
Visual Analytics for Robust Investigations of Placental Aquaporin Gene Expression in Response to Maternal SARS-CoV-2 Infection
 , ,
, ,
Abstract
1. Introduction
2. Methods
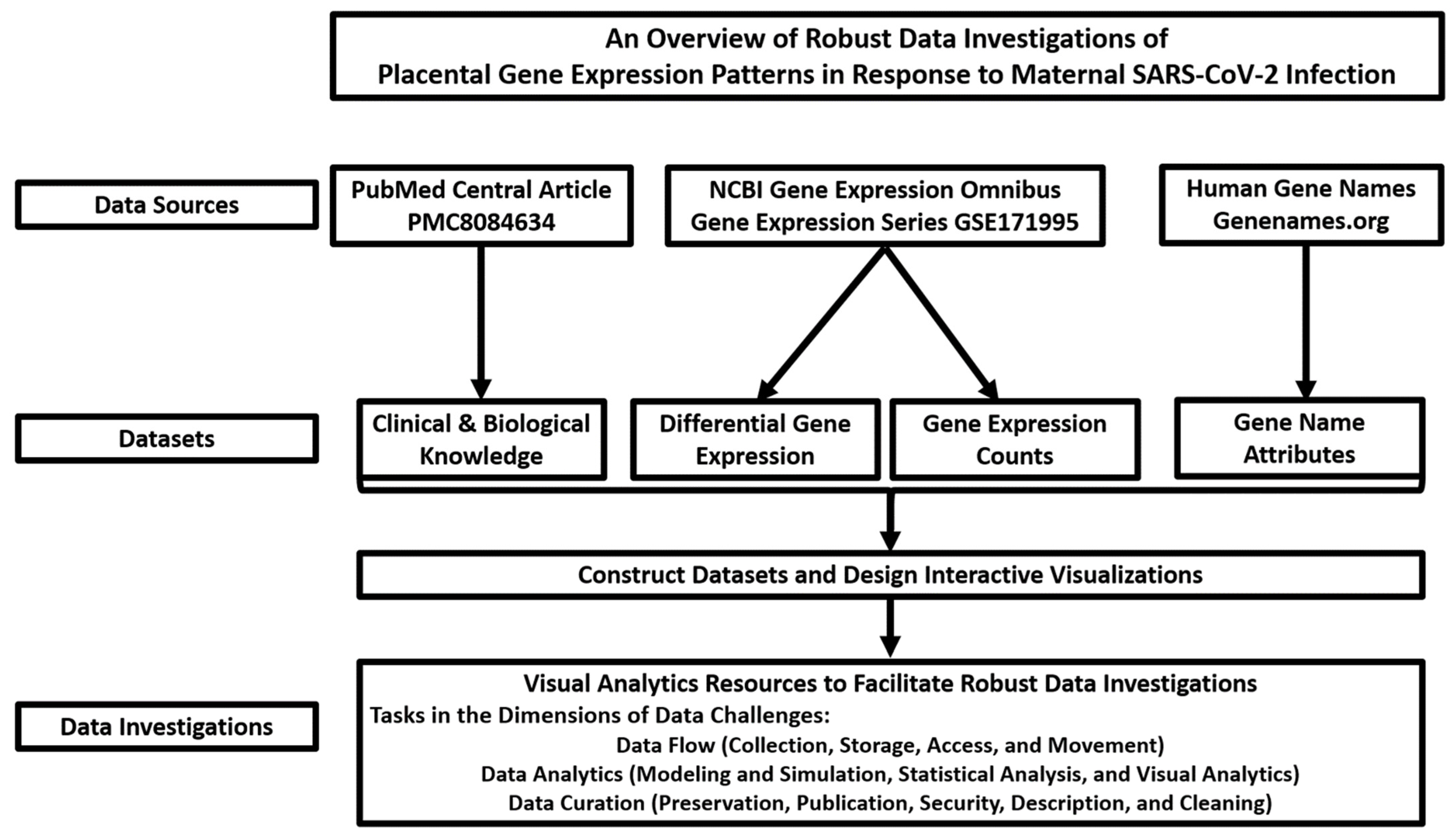
2.1. Data Sources and Overview of Robust Data Investigations of Placental Gene Expression Patterns in Response to Maternal SARS-CoV-2 Infection
2.2. Construction of Value-Added Datasets
2.3. Design and Implementation of Visual Analytics Resources
2.4. Verification of Accuracy of Visual Analytics Resources Content and Gene Family Expression Use Cases
2.5. Searches for Biological Networks and Pathways associated with Gene Sets
3. Results
3.1. Constructed Datasets
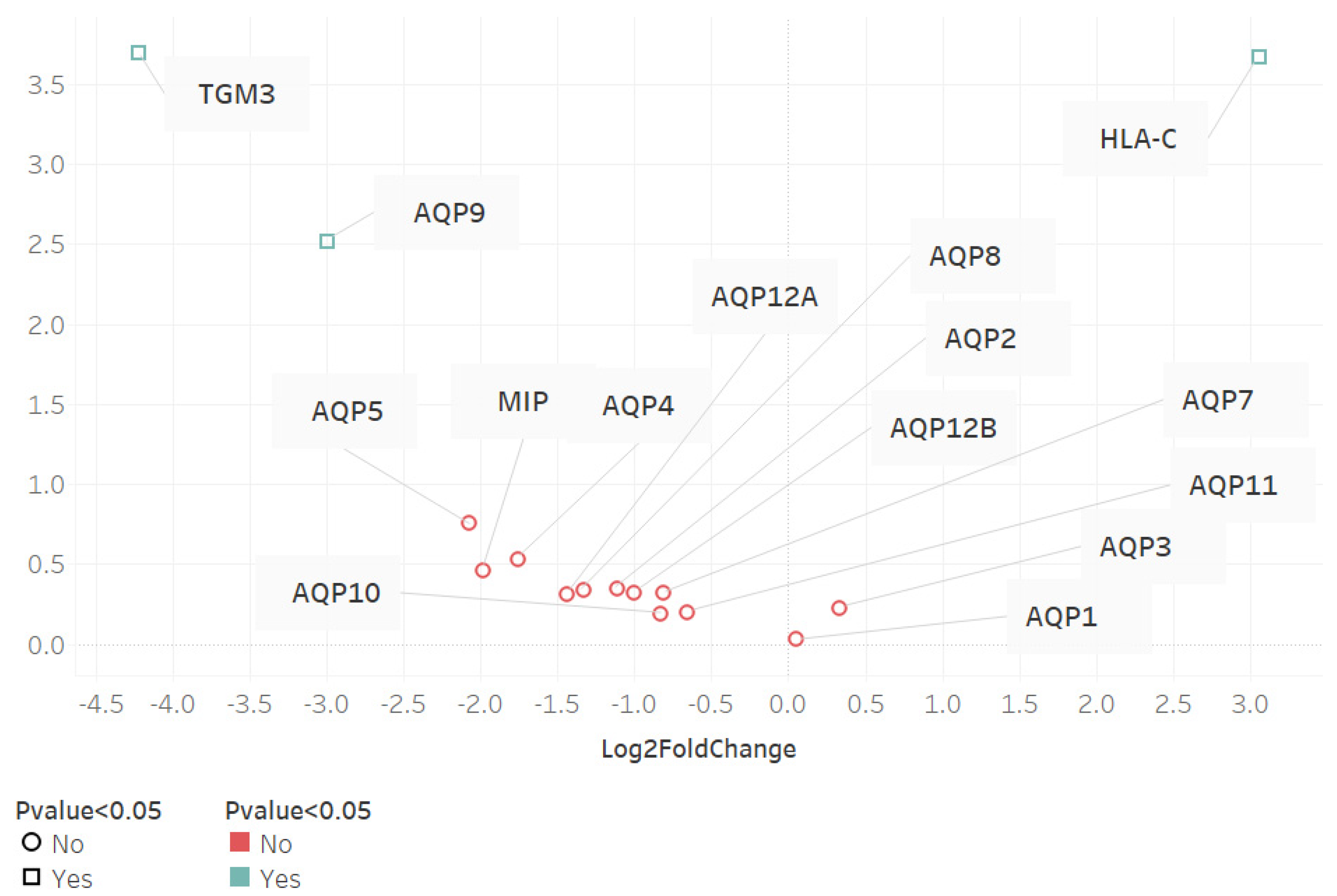
3.2. Volcano Plot of Differential Gene Expression
3.3. Combination of Gene Expression Box Plots and Data Table of Placenta Samples according to Sex of Infant, SARS-CoV-2 Infection Status, and Severity of SARS-CoV-2 Infection
3.4. Data Table of Genes with Gene Expression Values Grouped by Binary Number Patterns
3.5. Combination of Heatmap and Box Plot of Gene Expression Levels
3.6. Biological Networks and Pathways associated with Gene Sets
- Inflammatory response to SARS-CoV-2 (IFNA8, IFNA13, IFNW1, NOX1, and TLR9)
- Immune response to SARS-CoV-2 (CSF2, IFNA13, and REN)
- Linoleic acid metabolism affected by coronavirus infection (ACOT2)
- Pathogenesis of SARS-CoV-2 mediated by nsp9-nsp10 complex (IGLL1)
- SARS-CoV-2 host-pathogen interaction (NUP210)
- Iron metabolism in placenta (HEPHL1)
- SARS-CoV-2 signaling pathway (HP, ITIH4, and SERPINA10)
- Microglia pathogen phagocytosis (TREM1)
- Hematopoietic stem cell differentiation (CSF2)
- Pregnane X receptor pathway (GSTA2)
- Prolactin signaling pathway
- Glycolysis and gluconeogenesis (PGK2)
3.7. Visual Analytics Resources for Robust Investigations of Placental Gene Expression Data in Response to Maternal SARS-CoV-2 Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Griffiths, S.K.; Campbell, J.P. Placental structure, function and drug transfer. Contin. Educ. Anaesth. Crit. Care Pain 2015, 15, 84–89. [Google Scholar] [CrossRef]
- Guttmacher, A.E.; Maddox, Y.T.; Spong, C.Y. The Human Placenta Project: Placental structure, development, and function in real time. Placenta 2014, 35, 303–304. [Google Scholar] [CrossRef]
- Ortega, M.A.; Fraile-Martínez, O.; García-Montero, C.; Sáez, M.A.; Álvarez-Mon, M.A.; Torres-Carranza, D.; Álvarez-Mon, M.; Bujan, J.; García-Honduvilla, N.; Bravo, C. The pivotal role of the placenta in normal and pathological pregnancies: A focus on preeclampsia, fetal growth restriction, and maternal chronic venous disease. Cells 2022, 11, 568. [Google Scholar] [CrossRef] [PubMed]
- Mate, A.; Reyes-Goya, C.; Santana-Garrido, Á.; Sobrevia, L.; Vázquez, C.M. Impact of maternal nutrition in viral infections during pregnancy. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166231. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Hu, X.; Cao, B. Viral infections during pregnancy: The big challenge threatening maternal and fetal health. Matern. Fetal Med. 2022, 4, 72–86. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, F.; Peng, Y.; Chen, R.; Zhou, W.; Wang, H.; OuYang, J.; Yu, B.; Xu, Z. Transcriptomic profiling of human placenta in gestational diabetes mellitus at the single-cell level. Front. Endocrinol. 2021, 12, 679582. [Google Scholar] [CrossRef]
- Sood, R.; Zehnder, J.L.; Druzin, M.L.; Brown, P.O. Gene expression patterns in human placenta. Proc. Natl. Acad. Sci. USA 2006, 103, 5478–5483. [Google Scholar] [CrossRef]
- Saben, J.; Zhong, Y.; McKelvey, S.; Dajani, N.K.; Andres, A.; Badger, T.M.; Gomez-Acevedo, H.; Shankar, K. A comprehensive analysis of the human placenta transcriptome. Placenta 2014, 35, 125–131. [Google Scholar] [CrossRef]
- Gheorghe, C.P.; Goyal, R.; Mittal, A.; Longo, L.D. Gene expression in the placenta: Maternal stress and epigenetic responses. Int. J. Dev. Biol. 2010, 54, 507. [Google Scholar] [CrossRef]
- Martinez, V.D.; Cohn, D.E.; Telkar, N.; Minatel, B.C.; Pewarchuk, M.E.; Marshall, E.A.; Price, E.M.; Robinson, W.P.; Lam, W.L. Profiling the small non-coding RNA transcriptome of the human placenta. Sci. Data 2021, 8, 166. [Google Scholar] [CrossRef]
- Gray, K.A.; Seal, R.L.; Tweedie, S.; Wright, M.W.; Bruford, E.A. A review of the new HGNC gene family resource. Hum. Genom. 2016, 10, 6. [Google Scholar] [CrossRef]
- Cox, B.; Leavey, K.; Nosi, U.; Wong, F.; Kingdom, J. Placental transcriptome in development and pathology: Expression, function, and methods of analysis. Am. J. Obstet. Gynecol. 2015, 213, S138–S151. [Google Scholar] [CrossRef]
- Ilieva, M.; Tschaikowski, M.; Vandin, A.; Uchida, S. The current status of gene expression profilings in COVID-19 patients. Clin. Transl. Discov. 2022, 2, e104. [Google Scholar] [CrossRef]
- Sacha, D.; Stoffel, A.; Stoffel, F.; Kwon, B.C.; Ellis, G.; Keim, D.A. Knowledge generation model for visual analytics. IEEE Trans. Vis. Comput. Graph. 2014, 20, 1604–1613. [Google Scholar] [CrossRef]
- Rutter, L.; Moran Lauter, A.N.; Graham, M.A.; Cook, D. Visualization methods for differential expression analysis. BMC Bioinform. 2019, 20, 458. [Google Scholar] [CrossRef]
- Ludt, A.; Ustjanzew, A.; Binder, H.; Strauch, K.; Marini, F. Interactive and reproducible workflows for exploring and modeling RNA-seq data with pcaExplorer, Ideal, and GeneTonic. Curr. Protoc. 2022, 2, e411. [Google Scholar] [CrossRef]
- Lu-Culligan, A.; Chavan, A.R.; Vijayakumar, P.; Irshaid, L.; Courchaine, E.M.; Milano, K.M.; Tang, Z.; Pope, S.D.; Song, E.; Vogels, C.B. Maternal respiratory SARS-CoV-2 infection in pregnancy is associated with a robust inflammatory response at the maternal-fetal interface. Med 2021, 2, 591–610.e510. [Google Scholar] [CrossRef]
- Zhao, F.; Tallarek, A.-C.; Wang, Y.; Xie, Y.; Diemert, A.; Lu-Culligan, A.; Vijayakumar, P.; Kittmann, E.; Urbschat, C.; Bayo, J. A unique maternal and placental galectin signature upon SARS-CoV-2 infection suggests galectin-1 as a key alarmin at the maternal–fetal interface. Front. Immunol. 2023, 14, 1196395. [Google Scholar] [CrossRef]
- Casula, M.; Rangarajan, N.; Shields, P. The potential of working hypotheses for deductive exploratory research. Qual. Quant. 2021, 55, 1703–1725. [Google Scholar] [CrossRef]
- Ahalt, S.; Bedard, D.; Carsey, T.; Crabtree, J.; Green, K.; Jeffries, C.; Knowles, D.; Kum, H.; Lander, H.; Nassar, N. Establishing a National Consortium for Data Science; Renaissance Computing Institute, University of North Carolina at Chapel Hill: Chapel Hill, NC, USA, 2012. [Google Scholar]
- Isokpehi, R.D.; Johnson, M.O.; Campos, B.; Sanders, A.; Cozart, T.; Harvey, I.S. Knowledge visualizations to inform decision making for improving food accessibility and reducing obesity rates in the United States. Int. J. Environ. Res. Public Health 2020, 17, 1263. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Johnson, C.P.; Tucker, A.N.; Gautam, A.; Brooks, T.J.; Johnson, M.O.; Cozart, T.; Wathington, D.J. Integrating datasets on public health and clinical aspects of sickle cell disease for effective community-based research and practice. Diseases 2020, 8, 39. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Simmons, S.S.; Johnson, M.O.; Payton, M. Genomic evidence for bacterial determinants influencing obesity development. Int. J. Environ. Res. Public Health 2017, 14, 345. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Valero, K.C.W.; Graham, B.E.; Pacurari, M.; Sims, J.N.; Udensi, U.K.; Ndebele, K. Secondary data analytics of aquaporin expression levels in glioblastoma stem-like cells. Cancer Inform. 2015, 14, 95–103. [Google Scholar] [CrossRef]
- Makolo, A.U.; Smile, O.; Ezekiel, K.B.; Destefano, A.M.; McCall, J.L.; Isokpehi, R.D. Leveraging H3Africa scholarly publications for technology-enhanced personalized bioinformatics education. Educ. Sci. 2022, 12, 859. [Google Scholar] [CrossRef]
- Sedig, K.; Parsons, P. Interaction design for complex cognitive activities with visual representations: A pattern-based approach. AIS Trans. Hum. Comput. Interact. 2013, 5, 84–133. [Google Scholar] [CrossRef]
- King, L.S.; Agre, P. Pathophysiology of the aquaporin water channels. Annu. Rev. Physiol. 1996, 58, 619–648. [Google Scholar] [CrossRef] [PubMed]
- Isokpehi, R.D.; Rajnarayanan, R.V.; Jeffries, C.D.; Oyeleye, T.O.; Cohly, H.H. Integrative sequence and tissue expression profiling of chicken and mammalian aquaporins. BMC Genom. 2009, 10, S7. [Google Scholar] [CrossRef] [PubMed]
- Day, R.E.; Kitchen, P.; Owen, D.S.; Bland, C.; Marshall, L.; Conner, A.C.; Bill, R.M.; Conner, M.T. Human aquaporins: Regulators of transcellular water flow. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 1492–1506. [Google Scholar] [CrossRef] [PubMed]
- Sha, X.-y.; Xiong, Z.-f.; Liu, H.-s.; Di, X.-d.; Ma, T.-h. Maternal-fetal fluid balance and aquaporins: From molecule to physiology. Acta Pharmacol. Sin. 2011, 32, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; He, H.; Liu, H.; Liu, Q.; Zhang, L.; Liu, B.; Sugimoto, K.; Wu, Q. Aquaporin-1, a new maternally expressed gene, regulates placental development in the mouse. Biol. Reprod. 2016, 95, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Alejandra, R.; Natalia, S. The blocking of aquaporin-3 (AQP3) impairs extravillous trophoblast cell migration. Biochem. Biophys. Res. Commun. 2018, 499, 227–232. [Google Scholar] [CrossRef]
- Liu, H.; Wintour, E.M. Aquaporins in development—A review. Reprod. Biol. Endocrinol. 2005, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Jiang, W.; Zhang, W.; Shen, Q.; Chen, M.; Zhu, X. Expression and significance of aquaporins during pregnancy. Front. Biosci.-Landmark 2013, 18, 1373–1383. [Google Scholar]
- Szpilbarg, N.; Castro-Parodi, M.; Reppetti, J.; Repetto, M.; Maskin, B.; Martinez, N.; Damiano, A.E. Placental programmed cell death: Insights into the role of aquaporins. Mol. Hum. Reprod. 2016, 22, 46–56. [Google Scholar] [CrossRef]
- Galán-Cobo, A.; Ramírez-Lorca, R.; Echevarría, M. Role of aquaporins in cell proliferation: What else beyond water permeability? Channels 2016, 10, 185–201. [Google Scholar] [CrossRef]
- Ishibashi, K.; Tanaka, Y.; Morishita, Y. The role of mammalian superaquaporins inside the cell: An update. Biochim. Biophys. Acta 2021, 1863, 183617. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Delporte, C. Involvement of aquaglyceroporins in energy metabolism in health and disease. Biochimie 2021, 188, 20–34. [Google Scholar] [CrossRef]
- Wagner, K.; Unger, L.; Salman, M.M.; Kitchen, P.; Bill, R.M.; Yool, A.J. Signaling mechanisms and pharmacological modulators governing diverse aquaporin functions in human health and disease. Int. J. Mol. Sci. 2022, 23, 1388. [Google Scholar] [CrossRef]
- Burton, G.J.; Cindrova-Davies, T.; wa Yung, H.; Jauniaux, E. Hypoxia and reproductive health: Oxygen and development of the human placenta. Reproduction 2021, 161, F53–F65. [Google Scholar] [CrossRef]
- Castro-Parodi, M.; Szpilbarg, N.; Dietrich, V.; Sordelli, M.; Reca, A.; Abán, C.; Maskin, B.; Farina, M.; Damiano, A.E. Oxygen tension modulates AQP9 expression in human placenta. Placenta 2013, 34, 690–698. [Google Scholar] [CrossRef]
- Elahi, S. Hematopoietic responses to SARS-CoV-2 infection. Cell. Mol. Life Sci. 2022, 79, 187. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2021, 49, D10. [Google Scholar] [CrossRef]
- Tweedie, S.; Braschi, B.; Gray, K.; Jones, T.E.; Seal, R.L.; Yates, B.; Bruford, E.A. Genenames.org: The HGNC and VGNC resources in 2021. Nucleic Acids Res. 2021, 49, D939–D946. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef] [PubMed]
- Glusman, G.; Caballero, J.; Robinson, M.; Kutlu, B.; Hood, L. Optimal scaling of digital transcriptomes. PLoS ONE 2013, 8, e77885. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Sun, T.; Song, D.; Li, J.J. Statistics or biology: The zero-inflation controversy about scRNA-seq data. Genome Biol. 2022, 23, 31. [Google Scholar] [CrossRef] [PubMed]
- Beard, L.; Aghassibake, N. Tableau (version 2020.3). J. Med. Libr. Assoc. JMLA 2021, 109, 159. [Google Scholar] [CrossRef]
- Silverman, J.D.; Roche, K.; Mukherjee, S.; David, L.A. Naught all zeros in sequence count data are the same. Comput. Struct. Biotechnol. J. 2020, 18, 2789–2798. [Google Scholar] [CrossRef] [PubMed]
- Bailer, A.J. Combining, Extracting, and Reshaping Data. In Statistical Programming in SAS; Chapman and Hall/CRC: Boca Raton, FL, USA, 2020; pp. 137–190. [Google Scholar]
- Niglas, K. Media Review: Microsoft Office Excel Spreadsheet Software. J. Mix. Methods Res. 2007, 1, 297–299. [Google Scholar] [CrossRef]
- Mourad, M.; Jacob, T.; Sadovsky, E.; Bejerano, S.; Simone, G.S.-D.; Bagalkot, T.R.; Zucker, J.; Yin, M.T.; Chang, J.Y.; Liu, L. Placental response to maternal SARS-CoV-2 infection. Sci. Rep. 2021, 11, 14390. [Google Scholar] [CrossRef]
- Curtis, A.E.; Smith, T.A.; Ziganshin, B.A.; Elefteriades, J.A. The mystery of the Z-score. Aorta 2016, 4, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Clarkson-Townsend, D.A.; Kennedy, E.; Everson, T.M.; Deyssenroth, M.A.; Burt, A.A.; Hao, K.; Chen, J.; Pardue, M.T.; Marsit, C.J. Seasonally variant gene expression in full-term human placenta. FASEB J. 2020, 34, 10431–10442. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2012, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Pfannkuch, M. Year 11 students’ informal inferential reasoning: A case study about the interpretation of box plots. Int. Electron. J. Math. Educ. 2007, 2, 149–167. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.G. A picture is worth a thousand words: A cross-curricular approach to learning about visuals in STEM. Int. J. Eng. Edu. 2013, 29, 822–828. [Google Scholar]
- Nuzzo, A.; Carapezza, G.; Di Bella, S.; Pulvirenti, A.; Isacchi, A.; Bosotti, R. KAOS: A new automated computational method for the identification of overexpressed genes. BMC Bioinform. 2016, 17, 5–14. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Krzywinski, M.; Altman, N. Visualizing samples with box plots: Use box plots to illustrate the spread and differences of samples. Nat. Methods 2014, 11, 119–121. [Google Scholar] [CrossRef]
- Parsons, P.; Sedig, K. Common visualizations: Their cognitive utility. In Handbook of Human Centric Visualization; Springer: Berlin/Heidelberg, Germany, 2014; pp. 671–691. [Google Scholar]
- Simmons, S.S.; Isokpehi, R.D.; Brown, S.D.; McAllister, D.L.; Hall, C.C.; McDuffy, W.M.; Medley, T.L.; Udensi, U.K.; Rajnarayanan, R.V.; Ayensu, W.K. Functional annotation analytics of Rhodopseudomonas palustris genomes. Bioinform. Biol. Insights 2011, 5, 115–129. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Udensi, U.K.; Simmons, S.S.; Hollman, A.L.; Cain, A.E.; Olofinsae, S.A.; Hassan, O.A.; Kashim, Z.A.; Enejoh, O.A.; Fasesan, D.E. Evaluative profiling of arsenic sensing and regulatory systems in the human microbiome project genomes. Microbiol. Insights 2014, 7, MBI. S18076. [Google Scholar] [CrossRef]
- Fadiel, A.; Isokpehi, R.D.; Stambouli, N.; Hamza, A.; Benammar-Elgaaied, A.; Scalise, T.J. Protozoan parasite aquaporins. Expert Rev. Proteom. 2009, 6, 199–211. [Google Scholar] [CrossRef]
- Damiano, A.E. Aquaporins during pregnancy. In Vitamins and Hormones; Elsevier: Amsterdam, The Netherlands, 2020; Volume 112, pp. 327–355. [Google Scholar]
- Ma, N.; Liu, B.; Jin, Y.; Wang, J.; Qin, W.; Zheng, F.; Qin, R.; Li, J.; Hang, F.; Qin, A. AQP9 causes recurrent spontaneous abortion by inhibiting trophoblast cell EMT and invasion through the PI3K/AKT pathway. Biol. Reprod. 2023, 109, 736–748. [Google Scholar] [CrossRef]
- Sun, Y.V. Integration of biological networks and pathways with genetic association studies. Hum. Genet. 2012, 131, 1677–1686. [Google Scholar] [CrossRef][Green Version]
- Pillich, R.T.; Chen, J.; Churas, C.; Fong, D.; Gyori, B.M.; Ideker, T.; Karis, K.; Liu, S.N.; Ono, K.; Pico, A. NDEx IQuery: A multi-method network gene set analysis leveraging the Network Data Exchange. Bioinformatics 2023, 39, btad118. [Google Scholar] [CrossRef]
- Seal, R.L.; Braschi, B.; Gray, K.; Jones, T.E.; Tweedie, S.; Haim-Vilmovsky, L.; Bruford, E.A. Genenames.org: The HGNC resources in 2023. Nucleic Acids Res. 2023, 51, D1003–D1009. [Google Scholar] [CrossRef]
- Darden, D.B.; Stortz, J.A.; Hollen, M.K.; Cox, M.C.; Apple, C.G.; Hawkins, R.B.; Rincon, J.C.; Lopez, M.-C.; Wang, Z.; Navarro, E.; et al. Identification of unique mRNA and miRNA expression patterns in bone marrow hematopoietic stem and progenitor cells after trauma in older adults. Front. Immunol. 2020, 11, 1289. [Google Scholar] [CrossRef]
- Matsuda, S.; Kobayashi, M.; Kitagishi, Y. Expression and function of PPARs in placenta. PPAR Res. 2013, 2013, 256508. [Google Scholar] [CrossRef]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, R.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef]
- Herrera, C.L.; Kim, M.J.; Do, Q.N.; Owen, D.M.; Fei, B.; Madhuranthakam, A.J.; Xi, Y.; Lewis, M.A.; Twickler, D.M.; Spong, C.Y. The Human Placenta Project: Funded projects, imaging innovation, and persistent gaps. Am. J. Obstet. Gynecol. 2022, 226, S233–S234. [Google Scholar] [CrossRef]
- Herrera, C.L.; Kim, M.J.; Do, Q.N.; Owen, D.M.; Fei, B.; Twickler, D.M.; Spong, C.Y. The human placenta project: Funded studies, imaging technologies, and future directions. Placenta 2023, 142, 27–35. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Wang, Y. The Placenta: A Maternofetal Interface. Annu. Rev. Nutr. 2023, 43, 301–325. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.E.; Chan, S.-Y. Current approaches and developments in transcript profiling of the human placenta. Hum. Reprod. Update 2020, 26, 799–840. [Google Scholar] [CrossRef]
- McDermaid, A.; Monier, B.; Zhao, J.; Liu, B.; Ma, Q. Interpretation of differential gene expression results of RNA-seq data: Review and integration. Brief. Bioinform. 2019, 20, 2044–2054. [Google Scholar] [CrossRef]
- Buchrieser, J.; Schwartz, O. Pregnancy complications and Interferon-induced transmembrane proteins (IFITM): Balancing antiviral immunity and placental development. Comptes Rendus Biol. 2021, 344, 145–156. [Google Scholar] [CrossRef]
- Buchrieser, J.; Degrelle, S.A.; Couderc, T.; Nevers, Q.; Disson, O.; Manet, C.; Donahue, D.A.; Porrot, F.; Hillion, K.-H.; Perthame, E. IFITM proteins inhibit placental syncytiotrophoblast formation and promote fetal demise. Science 2019, 365, 176–180. [Google Scholar] [CrossRef]
- Zani, A.; Zhang, L.; McMichael, T.M.; Kenney, A.D.; Chemudupati, M.; Kwiek, J.J.; Liu, S.-L.; Yount, J.S. Interferon-induced transmembrane proteins inhibit cell fusion mediated by trophoblast syncytins. J. Biol. Chem. 2019, 294, 19844–19851. [Google Scholar] [CrossRef]
- Robin, C.; Bollerot, K.; Mendes, S.; Haak, E.; Crisan, M.; Cerisoli, F.; Lauw, I.; Kaimakis, P.; Jorna, R.; Vermeulen, M. Human placenta is a potent hematopoietic niche containing hematopoietic stem and progenitor cells throughout development. Cell Stem Cell 2009, 5, 385–395. [Google Scholar] [CrossRef]
- Fraser, S.T. The modern primitives: Applying new technological approaches to explore the biology of the earliest red blood cells. Int. Sch. Res. Not. 2013, 568928. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Paltoo, A.; Kelly, R.; Sallenave, J.-M.; Bocking, A.; Challis, J. Expression of natural antimicrobials by human placenta and fetal membranes. Placenta 2007, 28, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Beach, D.; Su, L.; Zhai, R.; Christiani, D.C. A genome-wide expression analysis in blood identifies pre-elafin as a biomarker in ARDS. Am. J. Respir. Cell Mol. Biol. 2008, 38, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Kothiyal, P.; Schulkers, K.; Liu, X.; Hazrati, S.; Vilboux, T.; Gomez, L.M.; Huddleston, K.; Wong, W.S.; Niederhuber, J.E.; Conrads, T.P. Differences in maternal gene expression in Cesarean section delivery compared with vaginal delivery. Sci. Rep. 2020, 10, 17797. [Google Scholar] [CrossRef]
- Brundage, J.F. Interactions between influenza and bacterial respiratory pathogens: Implications for pandemic preparedness. Lancet Infect. Dis. 2006, 6, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Sharps, M.C.; Hayes, D.J.; Lee, S.; Zou, Z.; Brady, C.A.; Almoghrabi, Y.; Kerby, A.; Tamber, K.K.; Jones, C.J.; Waldorf, K.M.A. A structured review of placental morphology and histopathological lesions associated with SARS-CoV-2 infection. Placenta 2020, 101, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Tuuli, M.; Longtine, M.; Nelson, D. Oxygen and trophoblast biology–a source of controversy. Placenta 2011, 32, S109–S118. [Google Scholar] [CrossRef] [PubMed]
- Ölmez, F.; Oğlak, S.C.; Can, E. The implication of aquaporin-9 in the pathogenesis of preterm premature rupture of membranes. Z. Geburtshilfe Neonatol. 2022, 226, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Ölmez, F.; Oğlak, S.C.; Gedik Özköse, Z. Increased maternal serum aquaporin-9 expression in pregnancies complicated with early-onset preeclampsia. J. Obstet. Gynaecol. Res. 2022, 48, 647–653. [Google Scholar] [CrossRef]
- Prins, J.R.; Eskandar, S.; Eggen, B.J.; Scherjon, S.A. Microglia, the missing link in maternal immune activation and fetal neurodevelopment; and a possible link in preeclampsia and disturbed neurodevelopment? J. Reprod. Immunol. 2018, 126, 18–22. [Google Scholar] [CrossRef]
- Xu, P.; Zhang, X.; Liu, Q.; Xie, Y.; Shi, X.; Chen, J.; Li, Y.; Guo, H.; Sun, R.; Hong, Y. Microglial TREM-1 receptor mediates neuroinflammatory injury via interaction with SYK in experimental ischemic stroke. Cell Death Dis. 2019, 10, 555. [Google Scholar] [CrossRef]
- Linzer, D.I.; Fisher, S.J. The placenta and the prolactin family of hormones: Regulation of the physiology of pregnancy. Mol. Endocrinol. 1999, 13, 837–840. [Google Scholar] [CrossRef]
- Napso, T.; Yong, H.E.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The role of placental hormones in mediating maternal adaptations to support pregnancy and lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef]
- He, M.; Liu, P.; Lawrence-Dill, C.J. A hypothesis-driven approach to assessing significance of differences in RNA expression levels among specific groups of genes. Curr. Plant Biol. 2017, 11, 46–51. [Google Scholar] [CrossRef]
- Suh, A.; Jiang, Y.; Mosca, A.; Wu, E.; Chang, R. A Grammar for Hypothesis-Driven Visual Analysis. arXiv 2022, arXiv:2204.14267. [Google Scholar]
- Guo, W.; Tzioutziou, N.A.; Stephen, G.; Milne, I.; Calixto, C.P.; Waugh, R.; Brown, J.W.; Zhang, R. 3D RNA-seq: A powerful and flexible tool for rapid and accurate differential expression and alternative splicing analysis of RNA-seq data for biologists. RNA Biol. 2021, 18, 1574–1587. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, D.H. Human Placenta Project. Available online: https://www.nichd.nih.gov/research/supported/human-placenta-project/default (accessed on 25 July 2023).
- Clark, A.R.; Chernyavsky, I.L.; Jensen, O.E. The complexities of the human placenta. Phys. Today 2023, 76, 26–32. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aquaporin | Binary Number Pattern of Gene Expression | Genes with Binary Number Pattern | Example Genes with Binary Number Pattern * |
|---|---|---|---|
| AQP0 (MIP) | 010010000010000000 | 656 | ESX1 TLX1 |
| AQP1 | 000011111011111011 | 45 | BRCA1 CD9 |
| AQP2 | 010011010010000010 | 95 | TSLP VIP |
| AQP3 | 000011011010011111 | 17 | HOXD8 TPRV2 |
| AQP4 | 000010010010000000 | 54 | CD1C NOX1 |
| AQP5 | 000010010000010000 | 15 | HEPHL1 KLF4 |
| AQP6, AQP7 | 000011010010010000 | 588 | CFTR HNF4A |
| AQP8 | 010011010010000000 | 352 | CALHM1 FGF21 |
| AQP9 | 000000101000000000 | 4 | PI3 TREM1 |
| AQP10 | 010011000010010000 | 212 | OR4C16 TLR9 |
| AQP11 | 000011011011000000 | 25 | IL4I1 SLC6A4 |
| AQP12A | 010010000000000010 | 36 | MDFIC2 PRL |
| AQP12B | 010011010010010010 | 473 | CETP CACNA1I |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isokpehi, R.D.; Abioye, A.O.; Hamilton, R.S.; Fryer, J.C.; Hollman, A.L.; Destefano, A.M.; Ezekiel, K.B.; Taylor, T.L.; Brooks, S.F.; Johnson, M.O.; et al. Visual Analytics for Robust Investigations of Placental Aquaporin Gene Expression in Response to Maternal SARS-CoV-2 Infection. Analytics 2024, 3, 116-139. https://doi.org/10.3390/analytics3010007
Isokpehi RD, Abioye AO, Hamilton RS, Fryer JC, Hollman AL, Destefano AM, Ezekiel KB, Taylor TL, Brooks SF, Johnson MO, et al. Visual Analytics for Robust Investigations of Placental Aquaporin Gene Expression in Response to Maternal SARS-CoV-2 Infection. Analytics. 2024; 3(1):116-139. https://doi.org/10.3390/analytics3010007
Chicago/Turabian StyleIsokpehi, Raphael D., Amos O. Abioye, Rickeisha S. Hamilton, Jasmin C. Fryer, Antoinesha L. Hollman, Antoinette M. Destefano, Kehinde B. Ezekiel, Tyrese L. Taylor, Shawna F. Brooks, Matilda O. Johnson, and et al. 2024. "Visual Analytics for Robust Investigations of Placental Aquaporin Gene Expression in Response to Maternal SARS-CoV-2 Infection" Analytics 3, no. 1: 116-139. https://doi.org/10.3390/analytics3010007
APA StyleIsokpehi, R. D., Abioye, A. O., Hamilton, R. S., Fryer, J. C., Hollman, A. L., Destefano, A. M., Ezekiel, K. B., Taylor, T. L., Brooks, S. F., Johnson, M. O., Smile, O., Ramroop-Butts, S., Makolo, A. U., & Hayward, A. G., II. (2024). Visual Analytics for Robust Investigations of Placental Aquaporin Gene Expression in Response to Maternal SARS-CoV-2 Infection. Analytics, 3(1), 116-139. https://doi.org/10.3390/analytics3010007