
Quantification and Immunolocalization of Auxin in Prunus dulcis (Mill.) D. A. Webb Micrografts †


 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Micrografts
2.3. IAA Quantification
2.4. IAA Immunolocalization
2.5. Statistical Analysis
3. Results and Discussion
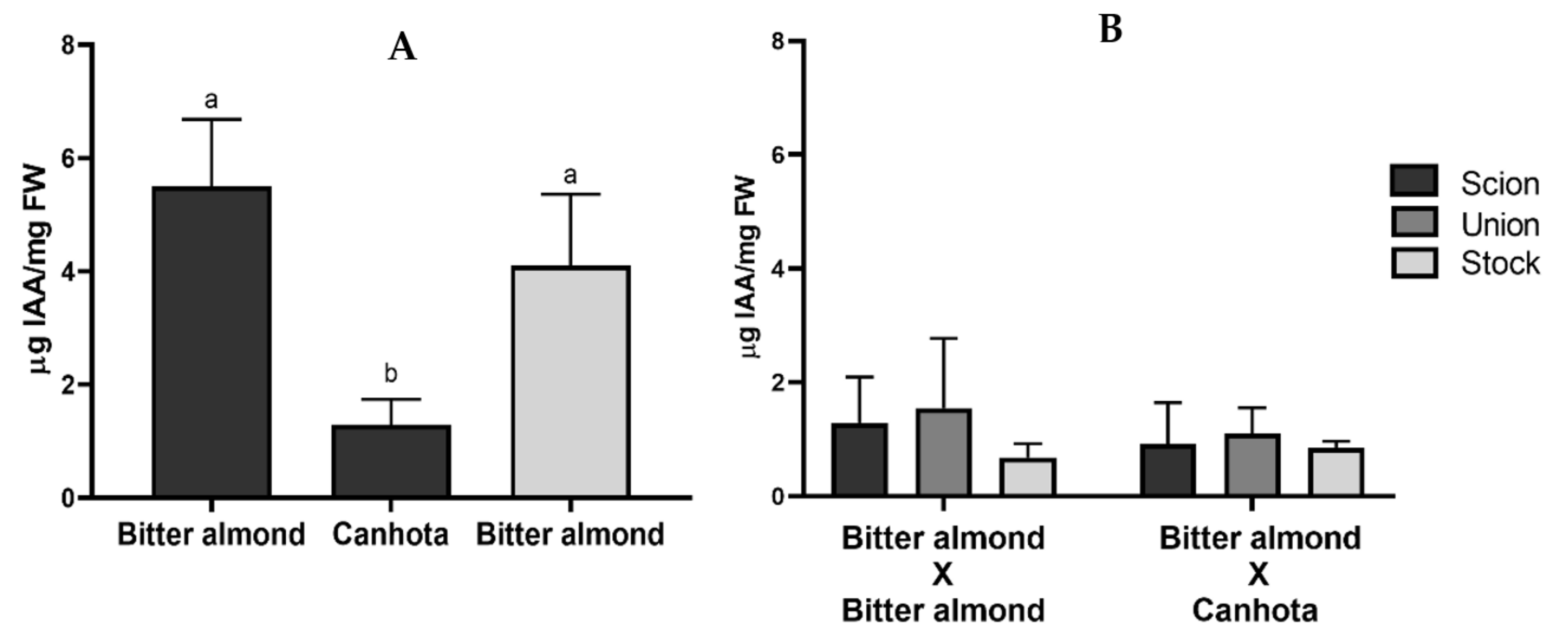
3.1. IAA Quantification
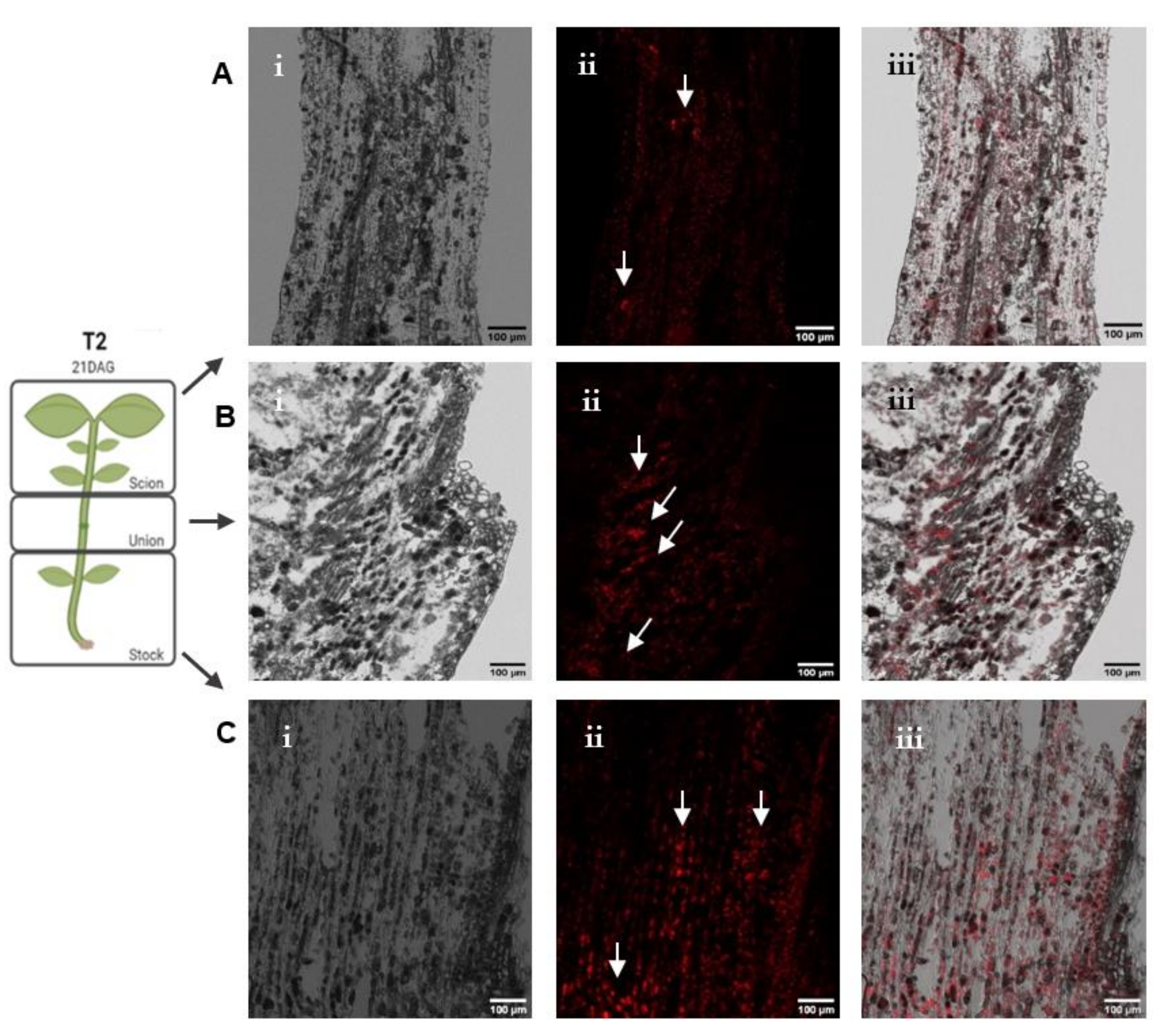
3.2. IAA Immunolocalization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguiar, C.; Pereira, J.A.; Arrobas, M.; Almeida, A.; Bento, A.; Cortés, I.L.; Rodrigues, N.; Rodrigues, M.Â.; Ribeiro, A.C.; Santos, S.A.P.; et al. Amendoeira: Estado de Produção; Centro Nacional de Competências dos Frutos Secos: Bragança, Portugal, 2017. [Google Scholar]
- Gautier, A.T.; Chambaud, C.; Brocard, L.; Ollat, N.; Gambetta, G.A.; Delrot, S.; Cookson, S.J. Merging genotypes: Graft union formation and scion/rootstock interactions. J. Exp. Bot. 2018, 70, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, C.W. Plant grafting: Insights into tissue regeneration. Regeneration 2016, 4, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in Rootstock−Scion interactions. Sci. Hortic. 2010, 127, 119–126. [Google Scholar] [CrossRef]
- Aloni, R. Differentiation of vascular tissues. Annu. Rev. Plant Physiol. 1987, 38, 179–204. [Google Scholar] [CrossRef]
- Yin, H.; Yan, B.; Sun, J.; Jia, P.; Zhang, Z.; Yan, X.; Chai, J.; Ren, Z.; Zheng, G.; Liu, H. Graft-union development: A delicate process that involves Cell−Cell communication between scion and stock for local auxin accumulation. J. Exp. Bot. 2012, 63, 4219–4232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, A.K.; Melnyk, C.W. The role of plant hormones during grafting. J. Plant Res. 2017, 131, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravana Kumar, R.M.; Gao, L.X.; Yuan, H.W.; Xu, D.B.; Liang, Z.; Tao, S.C.; Edqvist, J. Auxin enhances grafting success in Carya cathayensis (Chinese hickory). Planta 2017, 247, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhao, J.; Hu, F.; Qin, Y.; Wang, X.; Hu, G. Transcriptome changes between compatible and incompatible graft combination of Litchi chinensis by digital gene expression profile. Sci. Rep. 2017, 7, 3954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friml, J.; Palme, K. Polar auxin Transport–Old questions and new concepts? Plant Mol. Biol. 2002, 49, 273–284. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caeiro, S.; Lopes, T.; Pedrosa, A.; Caeiro, A.; Pires, R.C.; Faustino, A.; Rosa, A.; Marreiros, A.; Canhoto, J.; Marum, L.; et al. Quantification and Immunolocalization of Auxin in Prunus dulcis (Mill.) D. A. Webb Micrografts. Biol. Life Sci. Forum 2022, 11, 10. https://doi.org/10.3390/IECPS2021-11919
Caeiro S, Lopes T, Pedrosa A, Caeiro A, Pires RC, Faustino A, Rosa A, Marreiros A, Canhoto J, Marum L, et al. Quantification and Immunolocalization of Auxin in Prunus dulcis (Mill.) D. A. Webb Micrografts. Biology and Life Sciences Forum. 2022; 11(1):10. https://doi.org/10.3390/IECPS2021-11919
Chicago/Turabian StyleCaeiro, Sandra, Tércia Lopes, Ana Pedrosa, André Caeiro, Rita Costa Pires, Ana Faustino, Armindo Rosa, António Marreiros, Jorge Canhoto, Liliana Marum, and et al. 2022. "Quantification and Immunolocalization of Auxin in Prunus dulcis (Mill.) D. A. Webb Micrografts" Biology and Life Sciences Forum 11, no. 1: 10. https://doi.org/10.3390/IECPS2021-11919
APA StyleCaeiro, S., Lopes, T., Pedrosa, A., Caeiro, A., Pires, R. C., Faustino, A., Rosa, A., Marreiros, A., Canhoto, J., Marum, L., & Correia, S. (2022). Quantification and Immunolocalization of Auxin in Prunus dulcis (Mill.) D. A. Webb Micrografts. Biology and Life Sciences Forum, 11(1), 10. https://doi.org/10.3390/IECPS2021-11919