
Overexpression of Plant-Specific Insert from Cardosin B (PSI B) in Arabidopsis Correlates with Cell Responses to Stresses †



Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Stress Assays
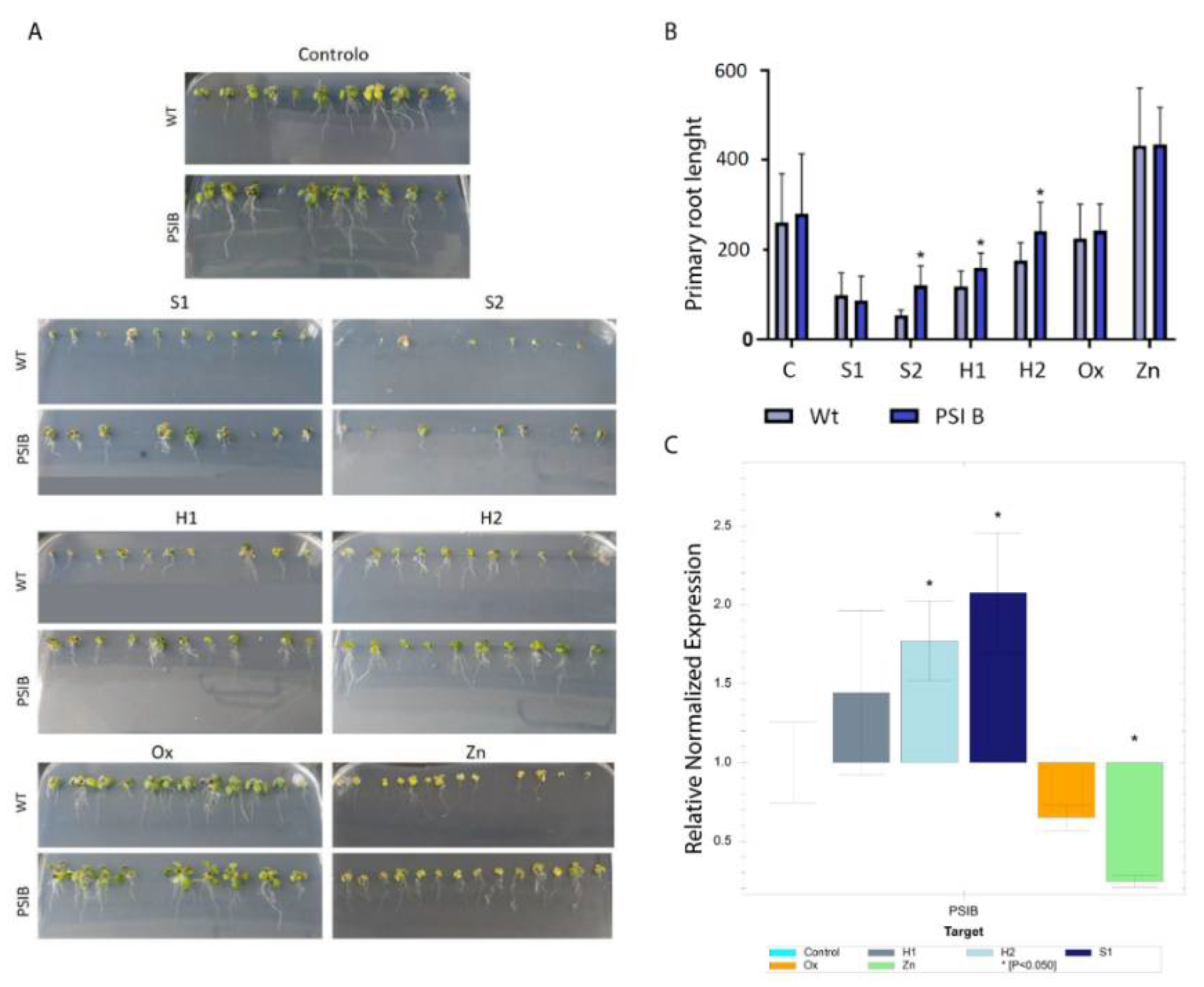
2.2. Root Biometrical Analysis
2.3. cDNA Preparation
2.4. Quantitative RT-PCR
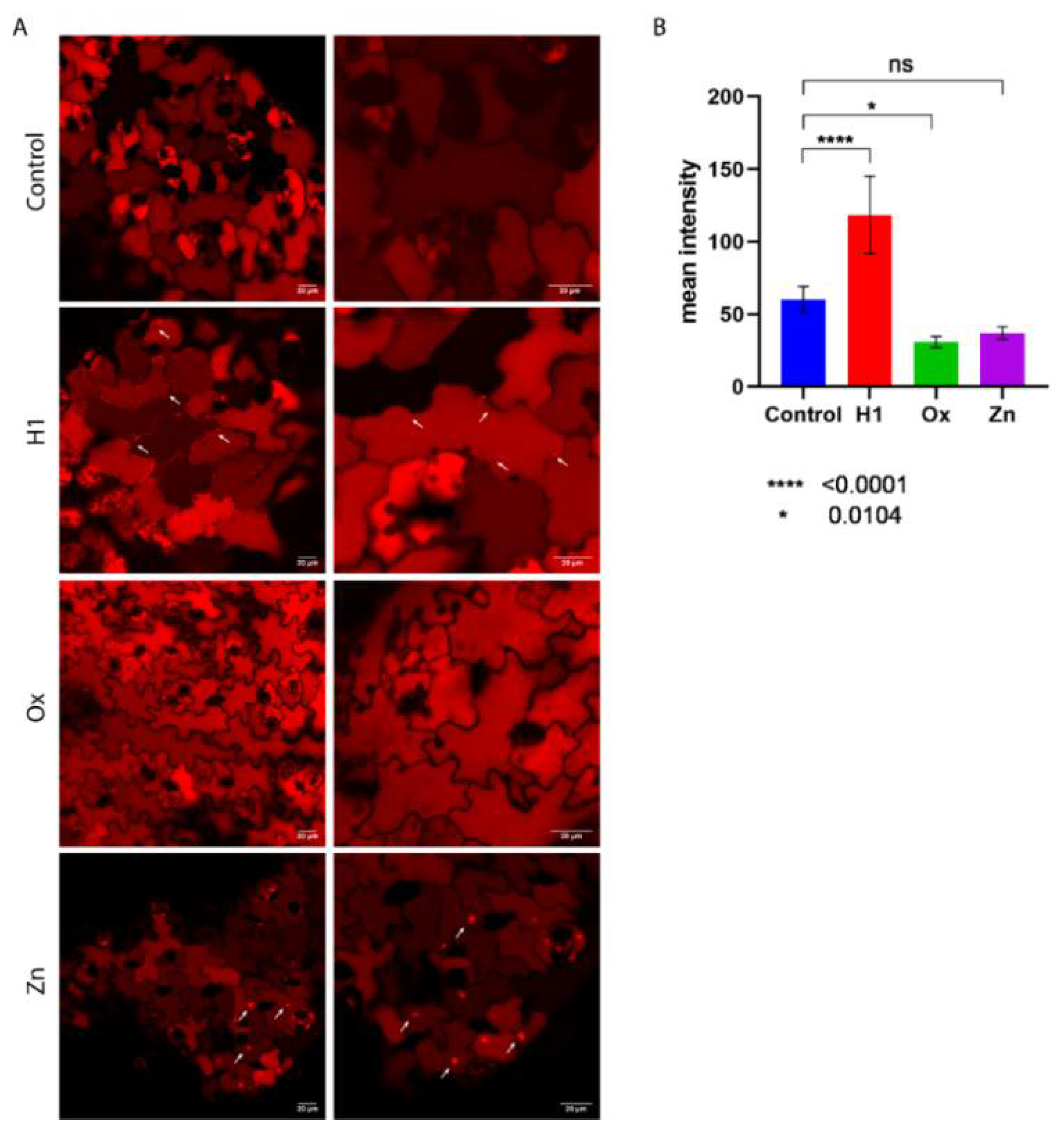
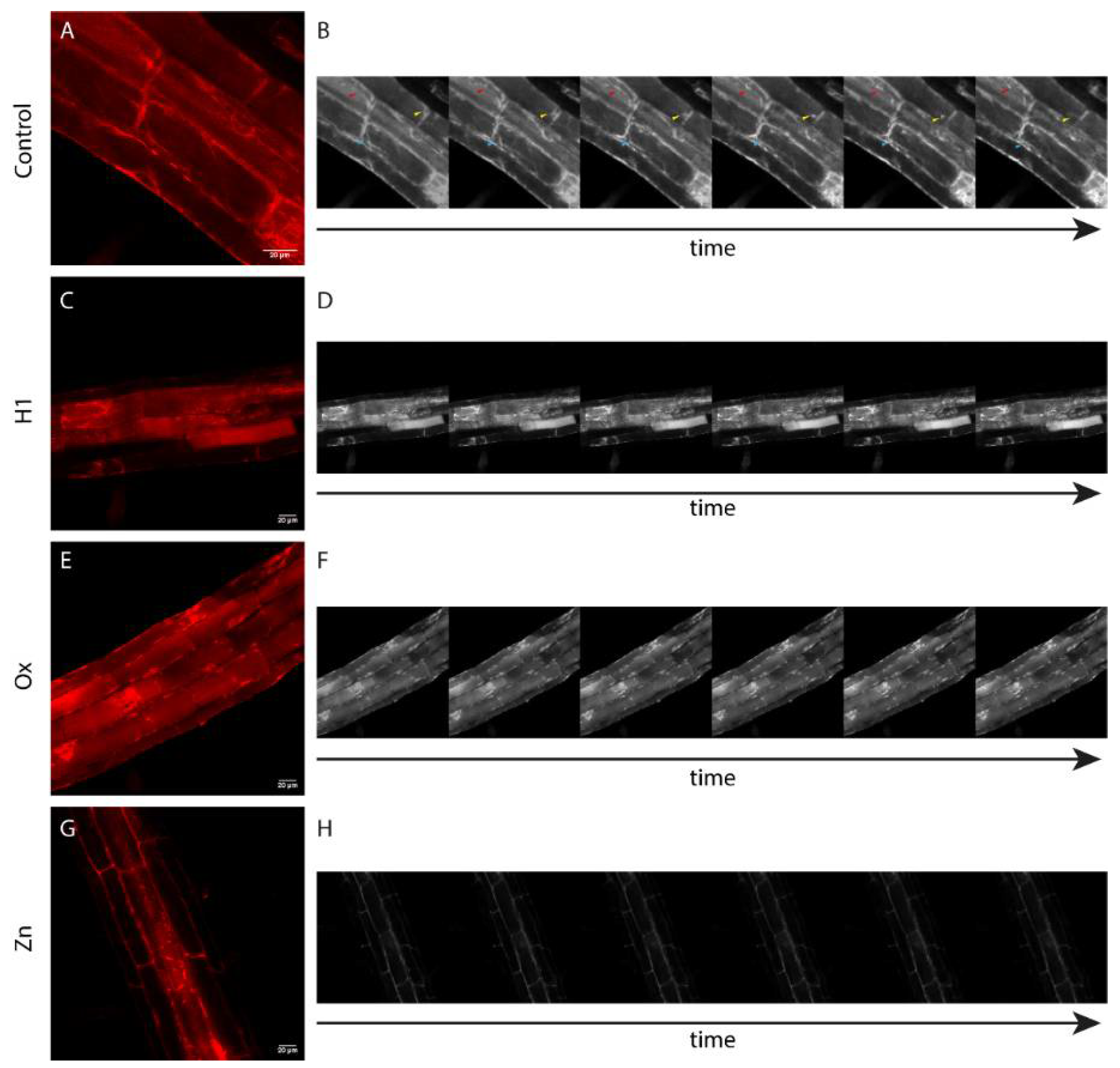
2.5. Confocal Microscopy
3. Results
3.1. PSI B Expression Changes in Plants under Abiotic Stress
3.2. PSI B Localization in Arabidopsis Plants under Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peleg, Z.; Apse, M.P.; Blumwald, E. Engineering Salinity and Water-Stress Tolerance in Crop Plants. Getting Closer to the Field; Academic Press: Cambridge, MA, USA, 2011; Volume 57. [Google Scholar]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigon, A.; Matos, A.R.; Laffray, D.; Zuily-Fodil, Y.; Pham-Thi, A.T. Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (Ecotype Columbia). Ann. Bot. 2004, 94, 345–351. [Google Scholar] [CrossRef]
- Sun, J.K.; Li, T.; Xia, J.B.; Tian, J.Y.; Lu, Z.H.; Wang, R.T. Influence of salt stress on ecophysiological parameters of Periploca sepium Bunge. Plant Soil Environ. 2011, 57, 139–144. [Google Scholar] [CrossRef]
- Scheuring, D.; Kleine-Vehn, J. On the discovery of an endomembrane compartment in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 10623–10624. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, M.; Gao, C.; Zeng, Y.; Cui, Y.; Shen, W.; Jiang, L. The roles of endomembrane trafficking in plant abiotic stress responses. J. Integr. Plant Biol. 2020, 62, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiong, Y.; Bassham, D.C. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy 2009, 5, 954–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signorelli, S.; Tarkowski, Ł.P.; Van den Ende, W.; Bassham, D.C. Linking Autophagy to Abiotic and Biotic Stress Responses. Trends Plant Sci. 2019, 24, 413–430. [Google Scholar] [CrossRef] [Green Version]
- Neves, J.; Sampaio, M.; Séneca, A.; Pereira, S.; Pissarra, J.; Pereira, C. Abiotic Stress Triggers the Expression of Genes Involved in Protein Storage Vacuole and Exocyst-Mediated Routes. Int. J. Mol. Sci. 2021, 22, 10644. [Google Scholar] [CrossRef] [PubMed]
- Faro, C.; Gal, S. Aspartic Proteinase Content of the Arabidopsis Genome. Curr. Protein Pept. Sci. 2005, 6, 493–500. [Google Scholar] [CrossRef]
- Kervinen, J.; Tormakangas, K.; Runeberg-Roos, P.; Guruprasad, K.; Blundell, T.; Teeri, T.H. Structure and possible function of aspartic proteinases in barley and other plants. In Proceedings of the Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 1995; Volume 362, pp. 241–254. [Google Scholar]
- Mutlu, A.; Susannah, G. Plant aspartic proteinases: Enzymes on the way to a function. Physiol. Plant. 1999, 105, 569–576. [Google Scholar] [CrossRef]
- Simões, I.; Faro, C. Structure and function of plant aspartic proteinases. Eur. J. Biochem. 2004, 271, 2067–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, L.K.Y.; Dupuis, J.H.; Dee, D.R.; Bryksa, B.C.; Yada, R.Y. Roles of Plant-Specific Inserts in Plant Defense. Trends Plant Sci. 2020, 25, 682–694. [Google Scholar] [CrossRef]
- Egas, C.; Lavoura, N.; Resende, R.; Brito, R.M.M.; Pires, E.; de Lima, M.C.P.; Faro, C. The Saposin-like Domain of the Plant Aspartic Proteinase Precursor Is a Potent Inducer of Vesicle Leakage. J. Biol. Chem. 2002, 275, 38190–38196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, F.F.; Mendieta, J.R.; Pagano, M.R.; Paggi, R.A.; Daleo, G.R.; Guevara, M.G. The swaposin-like domain of potato aspartic protease (StAsp-PSI) exerts antimicrobial activity on plant and human pathogens. Peptides 2010, 31, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Pereira, S.; Satiat-Jeunemaitre, B.; Pissarra, J. Cardosin A contains two vacuolar sorting signals using different vacuolar routes in tobacco epidermal cells. Plant J. 2013, 76, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Terauchi, K.; Asakura, T.; Ueda, H.; Tamura, T.; Tamura, K.; Matsumoto, I.; Misaka, T.; Hara-Nishimura, I.; Abe, K. Plant-specific insertions in the soybean aspartic proteinases, soyAP1 and soyAP2, perform different functions of vacuolar targeting. J. Plant Physiol. 2006, 163, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Vieira, V.; Peixoto, B.; Costa, M.; Pereira, S.; Pissarra, J.; Pereira, C. N-linked glycosylation modulates Golgi-independent vacuolar sorting mediated by the plant specific insert. Plants 2019, 8, 312. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.S.; da Costa, D.S.; Pereira, S.; de Moura Nogueira, F.; Albuquerque, P.M.; Teixeira, J.; Faro, C.; Pissarra, J. Cardosins in postembryonic development of cardoon: Towards an elucidation of the biological function of plant aspartic proteinases. Protoplasma 2008, 232, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R. Genome-Wide Identification and Testing of Superior Reference Genes for Transcript Normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Zhao, J.; Wang, X.; Guo, C.; Li, Z.; Wang, Y.; Wang, X. Constitutive expression of a grape aspartic protease gene in transgenic Arabidopsis confers osmotic stress tolerance. Plant Cell. Tissue Organ Cult. 2015, 121, 275–287. [Google Scholar] [CrossRef]
- Neves, J.; Séneca, A.; Pereira, S.; Pissarra, J.; Pereira, C. Abiotic Stress Upregulates the Expression of Genes Involved in PSV and Autophagy Routes. Biol. Life Sci. Forum 2021, 4, 40. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| UBQ_9 | TCACAATTTCCAAGGTGCTGC | TCACAATTTCCAAGGTGCTGC |
| SAND-1 | AACTCTATGCAGCATTTGATCCACT | TGATTGCATATCTTTATCGCCATC |
| m-Cherry | GACCACCTACAAGGCCAAG | GTGGGAGGTGATGTCCAACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, I.; Pereira, S.; Séneca, A.; Pissarra, J.; Pereira, C. Overexpression of Plant-Specific Insert from Cardosin B (PSI B) in Arabidopsis Correlates with Cell Responses to Stresses. Biol. Life Sci. Forum 2022, 11, 35. https://doi.org/10.3390/IECPS2021-11938
Moura I, Pereira S, Séneca A, Pissarra J, Pereira C. Overexpression of Plant-Specific Insert from Cardosin B (PSI B) in Arabidopsis Correlates with Cell Responses to Stresses. Biology and Life Sciences Forum. 2022; 11(1):35. https://doi.org/10.3390/IECPS2021-11938
Chicago/Turabian StyleMoura, Inês, Susana Pereira, Ana Séneca, José Pissarra, and Cláudia Pereira. 2022. "Overexpression of Plant-Specific Insert from Cardosin B (PSI B) in Arabidopsis Correlates with Cell Responses to Stresses" Biology and Life Sciences Forum 11, no. 1: 35. https://doi.org/10.3390/IECPS2021-11938
APA StyleMoura, I., Pereira, S., Séneca, A., Pissarra, J., & Pereira, C. (2022). Overexpression of Plant-Specific Insert from Cardosin B (PSI B) in Arabidopsis Correlates with Cell Responses to Stresses. Biology and Life Sciences Forum, 11(1), 35. https://doi.org/10.3390/IECPS2021-11938