
Antioxidant Properties of Chickpea (Cicer arietinum L.) Protein Hydrolysates: The In Vitro Evaluation of SOD Activity in THP-1 Cell Line †

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material and Reagent
2.2. Cell Culture and Treatment
2.3. Cell Cytotoxicity MTT Assay
2.4. Gene Expression Analysis and Cytokine Measurement
2.5. Nitric Oxide Measurement
2.6. Superoxide Dismutase Assay
2.7. Statistical Analysis
3. Results and Discussion
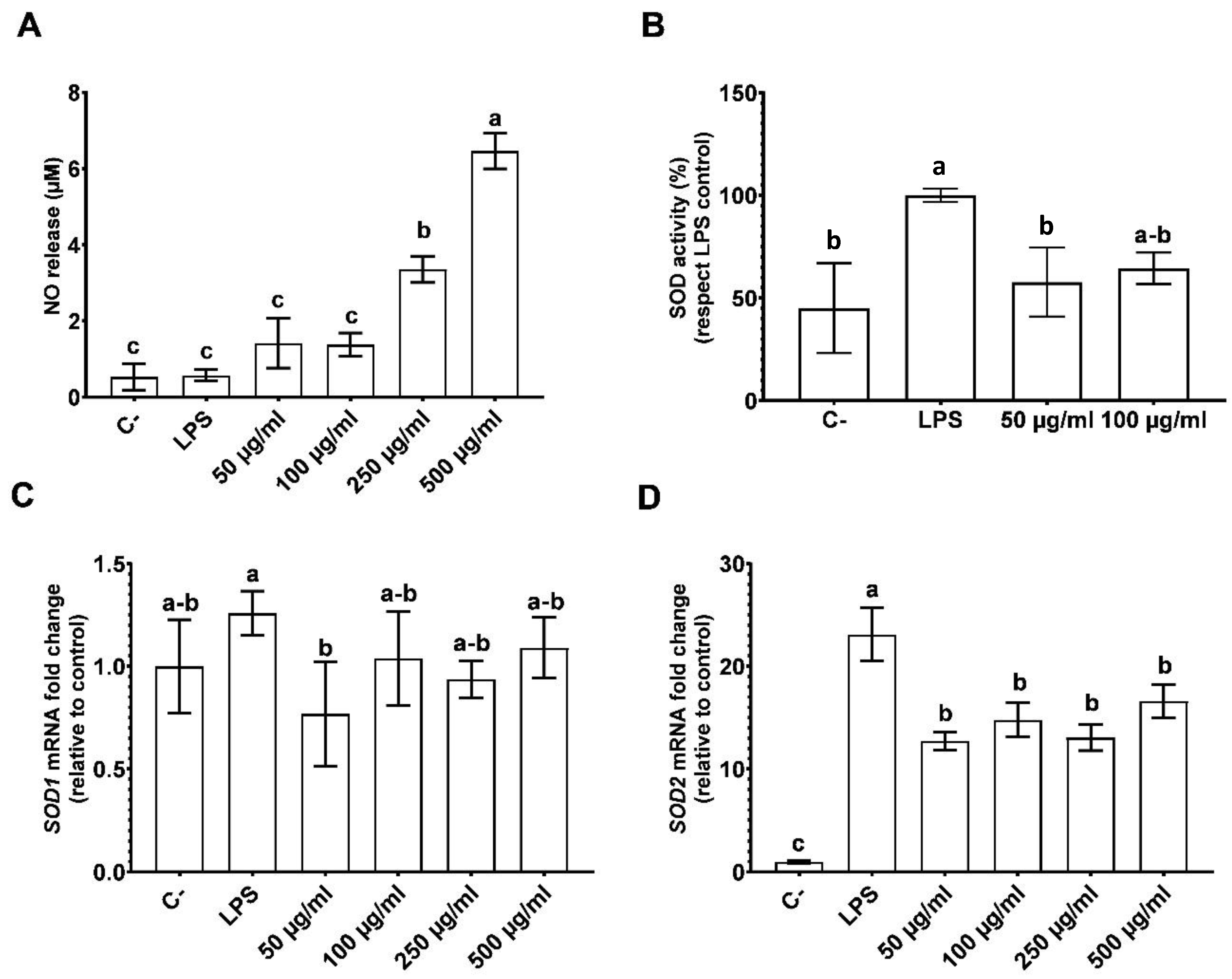
3.1. CPH Diminishing Inflammation in LPS Stimulated THP-1 Cells
3.2. The Effects of CPH on Oxidative Stress
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en (accessed on 1 August 2022).
- Boukid, F. Chickpea (Cicer arietinum L.) protein as a prospective plant-based ingredient: A review. Int. J. Food Sci. Technol. 2021, 56, 5435–5444. [Google Scholar] [CrossRef]
- Faridy, J.-C.M.; Stephanie, C.-G.M.; Gabriela, M.-M.O.; Cristian, J.-M. Biological Activities of Chickpea in Human Health (Cicer arietinum L.). A Review. Plant Foods Hum. Nutr. 2020, 75, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Duez, H.; Pourcet, B. Nuclear Receptors in the Control of the NLRP3 Inflammasome Pathway. Front. Endocrinol. 2021, 12, 630536. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios, E.C.S.; de Lima, T.M.; Moretti, A.I.S.; Soriano, F.G. The role of nitric oxide in the epigenetic regulation of THP-1 induced by lipopolysaccharide. Life Sci. 2016, 147, 110–116. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | NM Code | Forward Sequence | Reverse Sequence |
|---|---|---|---|
| IL-1β | NM_000576.2 | 5′-TCCTTCAGACACCCTCAACC-3′ | 5′-AGGCCCCAGTTTGAATTCTT-3′ |
| TNF-α | NM_000594.3 | 5′-CTGTCCTGCGTGTTGAAAGA-3′ | 5′-TTCTGCTTGAGAGGTGCTGA-3′ |
| SOD1 | NM_000454.5 | 5′-ACAAAGATGGTGTGGCCGAT-3′ | 5′-AACGACTTCCAGCGTTTCCT-3′ |
| SOD2 | NM_000636.4 | 5′-AACAACCTGAACGTCACCGA-3′ | 5′-CACGTTTGATGGCTTCCAGC-3′ |
| GADPH | NM_002046.6 | 5′-GAGTCAACGGATTTGGTCGT-3′ | 5′-GACAAGCTTCCCGTTCTCAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Martin, N.M.; Marquez, J.C.; Villanueva, Á.; Millán, F.; Millán-Linares, M.d.C.; Pedroche, J. Antioxidant Properties of Chickpea (Cicer arietinum L.) Protein Hydrolysates: The In Vitro Evaluation of SOD Activity in THP-1 Cell Line. Biol. Life Sci. Forum 2022, 17, 12. https://doi.org/10.3390/blsf2022017012
Rodríguez-Martin NM, Marquez JC, Villanueva Á, Millán F, Millán-Linares MdC, Pedroche J. Antioxidant Properties of Chickpea (Cicer arietinum L.) Protein Hydrolysates: The In Vitro Evaluation of SOD Activity in THP-1 Cell Line. Biology and Life Sciences Forum. 2022; 17(1):12. https://doi.org/10.3390/blsf2022017012
Chicago/Turabian StyleRodríguez-Martin, Noelia María, José Carlos Marquez, Álvaro Villanueva, Francisco Millán, María del Carmen Millán-Linares, and Justo Pedroche. 2022. "Antioxidant Properties of Chickpea (Cicer arietinum L.) Protein Hydrolysates: The In Vitro Evaluation of SOD Activity in THP-1 Cell Line" Biology and Life Sciences Forum 17, no. 1: 12. https://doi.org/10.3390/blsf2022017012
APA StyleRodríguez-Martin, N. M., Marquez, J. C., Villanueva, Á., Millán, F., Millán-Linares, M. d. C., & Pedroche, J. (2022). Antioxidant Properties of Chickpea (Cicer arietinum L.) Protein Hydrolysates: The In Vitro Evaluation of SOD Activity in THP-1 Cell Line. Biology and Life Sciences Forum, 17(1), 12. https://doi.org/10.3390/blsf2022017012