
Caletodraco cottardi: A New Furileusaurian Abelisaurid (Dinosauria: Theropoda) from the Cenomanian Chalk of Normandy (North-Western France) †




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
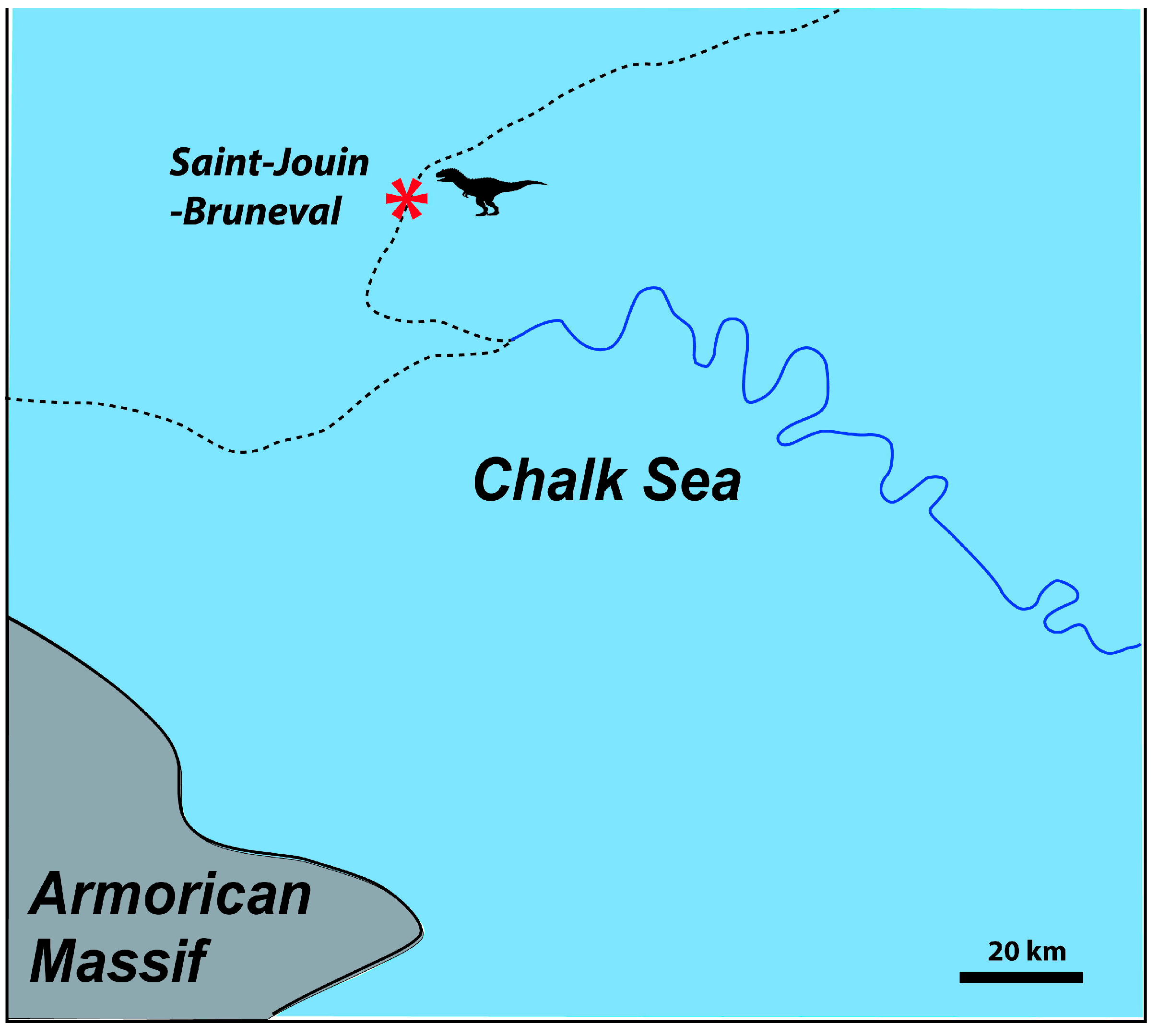
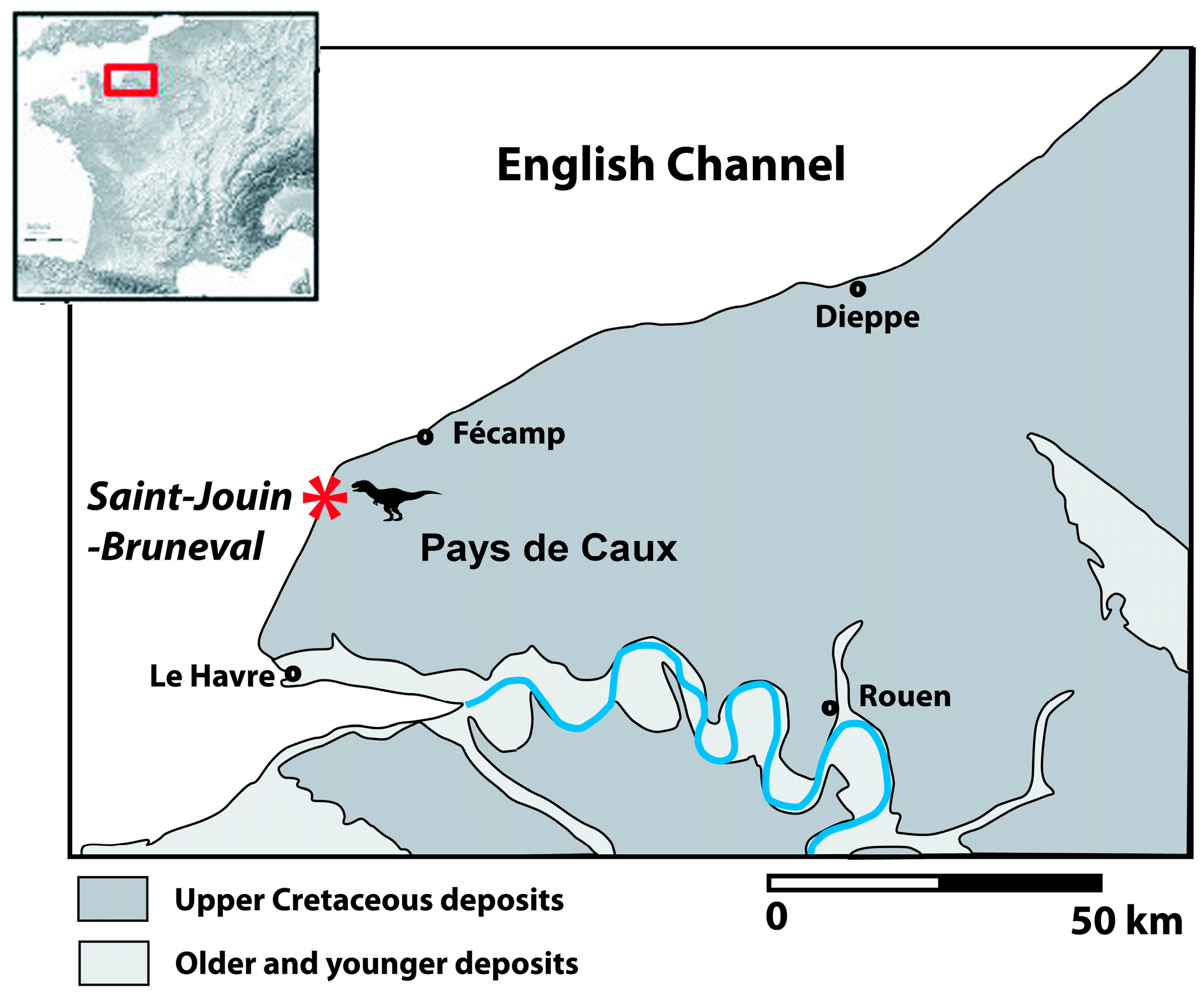
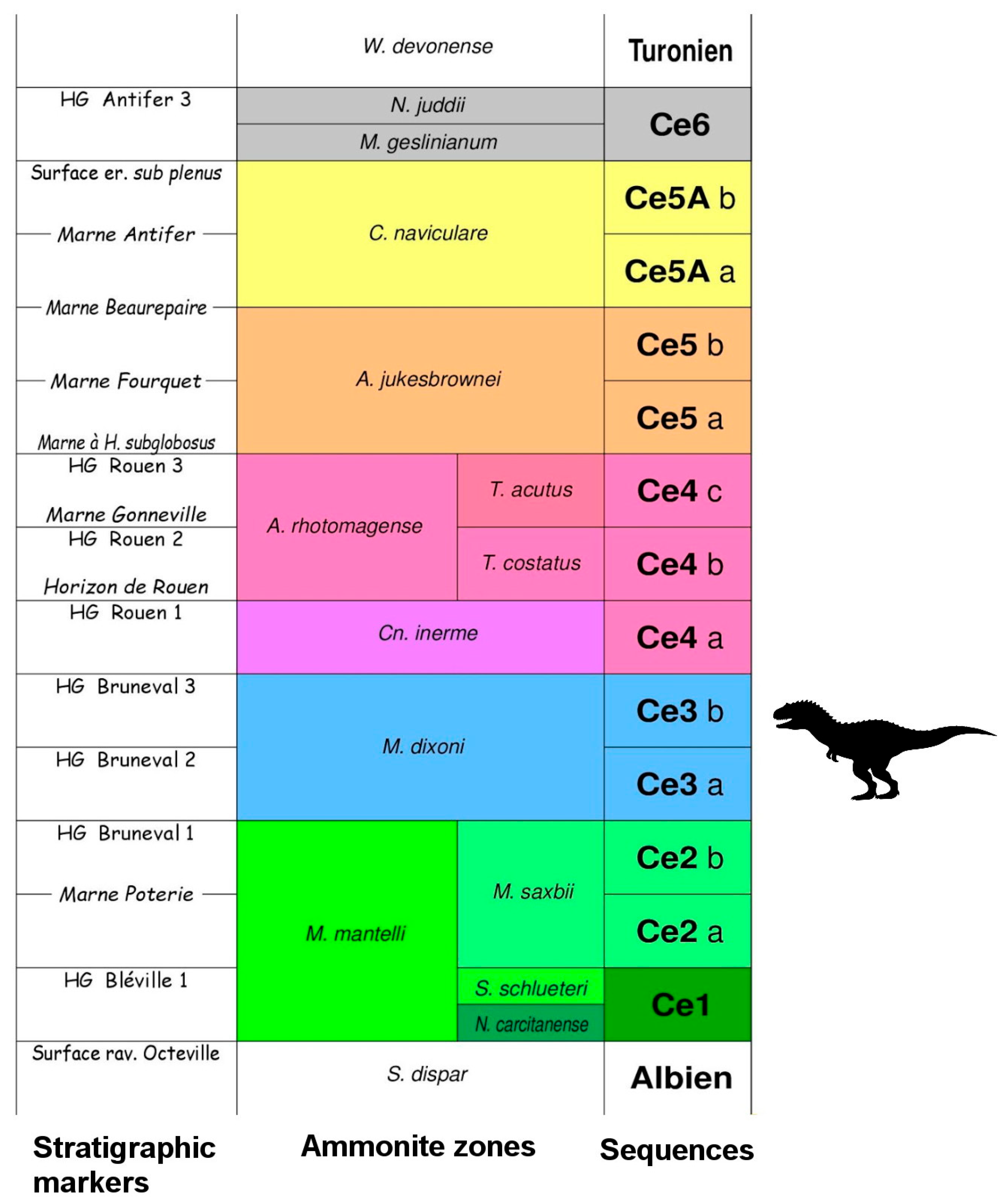
2. Geographical and Geological Settings

3. Material and Methods
4. Systematic Paleontology

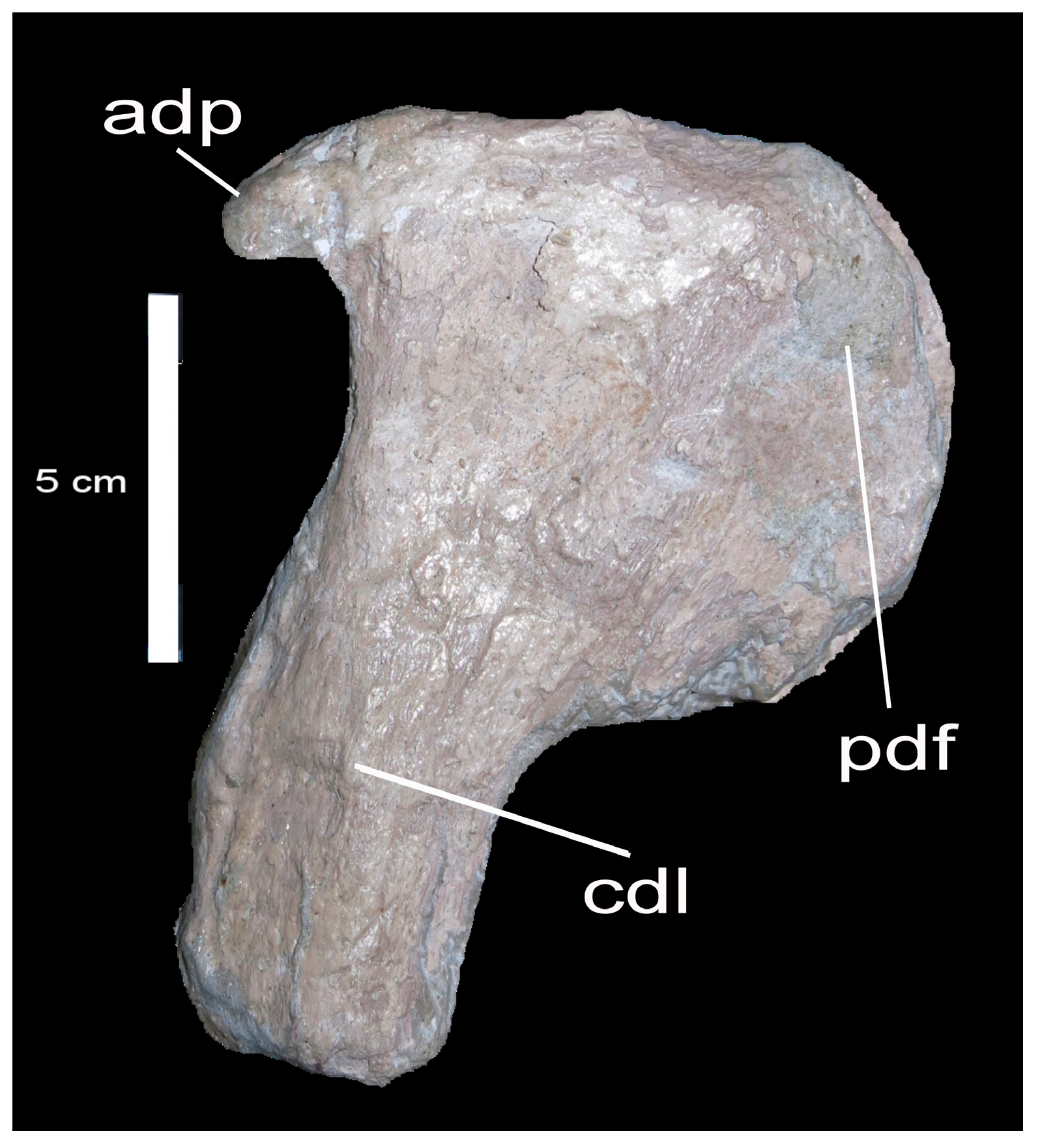
4.1. Description


Caudal Vertebra
4.2. Measurements and Size Estimate
4.3. Comparisons and Identification
- −
- Presence of six strongly fused sacral vertebrae. The number of sacral vertebrae is variable in abelisauroids [34]. The furileusaurians Carnotaurus and Aucasaurus have six, the majungasaurine Majungasaurus only five, but the noasaurid Masiakasaurus has six. The degree of fusion of the sacral vertebrae is also variable. In Majungasaurus [42], sutures are still visible between centra and neural arches. The advanced degree of fusion seen in Caletodraco cottardi can probably be considered as a characteristic of derived abelisaurids.
- −
- Sacrum constricted in its middle part. This character, resulting from the narrowness of the mid-sacral vertebrae, is present in many ceratosaurs [34] and well developed in furileusaurians such as Carnotaurus and Aucasaurus, but is apparently absent in Majungasaurus.
- −
- −
- Well-developed pneumaticity of the neural spines and transverse processes of the sacral vertebrae. This is a ceratosaurian character according to Carrano and Sampson [44].
- −
- Wide base of the neural arch of the first caudal vertebra. This character is widespread among abelisauroids according to Carrano and Sampson [44].
- −
- Presence of a hyposphene on the neural arch of the first caudal vertebra. A hyposphene–hypantrum accessory articulation is present in Carnotaurus, Aucasaurus, Ekrixinatosaurus, Skorpiovenator, Pycnonemosaurus [45], Kurupi [46] and Koleken [3] but absent in Ilokelesia, Majungasaurus [45] and Arcovenator [12].
- −
- −
- Distally expanded transverse processes of the first caudal vertebra, with a convex distal margin and a pointed anterior process. A great distal expansion of the transverse processes of anterior and mid-caudal vertebrae was considered as a synapomorphy of Brachyrostra by Canale et al. [35]. Filippi et al. [16] considered a convex distal edge of the transverse process, and a strong development of an anterior projection as a synapomorphy of Furileusauria. See below for a detailed discussion of this feature, which appears to be a distinguishing characteristic of furileusaurians.
- −
- Dorsal margin of the ilium straight (Figure 11). This character is found in derived abelisaurids such as the furileusaurian Brachyrostra Carnotaurus sastrei [48], Aucasaurus garridoi [34] and Koleken inakayali [3]. In the less derived Majungasaurus crenatissimus [42] and Skorpiovenator bustingorryi [49], the dorsal margin of the ilium is convex, while it is possibly undulated in Rahiolisaurus gujaratensis [50] and Ekrixinatosaurus novasi [51]. Interestingly, a straight dorsal margin is also present in Genusaurus sisteronis, from the Albian of France, a possible furileusaurian (see below and [34]). It should be noted, however, that this character is also known in the noasaurid Masiakasaurus knopfleri [52].
- −
- Posterior margin of the ilium with a deep notch and a posterodorsal process (Figure 11). This is a widespread character in Abelisauridae, being found in both majungasaurines (Majungasaurus) and brachyrostrans (Aucasaurus, Carnotaurus).
- −
- Brevis fossa becoming significantly wider posteriorly. This feature appears to be widespread in abelisaurids, and more generally in ceratosaurs [43].

5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonaparte, J.F.; Novas, F.E. Abelisaurus comahuensis, n.g., n. sp., Carnosauria del Cretácico tardio de Patagonia. Ameghiniana 1985, 21, 259–265. [Google Scholar]
- Delcourt, R. Ceratosaur palaeobiology: New insights on evolution and ecology of the southern rulers. Sci. Rep. 2018, 8, 9730. [Google Scholar] [CrossRef] [PubMed]
- Pol, D.; Baiano, M.A.; Černý, D.; Novas, F.E.; Cerda, I.A.; Pittman, M. A new abelisaurid dinosaur from the end Cretaceous of Patagonia and evolutionary rates among the Ceratosauria. Cladistics 2024, 40, 307–356. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.L.; Candeiro, C.A.S.; da Silva Vidal, L.; Brusatte, S.L.; Mortimer, M. Abelisauroidea (Theropoda, Dinosauria) from Africa: A review of the fossil record. Pap. Av. Zool. 2023, 63, e202363019. [Google Scholar] [CrossRef]
- Prasad, G.V.R.; Verma, V.; Grover, P.; Priyadarshini, R.; Sahni, A.; Lourembam, R.S. Isolated archosaur teeth from the green sandstone capping the Coralline Limeston (Bagh Groupp of the Narmada valley: Evidence for the presence of pre-Late to Late Maastrichtian abelisaurids in India. Island Arc 2016, 25, 410–420. [Google Scholar] [CrossRef]
- Krause, D.W.; Sampson, S.D.; Carrano, M.T.; O’Connor, P.M. Overview of the history, taxonomy, phylogeny, and biogeography of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Soc. Vert. Paleont. Mem. 2007, 8, 1–20. [Google Scholar]
- Buffetaut, E.; Mechin, P.; Mechin-Salessy, A. Un dinosaure théropode d’affinités gondwaniennes dans le Crétacé supérieur de Provence. C. R. Acad. Sc. Paris IIA 1988, 206, 153–158. [Google Scholar]
- Buffetaut, E. The discontinuous fossil record of European Abelisauridae and its significance for abelisaurid evolution and palaeobiogeography. Evoluçao 2024, 3, 19–21. [Google Scholar]
- Le Loeuff, J.; Buffetaut, E. Tarascosaurus salluvicus nov. gen., nov. sp., dinosaure théropode du Crétacé supérieur du Sud de la France. Geobios 1991, 25, 585–594. [Google Scholar] [CrossRef]
- Buffetaut, E.; Le Loeuff, J. Commentaire à la note Découverte d’un dinosaure théropode nouveau (Genusaurus sisteronis n.g., n.sp.) dans l’Albien marin de Sisteron (Alpes de Haute-Provence, France) et extension au Crétacé inférieur de la lignée cératosaurienne. C. R. Acad. Sc. Paris IIa 1995, 320, 79–80. [Google Scholar]
- Buffetaut, E. Sur les Chemins des Dinosaures; Aurian: Béziers, France, 2005. [Google Scholar]
- Tortosa, T.; Buffetaut, E.; Vialle, N.; Dutour, Y.; Turini, E.; Cheylan, G. A new abelisaurid dinosaur from the Late Cretaceous of southern France: Palaeobiogeographical implications. Ann. Paléont. 2014, 100, 63–86. [Google Scholar] [CrossRef]
- Pérez-García, A.; Bardet, N.; Fregenal-Martinez, M.A.; Martin-Jiménez, M.; Mocho, P.; Narváez, I.; Torices, A.; Vullo, R.; Ortega, F. Cenomanian vertebrates from Algora (central Spain): New data on the establishment of the European Upper Cretaceous continental fauna. Cret. Res. 2020, 115, 104566. [Google Scholar] [CrossRef]
- Isasmendi, E.; Torices, A.; Canudo, J.I.; Currie, P.J.; Pereda-Suberbiola, X. Upper Cretaceous European theropod palaeobiodiversity, palaeobiogeography and the intra-Maastrichtian faunal turnover: New contributions from the Iberian fossil site of Laño. Papers Palaeont. 2022, 8, e1419. [Google Scholar] [CrossRef]
- Ösi, A.; Buffetaut, E. Additional non-avian theropod and bird remains from the early Late Cretaceous (Santonian) of Hungary and a review of the European abelisauroid record. Ann. Paléont. 2011, 97, 35–49. [Google Scholar] [CrossRef]
- Filippi, L.S.; Méndez, A.H.; Juárez Valieri, R.D.; Garrido, A.C. A new brachyrostran with hypertrophied axial structures reveals an unexpected radiation of latest Cretaceous abelisaurids. Cret. Res. 2016, 61, 209–219. [Google Scholar] [CrossRef]
- Hoyez, B.; Girard, J.; Cottard, N. Le Cénomanien du littoral normand entre la valleuse d’Antifer et le cap de la Hève; Carnets Géol.: Madrid, Spain, 2020; 225p, ISBN 13: 978-2-916733-19-7, Book 2020/01 (CG2020_B01). [Google Scholar]
- Juignet, P. Transgressions-régressions, variations eustatiques et influences tectoniques de l’Aptien au Maastrichtien dans le Bassin de Paris occidental et sur la bordure du Massif Armoricain. Cret. Res. 1980, 1, 341–357. [Google Scholar] [CrossRef]
- Juignet, P.; Kennedy, W.J. Faunes d’ammonites et biostratigraphie comparée du Cénomanien du nord-ouest de la France (Normandie) et du sud de l’Angleterre. Bull. Trim. Soc. géol. Normandie Amis Mus. Havre 1976, 63, 5–193. [Google Scholar]
- Lasseur, E.; Guillocheau, F.; Robin, C.; Hanot, F.; Vaslet, D.; Coueffe, R.; Néraudeau, D. A relative water-depth model for the Normandy Chalk (Cenomanian–Middle Coniacian, Paris Basin, France) based on facies patterns of metre-scale cycles. Sediment. Geol. 2009, 213, 1–26. [Google Scholar] [CrossRef]
- Buffetaut, E. The significance of dinosaur remains in marine sediments: An investigation based on the French record. Berlin. Geowiss. Abh. E 1994, 13, 125–133. [Google Scholar]
- Breton, G. Excursions géologiques sur le littoral entre le Havre et Etretat (Normandie, France). Bull. Trim. Soc. Géol. Normandie Amis Mus. Havre 1981, 68, 1–56. [Google Scholar]
- Lennier, G. Etudes Géologiques et Paléontologiques sur l’Embouchure de la Seine et les Falaises de la Haute-Normandie; Imprimerie Eugène Costey: Le Havre, France, 1868; 245p. [Google Scholar]
- Lennier, G. Muséum du Havre. Petit Guide Illustré du Visiteur; Imprimerie du Journal Le Havre: Le Havre, France, 1904; 371p. [Google Scholar]
- Accarie, H.; Beaudoin, B.; Dejax, J.; Friès, G.; Michard, J.G.; Taquet, P. Découverte d’un dinosaure théropode nouveau (Genusaurus sisteronis n.g., n.sp.) dans l’Albien marin de Sisteron (Alpes de Haute-Provence, France) et extension au Crétacé inférieur de la lignée cératosaurienne. C. R. Acad. Sc. Paris IIa 1995, 320, 327–334. [Google Scholar]
- Buffetaut, E.; Escuillié, F.; Pohl, B. First theropod dinosaur from the Maastrichtian phosphates of Morocco. Kaupia 2005, 14, 3–8. [Google Scholar]
- Longrich, N.R.; Pereda-Sberbiola, X.; Jalil, N.; Khaldoune, F.; Jourani, E. An abelisaurid from the latest Cretaceous (late Maastrichtian) of Morocco, North Africa. Cret. Res. 2017, 76, 40–52. [Google Scholar] [CrossRef]
- Longrich, N.R.; Isasmendi, E.; Pereda-Suberbiola, X.; Jalil, N.E. New fossils of Abelisauridae (Dinosauria: Theropoda) from the upper Maastrichtian of Morocco, North Africa. Cret. Res. 2023, 152, 105677. [Google Scholar] [CrossRef]
- Gemmellaro, M. Rettili maëstrichiani di Egitto. Giorn. Sci. Natur. Econ. 1921, 32, 339–351. [Google Scholar]
- Smith, J.B.; Lamanna, M. An abelisaurid from the Late Cretaceous of Egypt: Implications for theropod biogeography. Naturwissenschaften 2006, 93, 242–245. [Google Scholar] [CrossRef]
- Milner, A.C. Reptiles. In Fossils of the Chalk, 2nd ed.; Smith, A.B., Batten, D.J., Eds.; The Palaeontological Association: London, UK, 2002; pp. 325–343. [Google Scholar]
- Streiff, R. La Normandie avant les Normands. Epoque gallo-romaine. Etudes Normandes 1952, 4, 133–144. [Google Scholar] [CrossRef]
- Rogers, R.R.; Krause, D.W.; Curry Rogers, K. Cannibalism in the Madagascan dinosaur Majungatholus atopus. Nature 2003, 422, 515–518. [Google Scholar] [CrossRef]
- Baiano, M.A.; Coria, R.; Chiappe, L.M.; Zurriaguz, V.; Coria, L. Osteology of the axial skeleton of Aucasaurus garridoi: Phylogenetic and paleobiological inferences. PeerJ 2023, 11, e16236. [Google Scholar] [CrossRef]
- Canale, J.I.; Scanferla, C.A.; Agnolin, F.I.; Novas, F.E. New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods. Naturwissenschaften 2009, 96, 409–414. [Google Scholar] [CrossRef]
- Grillo, O.N.; Delcourt, R. Allometry and body length of abelisauroid theropods: Pycnocnemosaurus nevesi is the new king. Cret. Res. 2017, 69, 71–89. [Google Scholar] [CrossRef]
- Hendrickx, C.; Tschopp, E.; Ezcurra, M.D. Taxonomic identification of isolated theropod teeth: The case of the shed tooth crown associated with Aerosteon (Theropoda: Megarptora) and the dentition of Abelisauridae. Cret. Res. 2020, 108, 104312. [Google Scholar] [CrossRef]
- Meso, J.G.; Hendrickx, C.; Baiano, M.A.; Canale, J.I.; Salgado, L.; Díaz-Martínez, I. Isolated theropod teeth associated with a sauropod skeleton from the Late Cretaceous Allen Formation of Río Negro, Patagonia, Argentina. Acta Palaeont. Polon. 2021, 66, 409–423. [Google Scholar]
- Ribeiro, T.B.; Gomes da Costa Pereiro, P.V.L.; Brusatte, S.L.; Candeiro, C.R.A.; Bergqvist, L.P. An eye for an eye, a tooth for a tooth: Archosaurian teeth from the Açu Formation (Albian-Cenomanian), Potiguar Basin, Northeast Brazil. Cret. Res. 2022, 129, 105005. [Google Scholar] [CrossRef]
- Giaretta, A.A.; Coelho Rodrigues, S.; Buck, P.V. A new geographical record of Abelisauridae (Theropoda, Dinosauria) for the Bauru Group (Upper Cretaceous). J. S. Am. Earth Sc. 2024, 133, 104722. [Google Scholar] [CrossRef]
- Delcourt, R.; Brilhante, N.S.; Pires-Domingues, R.A.; Hendrickx, C.; Grillo, O.N.; Augusta, B.G.; Maciel, B.S.; Ghilardi, A.; Ricardi-Branco, F. Biogeography of theropod dinosaurs during the Late Cretaceous: Evidence from central South America. Zool. J. Linn. Soc. 2024, 20, 1–40. [Google Scholar] [CrossRef]
- O’Connor, P.M. The postcranial axial skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Soc. Vert. Paleont. Mem. 2007, 8, 127–162. [Google Scholar] [CrossRef]
- Cerroni, M.A.; Motta, M.J.; Agnolin, F.L.; Aranciaga Rolando, A.M.; Brissón Egli, F.; Novas, F.E. A new abelisaurid from the Huincul Formation (Cenomanian-Turonian; Upper Cretaceous) of Rio Negro province, Argentina. J. S. Am. Earth Sc. 2020, 98, 102445. [Google Scholar] [CrossRef]
- Carrano, M.T.; Sampson, S.D. The phylogeny of Ceratosauria (Dinosauria: Theropoda). J. System. Palaeont. 2008, 6, 183–236. [Google Scholar] [CrossRef]
- Méndez, A.H. The caudal vertebral series in abelisaurid dinosaurs. Acta Palaeont. Polon. 2014, 59, 99–107. [Google Scholar]
- Iori, F.V.; de Araújo-Júnior, H.I.; Simionato Tavares, S.A.; da Silva Marinho, T.; Martinelli, A.G. New theropod dinosaur from the Late Cretaceous. J. S. Am. Earth Sc. 2021, 112, 103551. [Google Scholar] [CrossRef]
- Ezcurra, M.D.; Méndez, A.H. First report of a derived abelisaurid theropod from the Bajo de la Carpa Formation (Late Cretaceous), Patagonia, Argentina. Bull. Geosc. 2009, 84, 547–554. [Google Scholar] [CrossRef]
- Bonaparte, J.F.; Novas, F.E.; Coria, R.A. Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the middle Cretaceous of Patagonia. Contrib. Sc. Nat. Hist. Mus. Los Angeles Cty. 1990, 416, 1–42. [Google Scholar] [CrossRef]
- Cerroni, M.; Baiano, M.A.; Canale, J.I.; Agnolin, F. Appendicular osteology of Skorpiovenator bustingorryi (Theropoda, Abelisauridae) with comments on phylogenetic features of abelisaurids. J. Syst. Palaeont. 2022, 20, 2093661. [Google Scholar] [CrossRef]
- Novas, F.E.; Chatterjee, S.; Rudra, D.K.; Datta, P.M. Rahiolisaurus gujaratensis, n.g. n.sp., a new abelisaurid theropod from the Late Cretaceous of India. In New aspects of Mesozoic Biodiversity; Bandyopadhyay, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 45–62. [Google Scholar]
- Calvo, J.O.; Rubilar-Rogers, D.; Moreno, K. A new Abelisauridae (Dinosauria: Theropoda) from northwest Patagonia. Ameghiniana 2004, 41, 555–563. [Google Scholar]
- Carrano, M.T.; Loewen, M.A.; Sertich, J.J.W. New materials of Masiakasaurus knofleri Sampson, Carrano, and Forster, 2001, and implications for the morphology of the Noasauridae (Theropoda: Ceratosauria). Smithson. Contrib. Paleobiol. 2011, 95, 1–53. [Google Scholar] [CrossRef]
- Filippi, L.S.; Méndez, A.H.; Gianechini, F.A.; Juárez Valieri, R.D.; Garrido, A.C. Osteology of Viavenator exxoni (Abelisauridae; Furileusauria) from the Bajo de la Carpa Formation, NW Patagonia, Argentina. Cret. Res. 2018, 83, 95–119. [Google Scholar] [CrossRef]
- Gianechini, F.A.; Méndez, A.; Filippi, L.; Garrido, A. A non-furileusurian caudal vertebra from the Bajo de la Carpa Formation (Upper Cretaceous, Santonian) and morphological variation in the tail of Abelisauridae. Pub. Electr. As. Paleont. Argent. 2022, 22, 58–70. [Google Scholar]
- Coria, R.A.; Salgado, L. A basal Abelisauria Novas, 1992 (Theropoda- Ceratosauria) from the Cretaceous of Patagonia, Argentina. Gaia 1998, 15, 89–102. [Google Scholar]
- Delcourt, R. Revised morphology of Pycnocnemosaurus nevesi Kellner & Campos, 2002 (Theropoda, Abelisauridae) and its phylogenetic relationships. Zootaxa 2017, 4276, 1–45. [Google Scholar]
- Coria, R.A.; Chiappe, L.M.; Dingus, L. A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda, Abelisauridae) from the Late Cretaceous of Patagonia. J. Vert. Paleont. 2002, 22, 460–465. [Google Scholar] [CrossRef]
- Carrano, M.T. The appendicular skeleton of of Majungasaurus crenatissimus (Theropode: Abelisauridae) from the Late Cretaceous of Madagascar. Soc. Vert. Paleont. Mem. 2007, 8, 163–179. [Google Scholar] [CrossRef]
- Zitouni, S.; Laurent, C.; Dyke, G.; Jalil, N. An abelisaurid (Dinosauria: Theropode) ilium from the Upper Cretaceous (Cenomanian) of the Kem Kem beds, Morocco. PLoS ONE 2019, 14, e0214055. [Google Scholar] [CrossRef] [PubMed]
- Novas, F.E.; Agnolin, F.L.; Bandyopadhyay, S. Cretaceous theropods from India: A review of specimens described by Huene and Matley (1933). Rev. Mus. Argent. Cienc. Nat. 2004, 6, 67–103. [Google Scholar] [CrossRef]
- Baiano, M.A.; Pol, D.; Bellardini, F.; Windholz, G.J.; Cerda, I.A.; Garrido, A.C.; Coria, R.A. Elemgasem nubilus: A new brachyrostran abelisaurid (Theropoda, Ceratosauria) from the Portezuelo Formation (Upper Cretaceous) of Patagonia, Argentina. Papers Palaeont. 2022, 8, e1462. [Google Scholar] [CrossRef]
- Accarie, H.; Beaudoin, B.; Dejax, J.; Friès, G.; Michard, J.G.; Taquet, P. Réponse au commentaire de Buffetaut et Le Loeuff sur la note C. R. Acad. Sci. Paris 1995: Découverte d’un dinosaure théropode nouveau (Genusaurus sisteronis n.g., n.sp.) dans l’Albien marin de Sisteron (Alpes de Haute-Provence, France) et extension au Crétacé inférieur de la lignée cératosaurienne. C. R. Acad. Sc. Paris IIa 1995, 321, 81–83. [Google Scholar]
- Agnolin, F.L.; Cerroni, M.A.; Scanferla, A.; Goswami, A.; Paulina-Carabajal, A.; Halliday, T.; Cuff, A.R.; Reuil, S. First definitive abelisaurid theropod from the Late Cretaceous of northwestern Argentina. J. Vert. Paleont. 2021, 41, e2002348. [Google Scholar] [CrossRef]
- Ezcurra, M.D.; Agnolin, F.L. An abelisauroid dinosaur from the Middle Jurassic of Laurasia and its implications on theropod palaeobiogeography and evolution. Proc. Geol. Ass. 2012, 123, 500–507. [Google Scholar] [CrossRef]
- Rauhut, O.W.M. A reappraisal of a putative record of abelisauroid theropod dinosaur from the Middle Jurassic of England. Proc. Geol. Ass. 2012, 123, 779–786. [Google Scholar] [CrossRef]
- Hendrickx, C.; Mateus, O. Abelisauridae (Dinosauria: Theropoda) from the Late Jurassic of Portugal and dentition-based phylogeny as a contribution for the identification of isolated theropod teeth. Zootaxa 2014, 3759, 1–74. [Google Scholar] [CrossRef]
- Malafaia, E.; Escaso, F.; Mocha, P.; Serrano-Martínez, A.; Torices, A.; Cachão, M.; Ortega, F. Analysis of diversity, stratigraphic and geographical distribution of isolated theropod teeth from the Upper Jurassic of the Lusitanian Basin, Portugal. J. Iber. Geol. 2017, 43, 257–291. [Google Scholar] [CrossRef]
- Gianechini, F.A.; Méndez, A.H.; Filippi, L.S.; Paulina-Carabajal, A.; Juárez Valieri, R.; Garrido, A.C. A new furileusaurian abelisaurid from La Invernada (Upper Cretaceous, Santonian, Bajo de la Carpa Formation), northern Patagonia, Argentina. J. Vert. Paleont. 2021, 40, e1877151. [Google Scholar] [CrossRef]
- Gianechini, F.A.; Apesteguía, S.; Landini, W.; Finotti, F.; Juárez Valieri, R.; Zandonai, F. New abelisaurid remains from the Anacleto Formation (Upper Cretaceous), Patagonia, Argentina. Cret. Res. 2015, 54, 1–16. [Google Scholar] [CrossRef]
- Allain, R.; Pereda-Suberbiola, X. Dinosaurs of France. C. R. Palevol. 2003, 2, 27–44. [Google Scholar] [CrossRef]
- Ezcurra, M.D.; Agnolin, F.L. A new global palaeobiogeographical model for the Late Mesozoic and Early Tertiary. Syst. Biol. 2012, 61, 553–566. [Google Scholar] [CrossRef]
- Agnolin, F. History of Cenozoic Mammals from South America. A New Model; Springer: Chur, Switzerland, 2024; 318p. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buffetaut, E.; Tong, H.; Girard, J.; Hoyez, B.; Párraga, J. Caletodraco cottardi: A New Furileusaurian Abelisaurid (Dinosauria: Theropoda) from the Cenomanian Chalk of Normandy (North-Western France). Foss. Stud. 2024, 2, 177-195. https://doi.org/10.3390/fossils2030009
Buffetaut E, Tong H, Girard J, Hoyez B, Párraga J. Caletodraco cottardi: A New Furileusaurian Abelisaurid (Dinosauria: Theropoda) from the Cenomanian Chalk of Normandy (North-Western France). Fossil Studies. 2024; 2(3):177-195. https://doi.org/10.3390/fossils2030009
Chicago/Turabian StyleBuffetaut, Eric, Haiyan Tong, Jérôme Girard, Bernard Hoyez, and Javier Párraga. 2024. "Caletodraco cottardi: A New Furileusaurian Abelisaurid (Dinosauria: Theropoda) from the Cenomanian Chalk of Normandy (North-Western France)" Fossil Studies 2, no. 3: 177-195. https://doi.org/10.3390/fossils2030009
APA StyleBuffetaut, E., Tong, H., Girard, J., Hoyez, B., & Párraga, J. (2024). Caletodraco cottardi: A New Furileusaurian Abelisaurid (Dinosauria: Theropoda) from the Cenomanian Chalk of Normandy (North-Western France). Fossil Studies, 2(3), 177-195. https://doi.org/10.3390/fossils2030009