip, L.; van Tussenbroek, B.I. Temporal Changes in the Composition and Biomass of Beached Pelagic Sargassum Species in the Mexican Caribbean. Aquat. Bot. 2020, 167, 103275. [Google Scholar] [CrossRef]
- Resiere, D.; Kallel, H.; Florentin, J.; Banydeen, R.; Compton, K.; Gueye, P.; Mehdaoui, H.; Neviere, R. Sargassum Seaweed in the Caribbean: A Major Public Health Problem Still Unsolved. J. Glob. Health 2023, 13, 03017. [Google Scholar] [CrossRef] [PubMed]
- Agence Nationale de Sécurité Sanitaire Alimentation, Environnement, Travail (ANSES). Expositions aux Émanations d’algues Sargasses en Décomposition au Antilles et en Guyane. 2017. Available online: https://www.anses.fr/fr/system/files/AIR2015SA0225Ra.pdf (accessed on 4 April 2024).
- Capron, A. Les Algues Sargasses, Cauchemar des Guadeloupéens. Available online: https://observers.france24.com/fr/20150819-algues-sargasses-cauchemar-guadeloupe-antilles (accessed on 2 July 2023).
- Resiere, D.; Mehdaoui, H.; Banydeen, R.; Florentin, J.; Kallel, H.; Nevière, R.; Mégarbane, B. Effets sanitaires de la décomposition des algues sargasses échouées sur les rivages des Antilles françaises. Toxicol. Anal. Clin. 2021, 33, 216–221. [Google Scholar] [CrossRef]
- Marsh, R.; Oxenford, H.A.; Cox, S.-A.L.; Johnson, D.R.; Bellamy, J. Forecasting Seasonal Sargassum Events across the Tropical Atlantic: Overview and Challenges. Front. Mar. Sci. 2022, 9, 914501. [Google Scholar] [CrossRef]
- Gao, H.; Duan, B.; Lu, A.; Deng, H.; Du, Y.; Shi, X.; Zhang, L. Fabrication of Cellulose Nanofibers from Waste Brown Algae and Their Potential Application as Milk Thickeners. Food Hydrocoll. 2018, 79, 473–481. [Google Scholar] [CrossRef]
- Amador-Castro, F.; García-Cayuela, T.; Alper, H.S.; Rodriguez-Martinez, V.; Carrillo-Nieves, D. Valorization of Pelagic Sargassum Biomass into Sustainable Applications: Current Trends and Challenges. J. Environ. Manag. 2021, 283, 112013. [Google Scholar] [CrossRef]
- Hsiao, W.-C.; Hong, Y.-H.; Tsai, Y.-H.; Lee, Y.-C.; Patel, A.K.; Guo, H.-R.; Kuo, C.-H.; Huang, C.-Y. Extraction, Biochemical Characterization, and Health Effects of Native and Degraded Fucoidans from Sargassum crispifolium. Polymers 2022, 14, 1812. [Google Scholar] [CrossRef] [PubMed]
- Zubia, M.; Payri, C.; Deslandes, E. Alginate, Mannitol, Phenolic Compounds and Biological Activities of Two Range-Extending Brown Algae, Sargassum mangarevense and Turbinaria ornata (Phaeophyta: Fucales), from Tahiti (French Polynesia). J. Appl. Phycol. 2008, 20, 1033–1043. [Google Scholar] [CrossRef]
- Oxenford, H.A.; Cox, S.-A.; van Tussenbroek, B.I.; Desrochers, A. Challenges of Turning the Sargassum Crisis into Gold: Current Constraints and Implications for the Caribbean. Phycology 2021, 1, 27–48. [Google Scholar] [CrossRef]
- Rossignolo, J.A.; Felicio Peres Duran, A.J.; Bueno, C.; Martinelli Filho, J.E.; Savastano Junior, H.; Tonin, F.G. Algae Application in Civil Construction: A Review with Focus on the Potential Uses of the Pelagic Sargassum spp. Biomass. J. Environ. Manag. 2022, 303, 114258. [Google Scholar] [CrossRef] [PubMed]
- Bilba, K.; Onésippe Potiron, C.; Arsène, M.-A. Invasive Biomass Algae Valorization: Assessment of the Viability of Sargassum Seaweed as Pozzolanic Material. J. Environ. Manag. 2023, 342, 118056. [Google Scholar] [CrossRef] [PubMed]
- Bauta, J.; Vaca-Medina, G.; Raynaud, C.D.; Simon, V.; Vandenbossche, V.; Rouilly, A. Development of a Binderless Particleboard from Brown Seaweed Sargassum spp. Materials 2024, 17, 539. [Google Scholar] [CrossRef] [PubMed]
- Abu-Khudir, R.; Ismail, G.A.; Diab, T. Antimicrobial, Antioxidant, and Anti-Tumor Activities of Sargassum linearifolium and Cystoseira crinita from Egyptian Mediterranean Coast. Nutr. Cancer 2021, 73, 829–844. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Mohamed, H.F.; Xu, C.; Sun, X.; Huang, L. Novel Antibacterial Activity of Sargassum fusiforme Extract against Coral White Band Disease. Electron. J. Biotechnol. 2022, 57, 12–23. [Google Scholar] [CrossRef]
- Spagnuolo, D.; Iannazzo, D.; Len, T.; Balu, A.M.; Morabito, M.; Genovese, G.; Espro, C.; Bressi, V. Hydrochar from Sargassum muticum: A Sustainable Approach for High-Capacity Removal of Rhodamine B Dye. RSC Sustain. 2023, 1, 1404–1415. [Google Scholar] [CrossRef]
- Milledge, J.J.; Maneein, S.; Arribas López, E.; Bartlett, D. Sargassum Inundations in Turks and Caicos: Methane Potential and Proximate, Ultimate, Lipid, Amino Acid, Metal and Metalloid Analyses. Energies 2020, 13, 1523. [Google Scholar] [CrossRef]
- Paredes-Camacho, R.M.; González-Morales, S.; González-Fuentes, J.A.; Rodríguez-Jasso, R.M.; Benavides-Mendoza, A.; Charles-Rodríguez, A.V.; Robledo-Olivo, A. Characterization of Sargassum spp. from the Mexican Caribbean and Its Valorization through Fermentation Process. Processes 2023, 11, 685. [Google Scholar] [CrossRef]
- Rosado-Espinosa, L.A.; Freile-Pelegrín, Y.; Hernández-Nuñez, E.; Robledo, D. A Comparative Study of Sargassum Species from the Yucatan Peninsula Coast: Morphological and Chemical Characterisation. Phycologia 2020, 59, 261–271. [Google Scholar] [CrossRef]
- Tonon, T.; Machado, C.B.; Webber, M.; Webber, D.; Smith, J.; Pilsbury, A.; Cicéron, F.; Herrera-Rodriguez, L.; Jimenez, E.M.; Suarez, J.V.; et al. Biochemical and Elemental Composition of Pelagic Sargassum Biomass Harvested across the Caribbean. Phycology 2022, 2, 204–215. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass; National Renewable Energy Laboratory: Golden, CO, USA, 2012.
- Cameron, M.C.; Ross, A.G.; Percival, E.G.V. Methods for the Routine Estimation of Mannitol, Alginic Acid, and Combined Fucose in Seaweeds. J. Chem. Technol. Biotechnol. 1948, 67, 161–164. [Google Scholar] [CrossRef]
- International Organization for Standardization Solid Biofuels—Determination of Moisture Content—Oven Dry Method—Part 2: Total Moisture—Simplified Method. Available online: https://www.iso.org/fr/standard/86024.html (accessed on 10 April 2024).
- International Organization for Standardization Solid Biofuels—Determination of Ash Content. Available online: https://www.iso.org/fr/standard/83190.html (accessed on 2 July 2023).
- International Organization for Standardization Starches and Derived Products—Determination of Nitrogen Content by the Kjeldahl Method—Titrimetric Method. Available online: https://www.iso.org/fr/standard/8379.html (accessed on 2 July 2023).
- Commission of the European Communities. COMMISSION DIRECTIVE 98/64/EC of 3 September 1998 Establishing Community Methods of Analysis for the Determination of Aminoacids, Crude Oils and Fats, and Olaquindox in Feedingstuffs and Amending Directive 71/393/EEC; Commission of the European Communities: Brussels, Belgium, 1998. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization Animal and Vegetable Fats and Oils—Gas Chromatography of Fatty Acid Methyl Esters—Part 3: Preparation of Methyl Esters Using Trimethylsulfonium Hydroxide (TMSH). Available online: https://www.iso.org/fr/standard/70249.html (accessed on 2 July 2023).
- Perez, R.; Kaas, R.; Campello, F.; Arbault, S.; Barbaroux, O. La Culture des Algues Marines dans le Monde; Institut Français de Recherche pour l’Exploitation de la Mer, Ed.; IFREMER: Plouzané, France, 1992; ISBN 978-2-905434-41-8. [Google Scholar]
- Salachna, P.; Grzeszczuk, M.; Meller, E.; Soból, M. Oligo-Alginate with Low Molecular Mass Improves Growth and Physiological Activity of Eucomis Autumnalis under Salinity Stress. Molecules 2018, 23, 812. [Google Scholar] [CrossRef] [PubMed]
- Rashid, T.; Kait, C.F.; Murugesan, T. A “Fourier Transformed Infrared” Compound Study of Lignin Recovered from a Formic Acid Process. Procedia Eng. 2016, 148, 1312–1319. [Google Scholar] [CrossRef]
- Oyesiku, O.O.; Egunyomi, A. Identification and Chemical Studies of Pelagic Masses of Sargassum natans (Linnaeus) Gaillon and S. fluitans (Borgessen) Borgesen (Brown Algae), Found Offshore in Ondo State, Nigeria. Afr. J. Biotechnol. 2014, 13, 1188–1193. [Google Scholar] [CrossRef]
- Torres-Conde, E.G.; van Tussenbroek, B.I.; Rodríguez-Martínez, R.E.; Martínez-Daranas, B. Temporal Changes in the Composition of Beached Holopelagic Sargassum spp. along the Northwestern Coast of Cuba. Phycology 2023, 3, 405–412. [Google Scholar] [CrossRef]
- Alleyne, K.S.T.; Johnson, D.; Neat, F.; Oxenford, H.A.; Vallès, H. Seasonal Variation in Morphotype Composition of Pelagic Sargassum Influx Events Is Linked to Oceanic Origin. Sci. Rep. 2023, 13, 3753. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.B.; Maddix, G.-M.; Francis, P.; Thomas, S.-L.; Burton, J.-A.; Langer, S.; Larson, T.R.; Marsh, R.; Webber, M.; Tonon, T. Pelagic Sargassum Events in Jamaica: Provenance, Morphotype Abundance, and Influence of Sample Processing on Biochemical Composition of the Biomass. Sci. Total Environ. 2022, 817, 152761. [Google Scholar] [CrossRef]
- Schell, J.M.; Goodwin, D.S.; Volk, R.H.; Siuda, A.N.S. Preliminary Explorations of Environmental Tolerances and Growth Rates of Holopelagic Sargassum Morphotypes. Aquat. Bot. 2024, 190, 103723. [Google Scholar] [CrossRef]
- Davis, D.; Simister, R.; Campbell, S.; Marston, M.; Bose, S.; McQueen-Mason, S.J.; Gomez, L.D.; Gallimore, W.A.; Tonon, T. Biomass Composition of the Golden Tide Pelagic Seaweeds Sargassum fluitans and S. natans (Morphotypes I and VIII) to Inform Valorisation Pathways. Sci. Total Environ. 2021, 762, 143134. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Flores, P.A.; Serviere-Zaragoza, E.; De Anda-Montañez, J.A.; Freile-Pelegrín, Y.; Robledo, D.; Méndez-Rodríguez, L.C. Trace Elements in Pelagic Sargassum Species in the Mexican Caribbean: Identification of Key Variables Affecting Arsenic Accumulation in S. fluitans. Sci. Total Environ. 2022, 806, 150657. [Google Scholar] [CrossRef] [PubMed]
- Olguin-Maciel, E.; Leal-Bautista, R.M.; Alzate-Gaviria, L.; Domínguez-Maldonado, J.; Tapia-Tussell, R. Environmental Impact of Sargassum spp. Landings: An Evaluation of Leachate Released from Natural Decomposition at Mexican Caribbean Coast. Environ. Sci. Pollut. Res. 2022, 29, 91071–91080. [Google Scholar] [CrossRef] [PubMed]
- Brust, G.E. Management Strategies for Organic Vegetable Fertility. In Safety and Practice for Organic Food; Elsevier: Amsterdam, The Netherlands, 2019; pp. 193–212. ISBN 978-0-12-812060-6. [Google Scholar]
- Mæhre, H.; Dalheim, L.; Edvinsen, G.; Elvevoll, E.; Jensen, I.-J. Protein Determination—Method Matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Sosulski, F.W.; Imafidon, G.I. Amino Acid Composition and Nitrogen-to-Protein Conversion Factors for Animal and Plant Foods. J. Agric. Food Chem. 1990, 38, 1351–1356. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The Protein Content of Seaweeds: A Universal Nitrogen-to-Protein Conversion Factor of Five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Mohammed, A.; Rivers, A.; Stuckey, D.C.; Ward, K. Alginate Extraction from Sargassum Seaweed in the Caribbean Region: Optimization Using Response Surface Methodology. Carbohydr. Polym. 2020, 245, 116419. [Google Scholar] [CrossRef] [PubMed]
- Gordillo Sierra, A.R.; Amador-Castro, L.F.; Ramírez-Partida, A.E.; García-Cayuela, T.; Carrillo-Nieves, D.; Alper, H.S. Valorization of Caribbean Sargassum Biomass as a Source of Alginate and Sugars for de Novo Biodiesel Production. J. Environ. Manag. 2022, 324, 116364. [Google Scholar] [CrossRef] [PubMed]
- Karim, H.; Ahmad, A.; Natzir, R.; Massi, M.N.; Arfah, R.; Asmi, N.; Karim, A. Isolation and Identification of Bioactive Proteins from the Brown Algae Sargassum, Sp. and Their Potential as Anticancer Agents. J. Phys. Conf. Ser. 2019, 1341, 032009. [Google Scholar] [CrossRef]
- O’ Connor, J.; Meaney, S.; Williams, G.A.; Hayes, M. Extraction of Protein from Four Different Seaweeds Using Three Different Physical Pre-Treatment Strategies. Molecules 2020, 25, 2005. [Google Scholar] [CrossRef] [PubMed]
- van Ginneken, V.J.; Helsper, J.P.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated Fatty Acids in Various Macroalgal Species from North Atlantic and Tropical Seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef]
- Christie, W.W.; Han, X. Lipid Analysis: Isolation, Separation, Identification and Lipidomic Analysis, 4th ed.; The Oily Press Lipid Library; The Oily Press, PJ Barnes & Associates: Bridgwater, UK, 2010; ISBN 978-0-9552512-4-5. [Google Scholar]
- Harrysson, H.; Krook, J.L.; Larsson, K.; Tullberg, C.; Oerbekke, A.; Toth, G.; Pavia, H.; Undeland, I. Effect of Storage Conditions on Lipid Oxidation, Nutrient Loss and Colour of Dried Seaweeds, Porphyra Umbilicalis and Ulva Fenestrata, Subjected to Different Pretreatments. Algal Res. 2021, 56, 102295. [Google Scholar] [CrossRef]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Evaluation of Food Grade Solvents for Lipid Extraction and Impact of Storage Temperature on Fatty Acid Composition of Edible Seaweeds Laminaria digitata (Phaeophyceae) and Palmaria palmata (Rhodophyta). Food Chem. 2016, 208, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Perez, P.; Lourenço-Lopes, C.; Silva, A.; Pereira, A.G.; Fraga-Corral, M.; Zhao, C.; Xiao, J.; Simal-Gandara, J.; Prieto, M.A. Pigment Composition of Nine Brown Algae from the Iberian Northwestern Coastline: Influence of the Extraction Solvent. Mar. Drugs 2022, 20, 113. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Hu, C.; Cannizzaro, J.; English, D.; Han, X.; Naar, D.; Lapointe, B.; Brewton, R.; Hernandez, F. Remote Sensing of Sargassum Biomass, Nutrients, and Pigments. Geophys. Res. Lett. 2018, 45, 12359–12367. [Google Scholar] [CrossRef]
- Davis, T.A.; Volesky, B.; Mucci, A. A Review of the Biochemistry of Heavy Metal Biosorption by Brown Algae. Water Res. 2003, 37, 4311–4330. [Google Scholar] [CrossRef] [PubMed]
- del Río, P.G.; Domínguez, E.; Domínguez, V.D.; Romaní, A.; Domingues, L.; Garrote, G. Third Generation Bioethanol from Invasive Macroalgae Sargassum muticum Using Autohydrolysis Pretreatment as First Step of a Biorefinery. Renew. Energy 2019, 141, 728–735. [Google Scholar] [CrossRef]
- Aparicio, E.; Rodríguez-Jasso, R.M.; Pinales-Márquez, C.D.; Loredo-Treviño, A.; Robledo-Olivo, A.; Aguilar, C.N.; Kostas, E.T.; Ruiz, H.A. High-Pressure Technology for Sargassum Spp Biomass Pretreatment and Fractionation in the Third Generation of Bioethanol Production. Bioresour. Technol. 2021, 329, 124935. [Google Scholar] [CrossRef] [PubMed]
- Bonilla Loaiza, A.M.; Rodríguez-Jasso, R.M.; Belmares, R.; López-Badillo, C.M.; Araújo, R.G.; Aguilar, C.N.; Chávez, M.L.; Aguilar, M.A.; Ruiz, H.A. Fungal Proteins from Sargassum spp. Using Solid-State Fermentation as a Green Bioprocess Strategy. Molecules 2022, 27, 3887. [Google Scholar] [CrossRef] [PubMed]
- Rhein-Knudsen, N.; Ale, M.T.; Ajalloueian, F.; Meyer, A.S. Characterization of Alginates from Ghanaian Brown Seaweeds: Sargassum spp. and Padina spp. Food Hydrocoll. 2017, 71, 236–244. [Google Scholar] [CrossRef]
- Hasnain, M.S.; Nayak, A.K. (Eds.) Alginates: Versatile Polymers in Biomedical Applications and Therapeutics, 1st ed.; Apple Academic Press: Palm Bay, FL, USA, 2019; ISBN 978-0-429-02343-9. [Google Scholar]
- Bi, D.; Yang, X.; Yao, L.; Hu, Z.; Li, H.; Xu, X.; Lu, J. Potential Food and Nutraceutical Applications of Alginate: A Review. Mar. Drugs 2022, 20, 564. [Google Scholar] [CrossRef] [PubMed]
- de Souza Coração, A.C.; dos Santos, F.S.; Duarte, J.A.D.; Lopes-Filho, E.A.P.; De-Paula, J.C.; Rocha, L.M.; Krepsky, N.; Fiaux, S.B.; Teixeira, V.L. What Do We Know about the Utilization of the Sargassum Species as Biosorbents of Trace Metals in Brazil? J. Environ. Chem. Eng. 2020, 8, 103941. [Google Scholar] [CrossRef]
- Chale-Dzul, J.; Pérez-Cabeza de Vaca, R.; Quintal-Novelo, C.; Olivera-Castillo, L.; Moo-Puc, R. Hepatoprotective Effect of a Fucoidan Extract from Sargassum fluitans Borgesen against CCl4-Induced Toxicity in Rats. Int. J. Biol. Macromol. 2020, 145, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Lee, H.G.; Kim, H.-S.; Vaas, A.P.J.P.; De Silva, H.I.C.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, W.W.; Lee, D.-S.; et al. Characterization and Cytoprotective Properties of Sargassum natans Fucoidan against Urban Aerosol-Induced Keratinocyte Damage. Int. J. Biol. Macromol. 2020, 159, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, Structure and Biofunctional Activities of Laminarin from Brown Algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, X.; Tang, Y.; Mao, J. Composition, Isolation, Purification and Biological Activities of Sargassum fusiforme Polysaccharides: A Review. Carbohydr. Polym. 2020, 228, 115381. [Google Scholar] [CrossRef] [PubMed]
- Alzate-Gaviria, L.; Domínguez-Maldonado, J.; Chablé-Villacís, R.; Olguin-Maciel, E.; Leal-Bautista, R.M.; Canché-Escamilla, G.; Caballero-Vázquez, A.; Hernández-Zepeda, C.; Barredo-Pool, F.A.; Tapia-Tussell, R. Presence of Polyphenols Complex Aromatic “Lignin” in Sargassum spp. from Mexican Caribbean. J. Mar. Sci. Eng. 2020, 9, 6. [Google Scholar] [CrossRef]
- Chen, H. Lignocellulose Biorefinery Feedstock Engineering. In Lignocellulose Biorefinery Engineering–Principles and Applications, 1st ed.; Woodhead Publishing: Sawston, UK, 2015; pp. 37–86. ISBN 978-0-08-100145-5. [Google Scholar]
- Weng, J.; Chapple, C. The Origin and Evolution of Lignin Biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Alzagameem, A.; Khaldi-Hansen, B.; Büchner, D.; Larkins, M.; Kamm, B.; Witzleben, S.; Schulze, M. Lignocellulosic Biomass as Source for Lignin-Based Environmentally Benign Antioxidants. Molecules 2018, 23, 2664. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, N.; Mitomo, H.; Yoshii, F.; Kume, T. Radiation-Induced Degradation of Sodium Alginate. Polym. Degrad. Stab. 2000, 69, 279–285. [Google Scholar] [CrossRef]
- Wasikiewicz, J.M.; Yoshii, F.; Nagasawa, N.; Wach, R.A.; Mitomo, H. Degradation of Chitosan and Sodium Alginate by Gamma Radiation, Sonochemical and Ultraviolet Methods. Radiat. Phys. Chem. 2005, 73, 287–295. [Google Scholar] [CrossRef]
- Borines, M.G.; de Leon, R.L.; Cuello, J.L. Bioethanol Production from the Macroalgae Sargassum spp. Bioresour. Technol. 2013, 138, 22–29. [Google Scholar] [CrossRef]
- Groisillier, A.; Shao, Z.; Michel, G.; Goulitquer, S.; Bonin, P.; Krahulec, S.; Nidetzky, B.; Duan, D.; Boyen, C.; Tonon, T. Mannitol Metabolism in Brown Algae Involves a New Phosphatase Family. EXBOTJ 2014, 65, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Komoe, K.; Sankare, Y.; Fofie, N.B.Y.; Bamba, A.; SAHR, A.G.-S. Taxonomic Study of Two Species of Sargassum: Sargassum fluitans (Børgesen) Børgesen and Sargassum natans (Linnaneus) Gaillon (Brown Algae) Collected in Côte d’Ivoire Coasts, West Africa. Nat. Sci. 2016, 14, 50–56. [Google Scholar] [CrossRef]
- Saha, B.C.; Racine, F.M. Biotechnological Production of Mannitol and Its Applications. Appl. Microbiol. Biotechnol. 2011, 89, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Imbs, T.I.; Zvyagintseva, T.N. Phlorotannins Are Polyphenolic Metabolites of Brown Algae. Russ. J. Mar. Biol. 2018, 44, 263–273. [Google Scholar] [CrossRef]
- Saldarriaga-Hernandez, S.; Melchor-Martínez, E.M.; Carrillo-Nieves, D.; Parra-Saldívar, R.; Iqbal, H.M.N. Seasonal Characterization and Quantification of Biomolecules from Sargassum Collected from Mexican Caribbean Coast—A Preliminary Study as a Step Forward to Blue Economy. J. Environ. Manag. 2021, 298, 113507. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, V.; Duan, X.; Agar, O.T.; Dunshea, F.R.; Barrow, C.J.; Suleria, H.A.R. Comparative Study on the Effect of Different Drying Techniques on Phenolic Compounds in Australian Beach-Cast Brown Seaweeds. Algal Res. 2023, 72, 103140. [Google Scholar] [CrossRef]
- Zhao, T.; Dong, Q.; Zhou, H.; Yang, H. Drying Kinetics, Physicochemical Properties, Antioxidant Activity and Antidiabetic Potential of Sargassum fusiforme Processed under Four Drying Techniques. LWT 2022, 163, 113578. [Google Scholar] [CrossRef]
- Le Lann, K.; Jégou, C.; Stiger-Pouvreau, V. Effect of Different Conditioning Treatments on Total Phenolic Content and Antioxidant Activities in Two Sargassacean Species: Comparison of the Frondose Sargassum muticum (Yendo) Fensholt and the Cylindrical Bifurcaria bifurcata R. Ross. Phycol. Res. 2008, 56, 238–245. [Google Scholar] [CrossRef]
- Charles, A.L.; Sridhar, K.; Alamsjah, M.A. Effect of Drying Techniques on Color and Bioactive Potential of Two Commercial Edible Indonesian Seaweed Cultivars. J. Appl. Phycol. 2020, 32, 563–572. [Google Scholar] [CrossRef]
- Uribe, E.; Pardo-Orellana, C.M.; Vega-Gálvez, A.; Ah-Hen, K.S.; Pastén, A.; García, V.; Aubourg, S.P. Effect of Drying Methods on Bioactive Compounds, Nutritional, Antioxidant, and Antidiabetic Potential of Brown Alga Durvillaea antarctica. Dry. Technol. 2020, 38, 1915–1928. [Google Scholar] [CrossRef]
- Amsler, C.D.; Fairhead, V.A. Defensive and Sensory Chemical Ecology of Brown Algae. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2005; Volume 43, pp. 1–91. ISBN 978-0-12-005943-0. [Google Scholar]
- Stiger, V.; Deslandes, E.; Payri, C.E. Phenolic Contents of Two Brown Algae, Turbinaria Ornata and Sargassum mangarevense on Tahiti (French Polynesia): Interspecific, Ontogenic and Spatio-Temporal Variations. Bot. Mar. 2004, 47, 402–409. [Google Scholar] [CrossRef]
- Fernandez, A.; Singh, A.; Jaffé, R. A Literature Review on Trace Metals and Organic Compounds of Anthropogenic Origin in the Wider Caribbean Region. Mar. Pollut. Bull. 2007, 54, 1681–1691. [Google Scholar] [CrossRef]
- Idota, Y.; Kogure, Y.; Kato, T.; Yano, K.; Arakawa, H.; Miyajima, C.; Kasahara, F.; Ogihara, T. Relationship between Physical Parameters of Various Metal Ions and Binding Affinity for Alginate. Biol. Pharm. Bull. 2016, 39, 1893–1896. [Google Scholar] [CrossRef]
- Mohammed, C.; Lalgee, L.; Kistow, M.; Jalsa, N.; Ward, K. On the Binding Affinity and Thermodynamics of Sodium Alginate-Heavy Metal Ion Interactions for Efficient Adsorption. Carbohydr. Polym. Technol. Appl. 2022, 3, 100203. [Google Scholar] [CrossRef]
- Lim, Y.-C.; Chen, C.-F.; Tsai, M.-L.; Wu, C.-H.; Lin, Y.-L.; Wang, M.-H.; Albarico, F.P.J.B.; Chen, C.-W.; Dong, C.-D. Impacts of Fishing Vessels on the Heavy Metal Contamination in Sediments: A Case Study of Qianzhen Fishing Port in Southern Taiwan. Water 2022, 14, 1174. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, R.E.; Roy, P.D.; Torrescano-Valle, N.; Cabanillas-Terán, N.; Carrillo-Domínguez, S.; Collado-Vides, L.; García-Sánchez, M.; van Tussenbroek, B.I. Element Concentrations in Pelagic Sargassum along the Mexican Caribbean Coast in 2018–2019. PeerJ 2020, 8, e8667. [Google Scholar] [CrossRef] [PubMed]
- European Commission. COMMISSION REGULATION (EU) 2019/1869 of 7 November 2019 Amending and Correcting Annex I to Directive 2002/32/EC of the European Parliament and of the Council as Regards Maximum Levels for Certain Undesirable Substances in Animal Feed; European Commission: Brussels, Belgium, 2019. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Location of Harvest | Harvest Date |
|---|---|---|
| DPbwn21 | Dominican Republic, Punta Cana | October 2021 |
| GLbwn21 | Guadeloupe, Le Gosier | September 2021 |
| GVbwn21 | Guadeloupe, Vieux Habitants | June 2021 |
| GBbwn22 | Guadeloupe, Bouillante en mer | January 2022 |
| GFpwn22 | Guadeloupe, Saint Felix seaport | July 2022 |
| MGbwn22 | Marie Galante, Plage des Galets | June 2022 |
| MGbfn22 | Marie Galante, Plage des Galets | June 2022 |
| MCbwn22 | Marie Galante, Capesterre | June 2022 |
| MCbws22 | Marie Galante, Capesterre | June 2022 |
| DPbwn21 | GLbwn21 | GVbwn21 | GBbwn22 | GFpwn22 | MGbwn22 | MGbfn22 | MCbwn22 | MCbws22 | |
|---|---|---|---|---|---|---|---|---|---|
| Proximate analysis | |||||||||
| Dry matter (%) | 83.30 ± 0.15 | 82.21 ± 0.10 | 84.80 ± 0.06 | 82.59 ± 0.19 | 86.23 ± 0.29 | 87.45 ± 0.20 | 85.80 ± 0.11 | 86.09 ± 0.40 | 85.69 ± 0.12 |
| Mineral matter | 15.49 ± 0.41 | 16.18 ± 0.57 | 14.81 ± 0.25 | 19.86 ± 0.53 | 14.48 ± 0.64 | 14.79 ± 0.40 | 17.06 ± 0.25 | 16.99 ± 0.39 | 18.55 ± 0.73 |
| C | 38.75 ± 0.02 | 37.28 ± 0.13 | 37.44 ± 0.16 | 36.82 ± 0.33 | 37.10 ± 0.13 | 38.89 ± 0.03 | 35.53 ± 0.07 | 37.22 ± 0.01 | 37.86 ± 0.08 |
| H | 6.28 ± 0.17 | 5.39 ± 0.03 | 5.47 ± 0.04 | 5.88 ± 0.09 | 6.59 ± 0.16 | 5.94 ± 0.02 | 5.41 ± 0.09 | 5.41 ± 0.02 | 5.88 ± 0.05 |
| N | 1.10 ± 0.10 | 0.98 ± 0.01 | 1.13 ± 0.11 | 1.25 ± 0.05 | 1.01 ± 0.02 | 1.00 ± 0.06 | 0.86 ± 0.08 | 1.13 ± 0.05 | 1.39 ± 0.02 |
| Proteins | |||||||||
| Kjeldahl organic Nitrogen | 0.95 ± 0.01 | 1.01 ± 0.01 | 0.98 ± 0.01 | 1.18 ± 0.01 | 0.97 ± 0.01 | 0.86 ± 0.01 | 0.74 ± 0.01 | 1.05 ± 0.01 | 1.17 ± 0.01 |
| Kjeldahl Proteins | 6.19 ± 0.06 e | 6.59 ± 0.05 c | 6.38 ± 0.02 d | 7.69 ± 0.08 a | 6.30 ± 0.02 de | 5.58 ± 0.08 f | 4.80 ± 0.02 g | 6.88 ± 0.02 b | 7.64 ± 0.06 a |
| Total amino acids | 5.27 ± 0.05 cd | 5.40 ± 0.07 c | 5.20 ± 0.14 cd | 6.11 ± 0.05 ab | 5.05 ± 0.22 d | 4.37 ± 0.07 e | 3.86 ± 0.04 f | 5.88 ± 0.10 b | 6.32 ± 0.14 a |
| Nitrogen in AA (%) | 15.40 ± 0.06 | 15.47 ± 0.02 | 15.46 ± 0.02 | 15.47 ± 0.04 | 15.21 ± 0.42 | 14.58 ± 0.07 | 14.04 ± 0.02 | 15.39 ± 0.04 | 15.41 ± 0.03 |
| N/Protein conversion factor (Nprot) | 6.49 ± 0.03 | 6.46 ± 0.01 | 6.47 ± 0.01 | 6.46 ± 0.02 | 6.58 ± 0.18 | 6.86 ± 0.03 | 7.12 ± 0.01 | 6.50 ± 6.50 | 6.49 ± 0.01 |
| Lipids | |||||||||
| Total lipid content | 0.50 ± 0.05 c | 0.81 ± 0.01 b | 0.58 ± 0.01 c | 0.28 ± 0.16 d | 1.05 ± 0.07 a | 1.10 ± 0.02 a | 0.40 ± 0.01 cd | 0.63 ± 0.08 bc | 1.03 ± 0.06 a |
| Total FAME | 0.14 ± 0.01 | 0.22 ± 0.01 | 0.14 ± 0.01 | 0.03 ± 0.01 | 0.32 ± 0.01 | 0.34 ± 0.01 | 0.10 ± 0.01 | 0.16 ± 0.01 | 0.28 ± 0.01 |
| Carbohydrates | |||||||||
| Glucan | 17.26 ± 0.64 bcd | 14.59 ± 2.37 cd | 15.94 ± 3.31 d | 16.94 ± 0.66 bcd | 19.74 ± 0.69 abc | 22.00 ± 0.40 a | 20.68 ± 1.17 ab | 20.44 ± 0.92 abc | 20.83 ± 1.86 ab |
| Fucan | 8.99 ± 0.21 de | 10.25 ± 0.11 bc | 11.28 ± 0.29 b | 12.63 ± 0.26 a | 8.21 ± 0.01 e | 8.42 ± 0.60 e | 9.55 ± 0.12 cd | 9.22 ± 0.56 cde | 9.23 ± 0.64 cde |
| AIR | 23.8 ± 1.4 ab | 24.8 ± 0.5 ab | 25.1 ± 0.1 ab | 25.8 ± 1.4 ab | 23.4 ± 0.5 b | 26.4 ± 0.3 a | 23.7 ± 1.0 b | 25.9 ± 0.7 ab | 23.2 ± 1.5 b |
| Alginate content | 25.83 ± 6.89 ab | 23.05 ± 1.14 ab | 25.45 ± 1.28 ab | 30.19 ± 1.80 a | 17.75 ± 0.44 b | 17.88 ± 1.57 b | 19.00 ± 4.32 b | 19.21 ± 1.40 b | 17.82 ± 3.24 b |
| Mannitol | 2.47 ± 0.09 ab | 2.24 ± 0.07 abc | 2.00 ± 0.27 bcd | 1.95 ± 0.08 cd | 2.80 ± 0.20 a | 2.53 ± 0.43 ab | 1.60 ± 0.08 d | 2.18 ± 0.15 bc | 2.14 ± 0.09 bc |
| TPC (mg/g GAE) | 10.79 ± 0.33 ab | 10.75 ± 0.31 ab | 8.12 ± 0.47 cd | 8.08 ± 0.22cd | 9.95 ± 0.20 b | 11.02 ± 0.29 a | 8.70 ± 0.29 c | 7.45 ± 0.22 d | 8.20 ± 0.36 cd |
| DPbwn21 | GLbwn21 | GVbwn21 | GBbwn22 | GFpwn22 | MGbwn22 | MGbfn22 | MCbwn22 | MCbws22 | |
|---|---|---|---|---|---|---|---|---|---|
| C12:0 | 0.44 ± 0.07 | 0.76 ± 0.44 | 0.58 ± 0.11 | 1.04 ± 0.43 | 0.83 ± 0.85 | 0.45 ± 0.17 | 1.68 ± 0.64 | 0.58 ± 0.47 | 0.67 ± 0.34 |
| C13:0 | 0.20 ± 0.07 | n.d. | 0.04 ± 0.06 | n.d. | 0.13 ± 0.01 | 0.21 ± 0.03 | 0.12 ± 0.10 | 0.11 ± 0.09 | 0.04 ± 0.07 |
| C14:0 | 1.82 ± 0.20 | 3.85 ± 0.21 | 3.36 ± 0.06 | 2.15 ± 0.87 | 3.78 ± 0.20 | 3.57 ± 0.13 | 2.98 ± 0.20 | 2.97 ± 0.12 | 3.22 ± 0.42 |
| C14:1n-5 | 0.31 ± 0.08 | 0.15 ± 0.13 | 0.06 ± 0.10 | 0.27 ± 0.46 | 0.09 ± 0.08 | 0.08 ± 0.07 | 0.42 ± 0.03 | 0.32 ± 0.01 | 0.14 ± 0.12 |
| C16:0 | 35.11 ± 4.04 | 55.57 ± 2.18 | 53.22 ± 1.14 | 37.97 ± 4.92 | 46.09 ± 2.47 | 41.09 ± 0.73 | 41.33 ± 2.08 | 57.33 ± 2.49 | 48.75 ± 0.70 |
| C16:1n-7 | 2.74 ± 0.37 | 7.39 ± 0.12 | 5.65 ± 0.16 | 2.09 ± 0.46 | 5.47 ± 0.26 | 6.38 ± 0.15 | 4.79 ± 0.11 | 3.11 ± 0.21 | 4.49 ± 0.08 |
| C17:0 | 0.44 ± 0.16 | n.d. | 0.40 ± 0.34 | 0.50 ± 0.86 | 0.57 ± 0.50 | 0.54 ± 0.07 | 0.69 ± 0.60 | 0.59 ± 0.12 | 0.51 ± 0.06 |
| C17:1 | 0.06 ± 0.05 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.04 ± 0.06 | 0.07 ± 0.07 |
| C18:0 | 6.79 ± 0.93 | 2.57 ± 0.27 | 3.74 ± 0.07 | 11.74 ± 3.93 | 3.86 ± 0.40 | 3.02 ± 0.23 | 8.69 ± 0.96 | 5.39 ± 0.29 | 3.99 ± 0.05 |
| C18:1n-9t | 0.56 ± 0.04 | n.d. | 0.10 ± 0.18 | n.d. | 0.40 ± 0.05 | 0.29 ± 0.06 | n.d. | 0.09 ± 0.16 | n.d. |
| C18:1n-9 | 24.57 ± 1.38 | 16.56 ± 0.06 | 18.84 ± 2.22 | 14.61 ± 2.10 | 17.15 ± 0.76 | 17.32 ± 1.66 | 19.80 ± 3.77 | 12.30 ± 0.46 | 16.63 ± 0.60 |
| C18:2n-6t | 0.34 ± 0.06 | 1.13 ± 0.14 | 0.61 ± 0.54 | n.d. | 0.71 ± 0.65 | 1.00 ± 0.88 | 1.31 ± 0.11 | 0.55 ± 0.15 | 0.84 ± 0.12 |
| C18:2n-6 | 15.89 ± 1.12 | 3.38 ± 3.42 | 5.40 ± 0.11 | 12.15 ± 6.15 | 6.38 ± 0.36 | 7.87 ± 0.35 | 10.68 ± 0.05 | 5.39 ± 0.66 | 6.06 ± 0.10 |
| C18:3n-6 | n.d. | n.d. | n.d. | n.d. | 0.16 ± 0.01 | 0.26 ± 0.01 | n.d. | n.d. | 0.11 ± 0.19 |
| C20:0 | 0.42 ± 0.38 | 0.39 ± 0.33 | 0.20 ± 0.35 | n.d. | 0.48 ± 0.08 | 0.34 ± 0.02 | 0.36 ± 0.31 | 0.39 ± 0.34 | 0.28 ± 0.24 |
| C18:3n-3a | 6.11 ± 4.38 | n.d. | 1.41 ± 0.32 | n.d. | 3.37 ± 0.64 | 3.91 ± 0.14 | 0.96 ± 0.02 | 3.27 ± 2.69 | 2.74 ± 0.31 |
| C22:0 | 2.36 ± 0.26 | 7.98 ± 0.03 | 5.90 ± 0.13 | n.d. | 9.66 ± 0.30 | 12.76 ± 0.24 | 4.03 ± 0.08 | 6.36 ± 0.30 | 10.54 ± 0.19 |
| Saturated | 49.42 ± 6.5 | 71.39 ± 3.57 | 67.92 ± 1.56 | 71.78 ± 4.00 | 66.27 ± 2.13 | 62.74 ± 1.51 | 62.05 ± 3.71 | 74.93 ± 3.45 | 68.93 ± 0.60 |
| Mono- unsaturated | 28.24 ± 0.94 | 24.11 ± 0.03 | 24.65 ± 2.04 | 16.96 ± 1.76 | 23.12 ± 0.56 | 24.21 ± 1.40 | 25.01 ± 3.64 | 15.86 ± 0.35 | 21.33 ± 0.48 |
| Poly- Unsaturated | 22.33 ± 5.56 | 4.51 ± 3.56 | 7.42 ± 0.82 | 12.15 ± 6.15 | 10.61 ± 1.58 | 13.04 ± 0.64 | 12.95 ± 0.09 | 9.21 ± 3.28 | 9.75 ± 0.42 |
| FAME/ Total lipid (%) | 17.04 ± 1.62 | 37.24 ± 1.90 | 27.40 ± 2.77 | 11.79 ± 6.97 | 30.19 ± 2.19 | 31.09 ± 0.99 | 24.00 ± 1.17 | 25.24 ± 3.34 | 26.8 ± 1.74 |
| Total %DW | 0.138 ± 0.013 | 0.216 ± 0.011 | 0.137 ± 0.002 | 0.033 ± 0.005 | 0.317 ± 0.090 | 0.342 ± 0.009 | 0.096 ± 0.004 | 0.159 ± 0.006 | 0.276 ± 0.008 |
| DPbwn21 | GLbwn21 | GVbwn21 | GBbwn22 | GFpwn22 | MGbwn22 | MGbfn22 | MCbwn22 | MCbws22 | |
|---|---|---|---|---|---|---|---|---|---|
| Na | 13,182 ± 509 | 7819 ± 83 | 5528 ± 116 | 6835 ± 116 | 9066 ± 322 | 6833 ± 182 | 17,001 ± 273 | 8227 ± 215 | 8808 ± 191 |
| Mg | 8229 ± 260 | 10,010 ± 128 | 11,244 ± 258 | 11,802 ± 198 | 8991 ± 256 | 9799 ± 50 | 15,111 ± 118 | 11,181 ± 234 | 11,400 ± 260 |
| S | 14,609 ± 766 | 14,692 ± 429 | 14,806 ± 388 | 15,260 ± 579 | 14,317 ± 708 | 14,888 ± 469 | 16,380 ± 393 | 16,213 ± 288 | 17,299 ± 595 |
| Ca | 44,315 ± 805 | 55,616 ± 868 | 62,398 ± 298 | 63,698 ± 180 | 43,208 ± 1253 | 48,415 ± 67 | 53,980 ± 377 | 59,024 ± 1044 | 60,833 ± 1378 |
| Cr | 0.595 ± 0.007 | 0.592 ± 0.015 | 0.857 ± 0.001 | 0.903 ± 0.015 | 2.222 ± 0.025 | 0.571 ± 0.011 | 0.916 ± 0.020 | 0.506 ± 0.002 | 0.737 ± 0.012 |
| Co | 1.096 ± 0.010 | 0.457 ± 0.006 | 0.824 ± 0.003 | 1.224 ± 0.014 | 0.686 ± 0.015 | 0.662 ± 0.006 | 0.517 ± 0.007 | 0.593 ± 0.004 | 0.611 ± 0.014 |
| Ni | 5.806 ± 0.087 | 5.277 ± 0.037 | 4.497 ± 0.041 | 5.061 ± 0.057 | 6.582 ± 0.139 | 5.391 ± 0.073 | 5.496 ± 0.045 | 4.104 ± 0.033 | 4.223 ± 0.047 |
| Cu | 4.361 ± 0.087 | 4.134 ± 0.018 | 10.650 ± 0.171 | 9.762 ± 0.060 | 4.236 ± 0.069 | 3.516 ± 0.006 | 3.322 ± 0.044 | 2.362 ± 0.018 | 2.495 ± 0.022 |
| Zn | 6.726 ± 0.040 | 3.726 ± 0.031 | 59.419 ± 0.917 | 47.667 ± 0.202 | 5.337 ± 0.025 | 4.786 ± 0.098 | 14.180 ± 0.060 | 2.860 ± 0.036 | 3.401 ± 0.078 |
| As | 48.280 ± 0.065 | 53.223 ± 0.434 | 16.764 ± 0.476 | 17.597 ± 0.184 | 85.979 ± 1.026 | 89.379 ± 0.674 | 19.062 ± 0.254 | 17.968 ± 0.188 | 17.719 ± 0.374 |
| Cd | 0.660 ± 0.004 | 0.587 ± 0.028 | 0.694 ± 0.003 | 0.849 ± 0.018 | 0.765 ± 0.004 | 0.739 ± 0.003 | 0.471 ± 0.006 | 0.598 ± 0.006 | 1.182 ± 0.021 |
| Pb | 0.232 ± 0.002 | 0.712 ± 0.005 | 0.388 ± 0.005 | 0.488 ± 0.011 | 0.440 ± 0.004 | 0.348 ± 0.006 | 1.551 ± 0.004 | 0.534 ± 0.008 | 0.394 ± 0.005 |
| Hg | 8.790 ± 1.034 | 7.687 ± 0.904 | 10.568 ± 1.243 | 11.157 ± 1.313 | 6.278 ± 0.739 | 4.514 ± 0.531 | 4.86 ± 0.572 | 4.434 ± 0.522 | 6.872 ± 0.809 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauta, J.; Calbrix, E.; Capblancq, S.; Cecutti, C.; Peydecastaing, J.; Delgado Raynaud, C.; Rouilly, A.; Simon, V.; Vaca-Medina, G.; Vandenbossche, V.; et al. Global Chemical Characterization of Sargassum spp. Seaweeds from Different Locations on Caribbean Islands: A Screening of Organic Compounds and Heavy Metals Contents. Phycology 2024, 4, 190-212. https://doi.org/10.3390/phycology4020011
Bauta J, Calbrix E, Capblancq S, Cecutti C, Peydecastaing J, Delgado Raynaud C, Rouilly A, Simon V, Vaca-Medina G, Vandenbossche V, et al. Global Chemical Characterization of Sargassum spp. Seaweeds from Different Locations on Caribbean Islands: A Screening of Organic Compounds and Heavy Metals Contents. Phycology. 2024; 4(2):190-212. https://doi.org/10.3390/phycology4020011
Chicago/Turabian StyleBauta, Jérôme, Elliot Calbrix, Sophie Capblancq, Christine Cecutti, Jérôme Peydecastaing, Christine Delgado Raynaud, Antoine Rouilly, Valérie Simon, Guadalupe Vaca-Medina, Virginie Vandenbossche, and et al. 2024. "Global Chemical Characterization of Sargassum spp. Seaweeds from Different Locations on Caribbean Islands: A Screening of Organic Compounds and Heavy Metals Contents" Phycology 4, no. 2: 190-212. https://doi.org/10.3390/phycology4020011
APA StyleBauta, J., Calbrix, E., Capblancq, S., Cecutti, C., Peydecastaing, J., Delgado Raynaud, C., Rouilly, A., Simon, V., Vaca-Medina, G., Vandenbossche, V., Vedrenne, E., & De Caro, P. (2024). Global Chemical Characterization of Sargassum spp. Seaweeds from Different Locations on Caribbean Islands: A Screening of Organic Compounds and Heavy Metals Contents. Phycology, 4(2), 190-212. https://doi.org/10.3390/phycology4020011