
Germination Kinetics of Ferula communis L. Seeds, a Potentially Multipurpose-Use Wild Species



Institute for BioEconomy, National Research Council of Italy, Via Paolo Gaifami 18, 95126 Catania, Italy
*
Author to whom correspondence should be addressed.
Seeds 2024, 3(2), 196-202; https://doi.org/10.3390/seeds3020015
Submission received: 29 February 2024
/
Revised: 2 April 2024
/
Accepted: 9 April 2024
/
Published: 14 April 2024
(This article belongs to the Special Issue Parameters of Seed Germination in Wild Plant Species)
Abstract
:Despite exhibiting intriguing features associated with its multipurpose applications and drought tolerance, Ferula communis remains a wild and uncultivated species, with limited experimental research on its biology, starting from seed germination and extending to its ecology. The purpose of this study was to investigate potential germination and kinetics in F. communis seeds in response to four cold stratification periods (0, 15, 45, and 90 days at a constant temperature of 5 °C) and four temperatures (5, 10, 15, and 20 °C) under continuous darkness. F. communis exhibited a pronounced germination potential exceeding 90%, with the optimal temperature for germination falling within the range of 5 °C to 15 °C, without necessitating cold stratification. A dramatic drop of the germination percentage was observed at 20 °C (<10%), suggesting a form of conditional dormancy attributed to the higher temperature tested.
1. Introduction
Ferula communis L. (giant fennel) is a perennial wild plant, belonging to the Apiaceae family, that grows naturally on the wasteland of the Mediterranean and Central Asia regions, blossoms in March and April, and goes dormant in early summer in dry climate conditions. It presents deep tap roots, clumps of leaves developing in winter and early spring, and a tall flowering stem (up to 3 m) with numerous yellow flowers clustered in umbels. Ferula communis has a fairly abundant production of seeds that varies in size and absolute weight of the seeds depending on the place of growth of the plant. Usually, they measure between 3 and 6 mm in length with an elongated and cylindrical shape, and a color that can vary, but they commonly appear dark brown or black. Seeds of the Ferula genus may exhibit dormancy, characterized by a hard seed coat that needs to be scarified or stratified to improve germination rates [1].
Ferula species have a long history of extract application in various medical and therapeutic contexts due to their well-documented significant biological activities. These extracts have been utilized in both human and veterinary practice to address a wide range of ailments, including headaches, digestive disorders, rheumatism, arthritis, and tumoral activity [2,3,4,5,6,7]. The biological activities of plant extracts from areal parts and roots were investigated in in vitro experiments involving several fungi, demonstrating fungitoxic effects on colony growth [8]. Historical records, dating back to the Roman period, document symbiotic relationships between Ferula species and other plants [9], notably with Pleurotus eryngii, a widely appreciated edible mushroom species [10]. Alternatively, some authors reported a toxic effect on animals and humans [11] and prenylcoumarin compounds were thought to be responsible for the toxic effects on sheep, goats, cattle, and horses [12]. Nevertheless, Arnoldi and coauthors [4] and subsequently Rubiolo and coauthors [13] differentiated two different chemotypes of F. communis, the ‘nonpoisonous’ and the ‘poisonous’, helping in interpreting the opposite effect and use described.
Traditional use of the stems is also reported for small furniture, but limited scientific studies are available on the properties of fibers. Ferula-plant-derived fillers have been used as an alternative reinforcing agent in composite materials due to its lightness, biodegradability, low cost, and renewability [14]. Recently, Touil and coauthors [15] investigated the potential integration of F. communis fiber into building materials to enhance their thermo-mechanical properties and reduce weight, thereby minimizing the utilization of landfills dedicated to this waste category. Potential use as an energy source was also proposed in bioethanol production [16].
All the aforementioned issues associated with F. communis suggest its potential valorization within the context of the new approach of the European Union’s common policy for agriculture and environment aimed at maintaining rural areas and landscapes, in which farmers are protectors and promoters not only of the agro-environment but also of the natural habitat including wild species. Due to its rapid growth, the ability to tolerate drought linked to the deep root system and xerophytic adaptations, the widespread distribution in wasteland characterizing many areas of the Mediterranean basin, and its versatile use, F. communis appears as an interesting wild species worthy of study.
Despite the recognition of its multipurpose potential, F. communis largely remains an undomesticated and uncultivated species with limited experimental research on its biology starting from seed germination and its ecology. The germination traits of diverse accessions within the Ferula genus were examined, revealing complex germination cues characterized by the presence of dormancy mechanisms that inhibit germination until the requirement for cold stratification is fulfilled [17,18,19,20,21].
With this consideration in mind, we conducted an experimental trial on the seed germination of F. communis L., specifically focusing on germinability and its kinetics, to evaluate the optimal seed germination temperature and the effects of cold stratification in terms of duration.
2. Materials and Methods
2.1. Seed Material and Collection Site
We identified and collected mature seeds of F. communis from Etna Mountain (Pedara, 700 m asl—37°37′25″ N, 15°03′24.19″ E) during the late summer of 2023. The collection area is classified as Csa (warm temperate, dry and hot summer) following the Köppen and Geiger climate classification, with 14.4 °C as the average annual temperature and an annual precipitation of 595 mm.
In the region where the seeds were collected, the physiological maturity of seeds begins in late spring, and their natural dispersal occurs during the summer and early autumn periods, characterized by acropetal kinetics of the composed umbels. In late July 2023, seeds were collected from disseminating flowers, selecting bottom simple umbels. Immediately after collection, seeds were air-dried for 1–2 days and then stored at room temperature (20 ± 2 °C) in paper bags until the start of the germination experiment. Immediately preceding with the germination test, seed surface disinfection was performed by immersing them in a 5% sodium hypochlorite (NaOCl) solution for 5 min, followed by two rinses with demineralized sterile water. No fungicide treatment was performed to avoid possible interference of the active ingredient on germination.
2.2. Germination Test
The experimental treatments were as follows: firstly, seeds were subjected to four different levels of cold stratification (0, 15, 45, and 90 days) at a constant temperature of 5 °C and a relative humidity of 70% (±2%); then, seeds were maintained in a cabinet germinator (KW-Apparecchi Scientifici, Italy) at four distinct temperature levels (5, 10, 15, 20 °C) in a condition of continuous darkness. The experiments were arranged in a completely randomized design with three replications per thesis. Each replication involved thirty seeds arranged in Petri dishes containing double layered Whatman No. 1 filter paper, moistened with 5 mL of distilled water. Sterilized–distilled water (maintained at the same treatment temperature) was added as required to ensure non-limiting moisture conditions for germination.
Germinated seeds were counted and removed every 24 h for 40 days. Seeds were considered germinated when a primary radicle protrusion 2 mm long was observed, according to the guidelines of the International Seed Testing Association [21].
2.3. Statistical Analysis
At the end of the trial, the final germination percentage (FGP) was computed. A factorial 2-way (cold stratification × temperature) ANOVA model was performed. When ANOVA indicated a significant effect, differences among the combination of treatments were tested with the Tukey multiple comparison test (HSD test) using the CoStat 6.4 statistics software package. Percentage data were transformed in arcsen before the statistical analysis, whereas data included in the tables represent original values.
To study seed germination kinetics, we plotted the cumulated number of germinated seeds over time and fitted the derived data modifying the three-parameter logistic curve proposed by P. F. Verhulst (Verhulst 1838, 1845) as follows:
y = a/(1 + exp(−(x − x0)/b))
This function allowed the direct biological interpretation of germination behavior and generates cumulative estimates of seed germination.
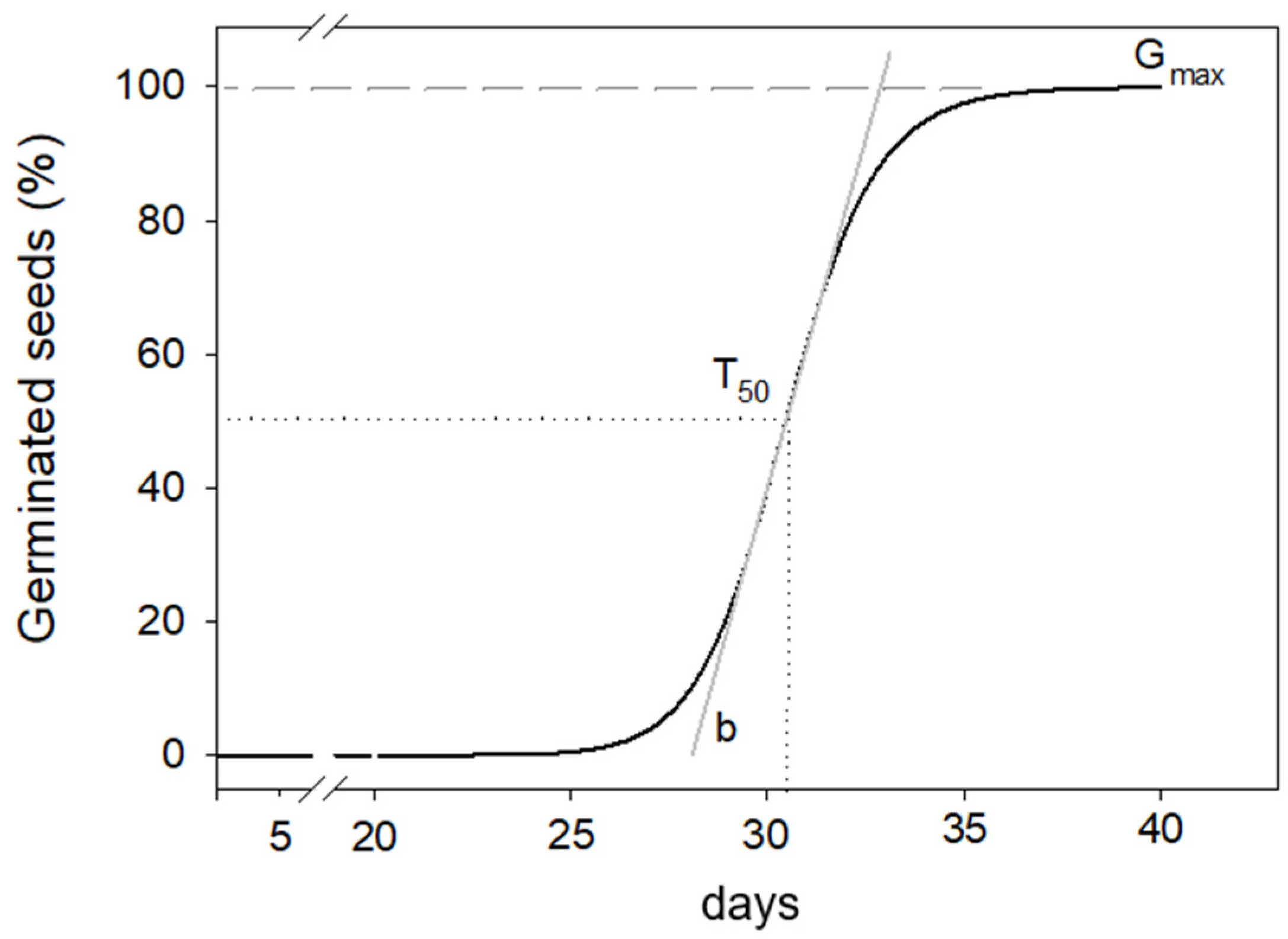
where Gcum(t) is the cumulative percentage of germination at time t (days); Gmax is the asymptotic final germination percentage value at t → +∞; t50 is the inflection point where Gcum equals half of Gmax, representing the time, in days, to reach 50% of germinated seeds; and b is the slope, a dimensionless “shape factor,” which primarily controls the steepness of the germination curve (Figure 1).
The obtained curves were compared, applying the extra sum of the square F test. When the null hypothesis was rejected, the best-fit values of each obtained parameter (Gmax, t50, and b) were compared with a t test using GraphPad 6.0 (GraphPad Software Inc., San Diego, CA, USA):
where the numerator is the difference between best-fit values of the target parameter for two specific theses and the denominator is an estimate of the standard error of that difference, computed as the square root of the sum of the squares of the two standard error values.
The three parameters’ sigmoid curve chosen for this experiment, and widely adopted for a germination test, is affected by the lack of information on some key phases of the germination process. Based on the experimental dataset, we computed the following intervals: (i) the initial stage encompassing imbibition and the lag phase (IS-LP), extending from the experiment’s onset to the date of the first seed germination, and (ii) the interval between the end of the lag phase and the date when the last seed germinated (LP-LSG), indirectly inferred through the dimensionless “shape factor” b. The ANOVA analysis was performed on obtained data.
3. Results
The analysis of variance for the final germination percentages (Table 1) showed significant differences exclusively due to temperature. The comparison of the mean values for the different levels elucidated that the observed result was mainly related to the germination failure observed under the highest temperature (10% of germinated seeds at 20 °C).
Conversely, 94% of germinated seeds were counted, averaging the remaining temperature levels (Table 2).
Considering the final germination percentage (FGP) as the principal variable in a germinability assessment and noting the absence of a discernible response to stratification for FGP, the investigation of germination kinetics has focused exclusively on temperature-related data. Furthermore, the low number of seeds germinated at the highest level of temperature (20 °C) conflicts with the biological interpretation of the sigmoid curve parameters. Consequently, the curve interpolation was restricted to datasets corresponding to temperatures of 5, 10, and 15 °C.
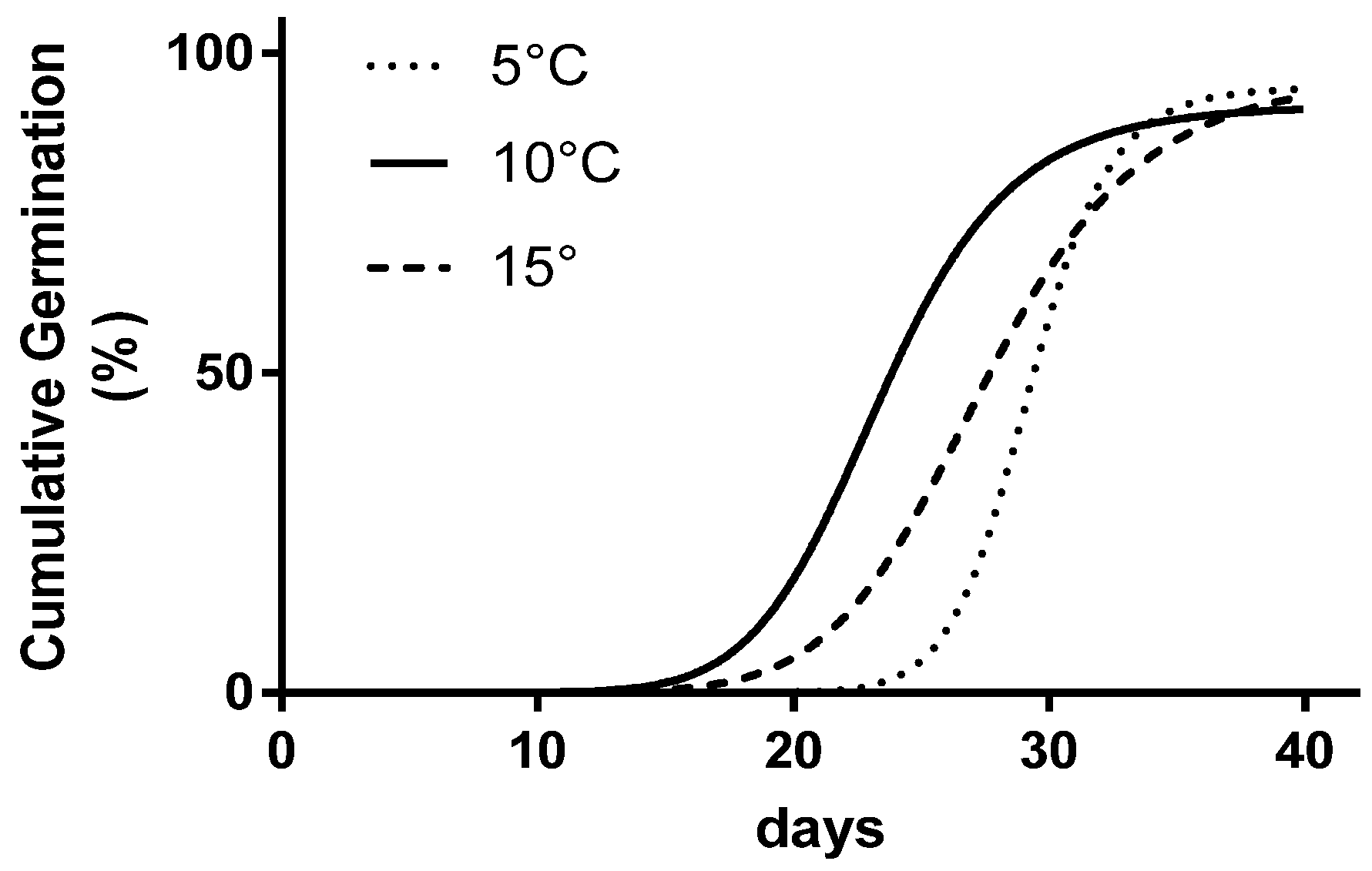
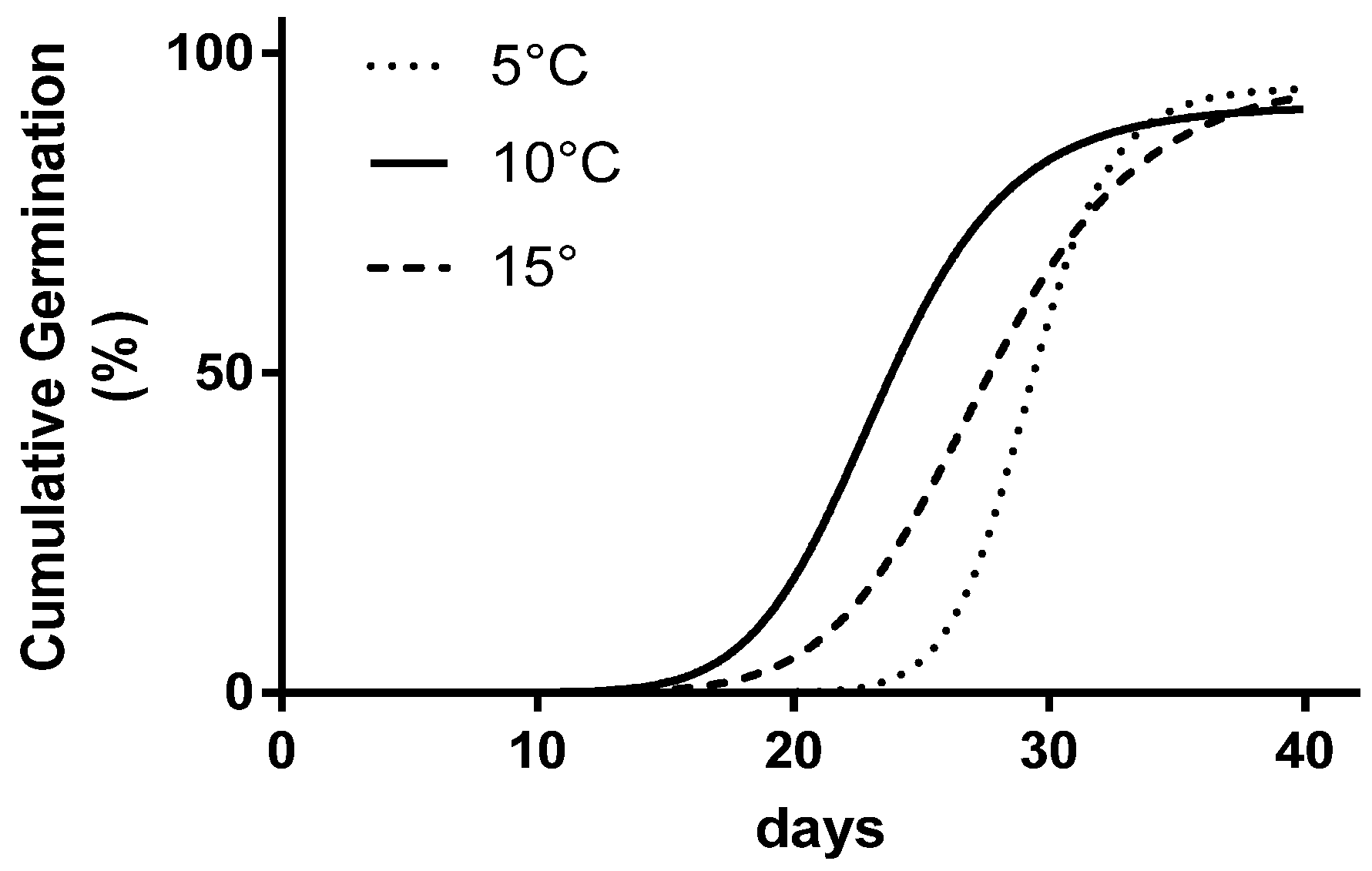
The extra sum of the square F test showed statistical differences between the interpolated curves for the three temperature levels (Figure 2).
From the analysis of the single parameters of each curve (Table 3), no differences emerged comparing the asymptote Gmax values (94.4%), whereas for T50, the extreme values were achieved at 10 °C (23.4 days) and 5 °C (29.2 days), respectively. The lowest temperature obtained the highest b (18.4), twofold higher when compared to the other levels, and the longest duration of the initial stage.
4. Discussion and Conclusions
Cold stratification is a vital mechanism for various species in temperate regions, where winters are cold, ensuring the timely cessation of dormancy when favorable conditions for seedling emergence are established after the winter season [22,23]. In the Mediterranean climate condition, characterized by mild/wet winters and hot/dry summers, the temperature plays a critical ecological role, governing the germination dynamics, ensuring that seeds remain dormant during the hot and dry summer months, thereby synchronizing their germination with the cooler and moister conditions, typically experienced during the winter season.
Our results emphasize that the F. communis exhibits high germination potential (>90%) within an optimal temperature range of 5 to 15 °C and does not require cold stratification for germination. However, it germinates at significantly lower percentages at 20 °C (10%), indicating a kind of conditional dormancy. These conditionally dormant seeds became nondormant the following winter, when lower temperatures and increased moisture levels signal the ideal condition for germination and seedling emergence [24,25,26].
Limited researchers addressed the germination of the wild genus of Ferula, whereas many authors observed primary seed dormancy in the Apiaceae family and reported a positive effect of cold stratification on germination [27,28,29,30]. In particular, on the Ferula genus, Nikolaeva [1] reported a cold stratification requirement of some species for germination and the effective temperature for this requirement was 0–3 °C. On the contrary, Aghilian and coauthors [30] stated that pre-chilling had no effect on seeds of Ferula gummosa Boiss dormancy breaking. Even fewer researchers focused on F. communis, and contrasting results have been obtained. Ari and coauthors [31] observed higher germination in F. tingitana L. (60%) than F. communis (3%) and hypothesized that F. communis might have seed dormancy and need prechilling treatment, but no specific trials have been described in their paper. On the contrary, Sanna and coauthors (2009, cit. by Dettori et al. [32]) reported a high germination percentage of F. communis (higher than 80%) in a temperature range of 10–15 °C without prechilling treatment. Our experiment’s findings, involving an even lower temperature (5 °C), provided further support for this hypothesis. Additionally, our analysis of germination kinetics revealed a specific impact of the coldest temperature, in particular on the duration of the period when radicle protrusion is observed. The significantly higher value of b calculated at 5 °C, reflecting the interval between the initial and final primary radicle protrusions, suggests a higher synchronicity of the protrusion phase at lower temperature. This result should be stated even though the ANOVA, conducted on the duration of the LP-LSG interval in response to germination temperatures, failed to define this difference (Table 2), likely due to the large variation observed in the different replications of 15 °C treatment. In addition, the dramatic drop in the germination percentage observed at 20 °C represents, as well, a relevant result of our experiment. Assuming the optimum range of temperature usually imposed in germination trials (20–25 °C), the sensitivity stated in our study could explain the very low values reached in many of the studies concerning this species or related genus. In addition, the positive effect on the germination of priming treatment reported by several authors in Ferula spp. suggests the opportunity to evaluate if the high percentage of non-germinated seeds observed at 20 °C remains viable in a soil seed bank for the following season.
In summary, our research provides valuable insights into managing Ferula communis L. seed germination, crucial for biodiversity conservation under the Mediterranean climatic conditions. However, it is imperative that future studies delve deeper into elucidating the underlying mechanisms governing seed germination activities, as well as the associated physiological processes. Such investigations aim to devise strategies for enhancing and standardizing germination rates.
Author Contributions
Conceptualization, E.R., M.D., G.A. and S.B.; methodology, E.R., M.D., G.A. and S.B.; investigation, E.R., M.D. and G.A.; data curation, E.R., M.D. and G.A.; writing—original draft preparation, E.R., M.D., G.A. and S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article —no new data generated.
Acknowledgments
We thank Alessandra Pellegrino and Salvatore La Rosa for their support in bibliographic research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nikolaeva, M.G. Physiology of Deep Dormancy in Seeds; Shapiro, Z., Translator; National Science Foundation: Washington, DC, USA, 1969. (In Russian)
- Al-Yahya, M.A.; Muhammad, I.; Mirza, H.H.; El-Ferali, F.S. Antibacterial constituents from the rhizomes of Ferula communis. Phytother. Res. 1998, 12, 335–339. [Google Scholar] [CrossRef]
- Chen, B.; Teranishi, R.; Kawazoe, K.; Takaishi, Y.; Honda, G.; Itoh, M.; Takeda, Y.; Kodzhimatoy, O.K. Sesquiterpenoides from Ferula kuhistanica. Phytochemistry 2000, 54, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Arnoldi, L.; Ballero, M.; Fuzzati, N.; Maxia, A.; Mercalli, E.; Pagni, L. HPLC-DAD-MS identification of bioactive secondary metabolites from Ferula communis roots. Fitoterapia 2004, 75, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Akaberi, M.; Iranshahy, M.; Iranshahi, M. Review of the traditional uses, phytochemistry, pharmacology and toxicology of giant fennel (Ferula communis L. subsp. communis). Iran. J. Basic Med. Sci. 2015, 18, 1050. [Google Scholar] [PubMed]
- Deniz, G.Y.; Laloglu, E.; Koc, K.; Geyikoglu, F. Hepatoprotective potential of Ferula communis extract for carbon tetrachloride induced hepatotoxicity and oxidative damage in rats. Biotech. Histochem. 2019, 94, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Kavaz, D.; Faraj, R.E. Investigation of composition, antioxidant, antimicrobial and cytotoxic characteristics from Juniperus sabina and Ferula communis extracts. Sci. Rep. 2023, 13, 7193. [Google Scholar] [CrossRef] [PubMed]
- Mamoci, E.; Cavoski, I.; Simeone, V.; Mondelli, D.; Al-Bitar, L.; Caboni, P. Chemical composition and in vitro activity of plant extracts from Ferula communis and Dittrichia viscosa against postharvest fungi. Molecules 2011, 22, 2609–2625. [Google Scholar] [CrossRef] [PubMed]
- Buller, R. The fungus lore of the Greeks and Romans. Trans. Br. Mycol. Soc. 1914, 5, 21–66. [Google Scholar] [CrossRef]
- Zervakis, G.I.; Balis, C. A pluralistic approach in the study of Pleurotus species with emphasis on compatibility and physiology of the European morphotaxa. Mycol. Res. 1996, 6, 717–731. [Google Scholar] [CrossRef]
- Marchi, A.; Appendino, G.; Pirisi, I.; Ballero, M.; Loi, M.C. Genetic differentiation of two distinct chemotypes of Ferula communis (Apiaceae) in Sardinia (Italy). Biochem. Syst. Ecol. 2003, 31, 1397–1408. [Google Scholar] [CrossRef]
- Appendino, G.; Tagliapietra, S.; Gariboldi, P.; Nano, G.M.; Picci, V. w-Oxygenated prenylated coumarins from F. communis. Phytochemistry 1988, 27, 3619–3624. [Google Scholar] [CrossRef]
- Rubiolo, P.; Matteodo, M.; Riccio, G.; Ballero, M.; Christen, P.; Fleury-Souverain, S.; Veuthey, J.L.; Bicchi, C. Analytical discrimination of poisonous and nonpoisonous chemotypes of Giant Fennel (Ferula communis L.) through their biologically active and volatile fractions. J. Agric. Food Chem. 2006, 54, 7556–7563. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Sarikanat, M.; Sever, K.; Durmuşkahya, C. Extraction and properties of Ferula communis (chakshir) fibers as novel reinforcement for composites materials. Compos. B Eng. 2013, 44, 517–523. [Google Scholar] [CrossRef]
- Touil, M.; Rahmoun, O.; Iken, O.; El Harti, K.; Saadani, R.; Rahmoune, M. Exploring the thermal behaviour and thermo-mechanical properties of Ferula communis reinforced plaster and mortar composites: An integrated experimental and numerical approach. Energy Convers. Manag. 2023, 289, 117119. [Google Scholar] [CrossRef]
- Polycarpou, P. Ethanol production from Ferula communis. Biomass Bioenergy 2012, 36, 289–292. [Google Scholar] [CrossRef]
- Zare, A.R.; Solouki, M.; Omidi, M.; Irvani, N.; Abasabadi, A.O.; Nezad, N.M. Effect of various treatments on seed germination and dormancy breaking in Ferula assa foetida L. (Asafetida), a threatened medicinal herb. Trakia J. Sci. 2011, 9, 57–61. [Google Scholar]
- Suran, D.; Bolor, T.; Bayarmaa, G.A. In Vitro seed germination and callus induction of Ferula ferulaeoides (Steud.) Korov. (Apiaceae). Mong. J. Biol. Sci. 2016, 14, 53–58. [Google Scholar]
- Nowruzian, A.; Masoumian, M.; Ebrahimi, A.; Bakhshi Khaniki, G.H. Effect of Breaking Dormancy Treatments on Germination of Ferula assa foetida L. Seeds. Iran. J. Seed Res. 2017, 3, 154–169. [Google Scholar] [CrossRef]
- Salehi Shanjani, P.; Falah Hoseini, L. Germination Responses of Ferula assa-foetida and Ferula gummosa Boiss. Seeds to Continuous Cold Stratification. J. Med. Plants Prod. 2022, 11, 9–16. [Google Scholar]
- ISTA. International rules for seed testing. Seed Sci. Technol. 1993, 21, 160–186. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution Dormancy and Germination, 2nd ed.; Academic: San Diego, CA, USA, 2014. [Google Scholar]
- Cavieres, L.A.; Sierra-Almeida, A. Assessing the importance of cold-stratification for seed germination in alpine plant species of the High-Andes of central Chile. Perspect. Plant Ecol. 2018, 30, 125–131. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, C.C.; Baskin, J.M. A graphical method for identifying the six types of non-deep physiological dormancy in seeds. Plant Biol. 2017, 19, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Cristaudo, A.; Catara, S.; Mingo, A.; Restuccia, A.; Onofri, A. Temperature and storage time strongly affect the germination success of perennial Euphorbia species in Mediterranean regions. Ecol. Evol. 2019, 9, 10984–10999. [Google Scholar] [CrossRef] [PubMed]
- Carruggio, F.; Onofri, A.; Catara, S.; Impelluso, C.; Castrogiovanni, M.; Lo Cascio, P.; Cristaudo, A. Conditional Seed Dormancy Helps Silene hicesiae Brullo & Signor. Overcome Stressful Mediterranean Summer Conditions. Plants. 2021, 10, 2130. [Google Scholar] [PubMed]
- Baskin, C.C.; Mayer, S.E.; Baskin, J.M. Two type of morphophysiological dormancy in seeds of two genera (Osmorhiza and Erythorium) with an Arcto-Tertiary distribution pattern. Am. J. Bot. 1995, 82, 293–298. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Okagami, N. Seed germination echophysiology of Asian species Osmorhiza aristata (Apiaceae): Comparison with its North American cogenera and implications for evolution of type of dormancy. Am. J. Bot. 2002, 89, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Nadjafi, F.; Bannayan, M.; Tabrizi, L.; Rastgoo, M. Seed germination and dormancy breaking techniques for Ferula gummosa and Teucrium polium. J. Arid. Environ. 2006, 64, 542–547. [Google Scholar] [CrossRef]
- Aghilian, S.; Khajeh-Hosseini, M.; Anvarkhah, S. Evaluation of seed dormancy in forty medicinal plant species. Int. J. Agric. Crop Sci. 2014, 7, 760–768. [Google Scholar]
- Ari, E.; Gürbüz, E.; Tuğrul, A.S. Seed germinations of 20 wild species growing in Antalya (Turkey) with outdoor ornamental plant potential. Fifth International Scientific Agricultural Symposium. Agrosym 2014, 2014, 439–445. [Google Scholar]
- Dettori, C.A.; Loi, M.C.; Brullo, S.; Arguimbau, P.F.; Tamburini, E.; Bacchetta, G. The genetic diversity and structure of the Ferula communis L. complex (Apiaceae) in the Tyrrhenian area. Flora 2016, 223, 138–146. [Google Scholar]
Figure 1.
Three parameters’ sigmoid curve chosen to interpolate percentage of germinated seeds over time.
Figure 1.
Three parameters’ sigmoid curve chosen to interpolate percentage of germinated seeds over time.
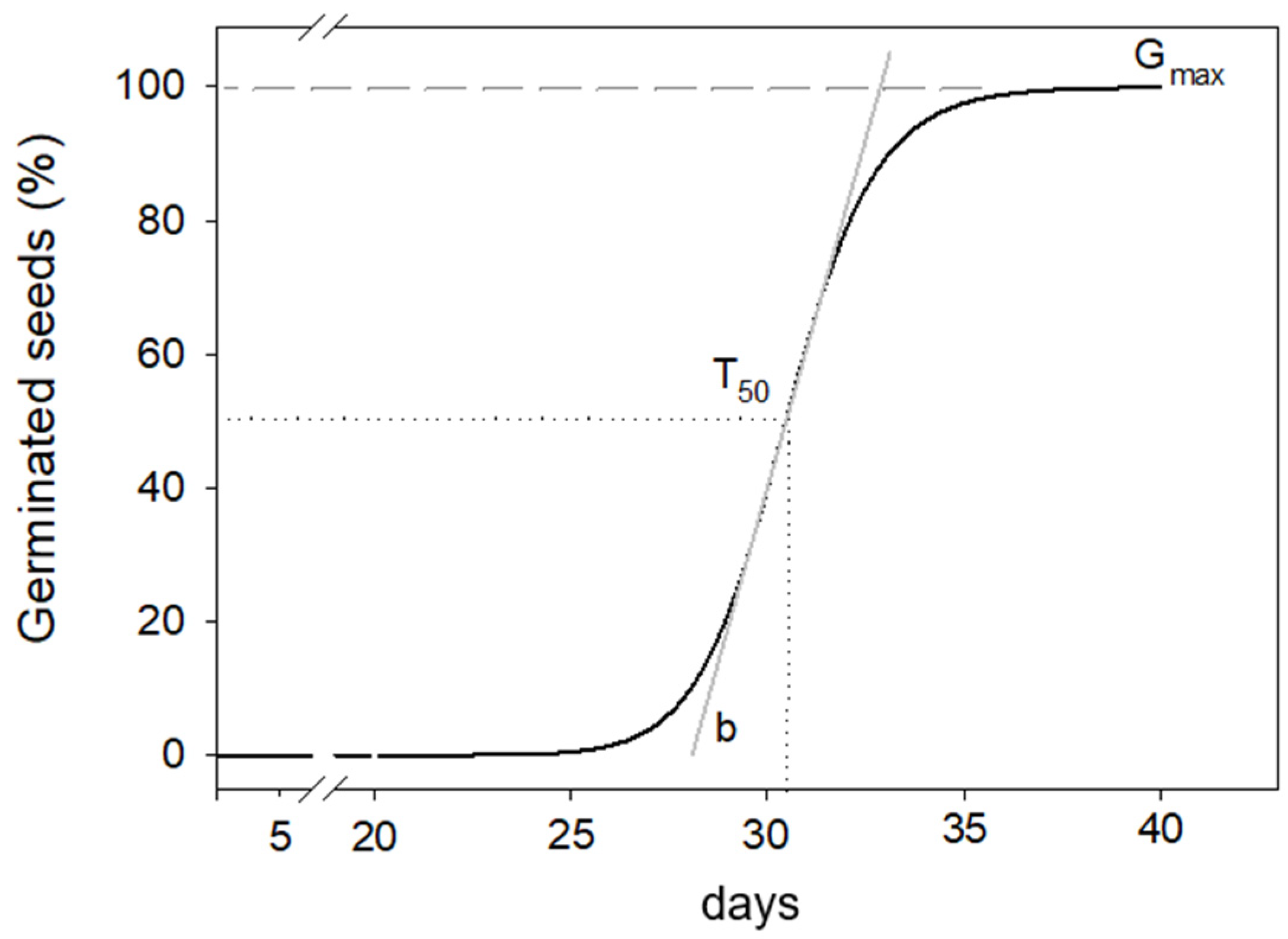
Figure 2.
The comparison of three parameters’ sigmoid curves fitting the three datasets (5, 10, and 15 °C) where seed germination kinetics has been studied.
Figure 2.
The comparison of three parameters’ sigmoid curves fitting the three datasets (5, 10, and 15 °C) where seed germination kinetics has been studied.

{kind=link}
{kind=link}
Table 1.
Result of two-way ANOVA for the effects of cold stratification and temperature.
| Source of Variation | FGP | IS-LP | LP-LSG | |||
|---|---|---|---|---|---|---|
| F Values | p | F Values | p | F Values | p | |
| Cold Stratification (CS) | 1.67 | ns | n.d. | n.d. | ||
| Temperature (T) | 113.14 | *** | 7.22 | * | 7.22 | ns |
| Interaction (CS × T) | 0.87 | ns | n.d. | n.d. | ||
*, *** Significant at the 0.01 and 0.001 probability level, respectively; ns = not significant; n.d. = not detected. FGP: final germination percentage; IS-LP: duration of the initial stage (imbibition and lag phase); LP-LSG: duration of the interval between the date when the last seed germinated and the end of the lag phase.
Table 2.
Final germination percentage and phase durations computed from experimental dataset of Ferula communis seeds.
Table 2.
Final germination percentage and phase durations computed from experimental dataset of Ferula communis seeds.
| Temperature | FGP | IS-LP | LP-LSG |
|---|---|---|---|
| % | days | days | |
| 5 °C | 96.7 a | 27.0 a | 27.0 |
| 10 °C | 90.0 a | 18.0 b | 18.0 |
| 15 °C | 96.7 a | 20.0 b | 20.0 |
| 20 °C | 10.0 b | -- | -- |
Means followed by the different letters in each column are significantly different based on the Tukey test at the 0.05 probability level. FGP: final germination percentage; IS-LP: duration of the initial stage (imbibition and lag phase); LP-LSG: duration of the interval between the date when the last seed germinated and the end of the lag phase.
Table 3.
Sigmoid curve parameters (mean ± standard error).
| Temperature | Gmax | T50 | b | |||
|---|---|---|---|---|---|---|
| % | s.e | days | s.e | s.e | ||
| 5 °C | 94.7 | ±1.67 | 29.2 | ±0.15 a | 18.4 | ±1.46 a |
| 10 °C | 91.9 | ±1.98 | 23.4 | ±0.26 b | 9.1 | ±0.80 b |
| 15 °C | 96.6 | ±2.97 | 27.5 | ±0.33 ab | 8.9 | ±0.73 b |
Gmax is the asymptotic final germination percentage value; t50 is the inflection point where the cumulative percentage of germination equals half of Gmax; b is the slope, a dimensionless “shape factor”. Means followed by the different letters in each column are significantly different based on the Tukey test at the 0.05 probability level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Distefano, M.; Avola, G.; Berti, S.; Riggi, E. Germination Kinetics of Ferula communis L. Seeds, a Potentially Multipurpose-Use Wild Species. Seeds 2024, 3, 196-202. https://doi.org/10.3390/seeds3020015
AMA Style
Distefano M, Avola G, Berti S, Riggi E. Germination Kinetics of Ferula communis L. Seeds, a Potentially Multipurpose-Use Wild Species. Seeds. 2024; 3(2):196-202. https://doi.org/10.3390/seeds3020015
Chicago/Turabian StyleDistefano, Miriam, Giovanni Avola, Stefano Berti, and Ezio Riggi. 2024. "Germination Kinetics of Ferula communis L. Seeds, a Potentially Multipurpose-Use Wild Species" Seeds 3, no. 2: 196-202. https://doi.org/10.3390/seeds3020015