The Effect of Green and Black Tea Polyphenols on BRCA2 Deficient Chinese Hamster Cells by Synthetic Lethality through PARP Inhibition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
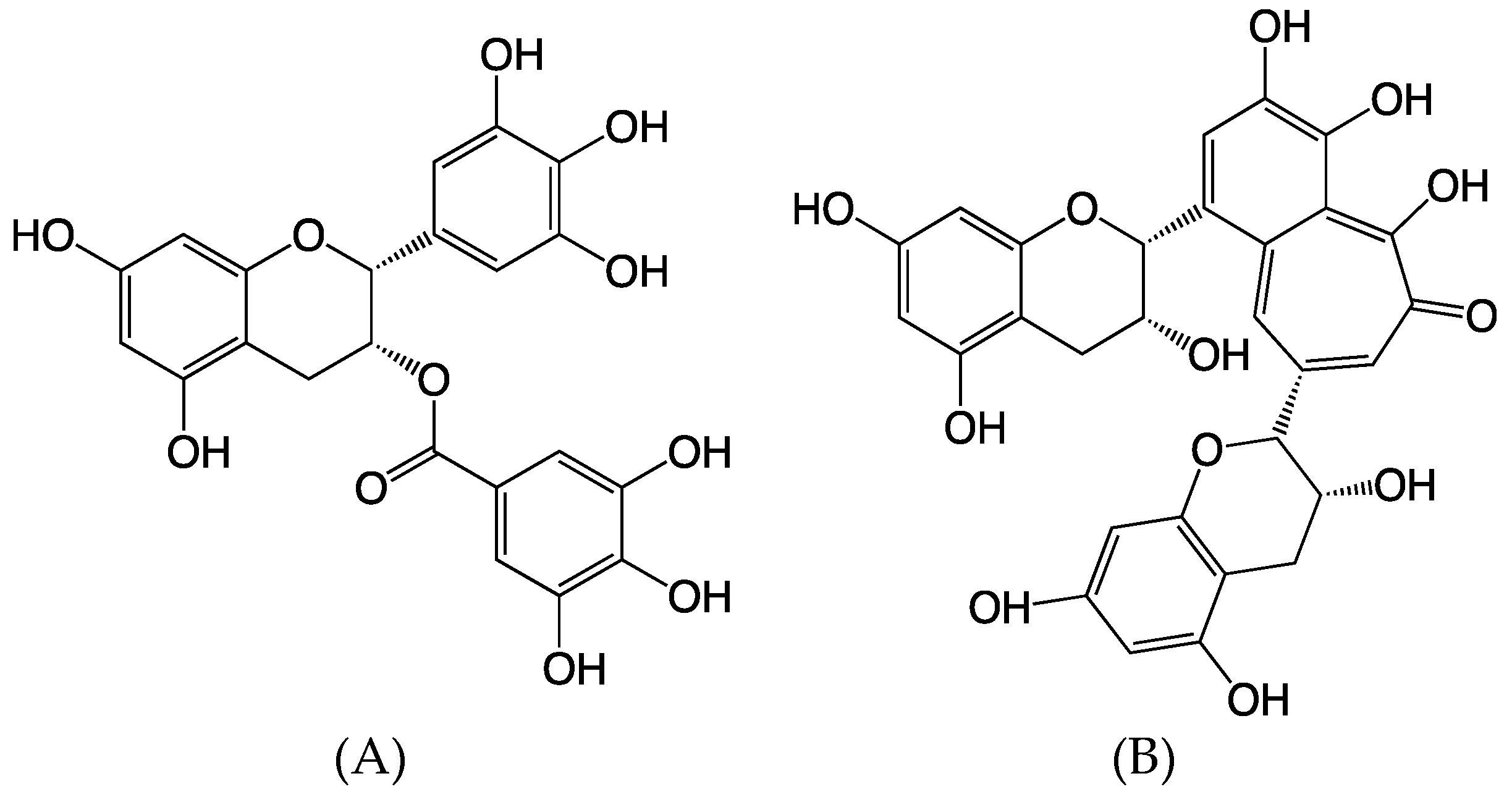
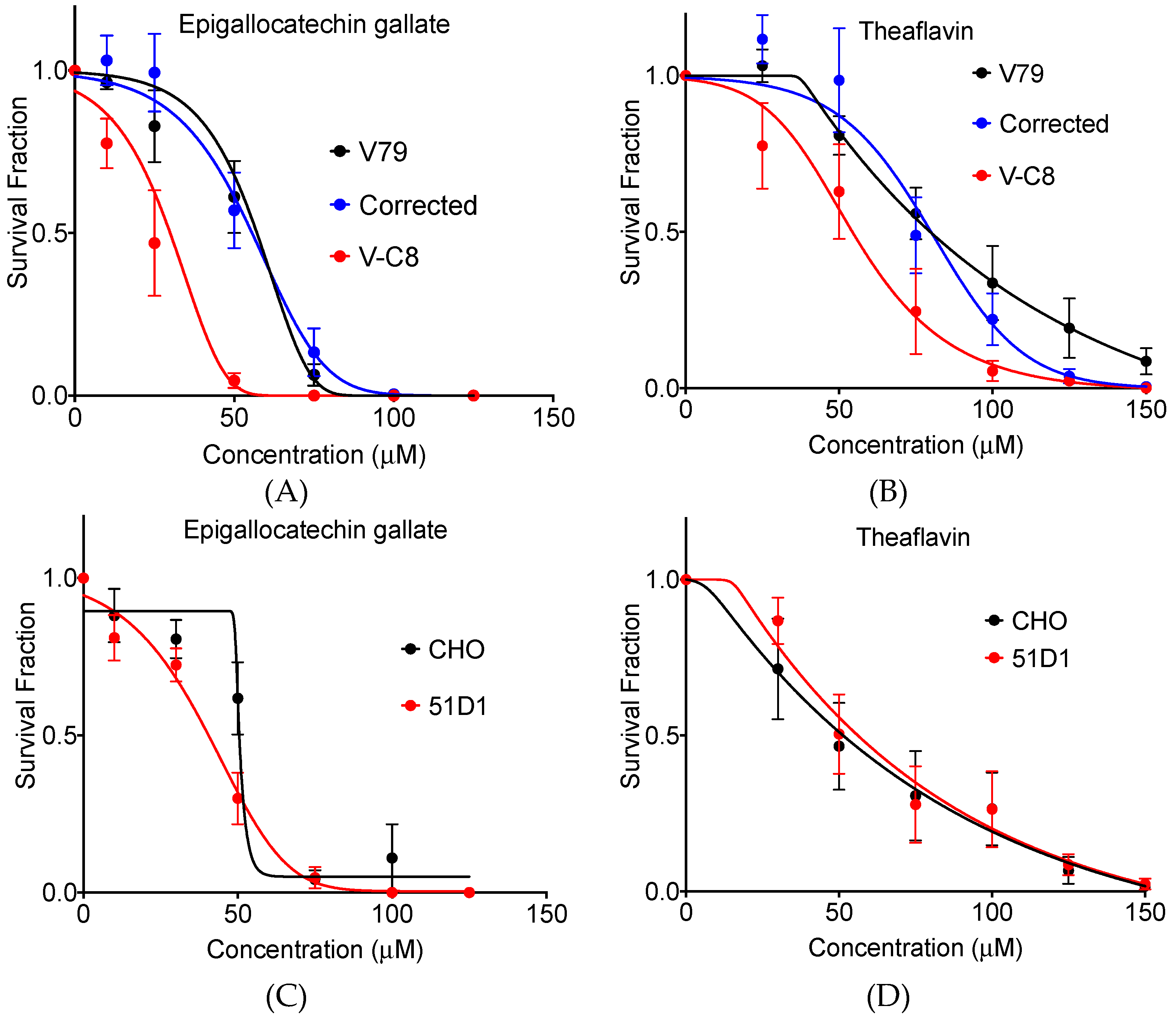
2.1. Clonogenic Cell Survival
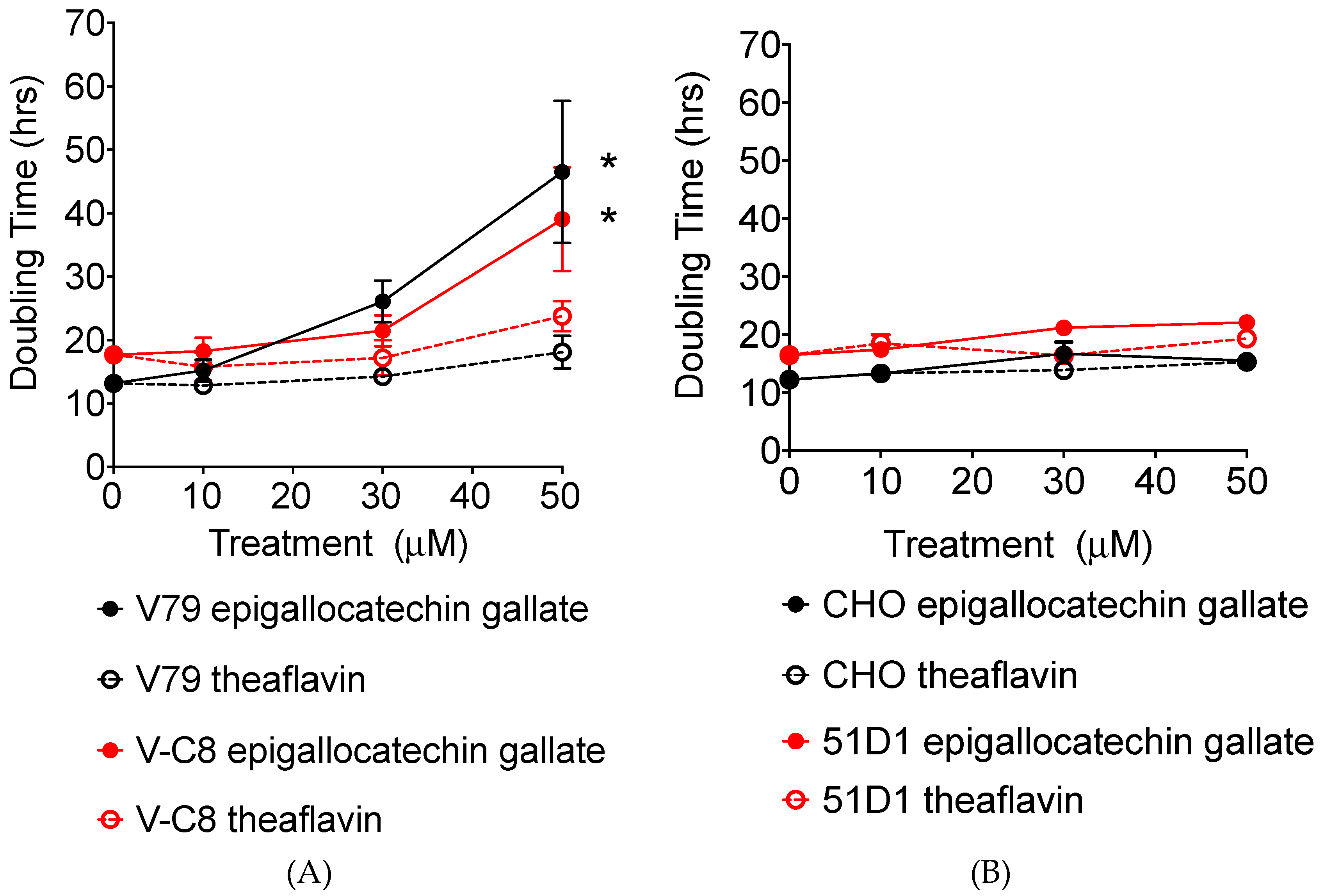
2.2. Growth Delay and Cell Cycle Analysis
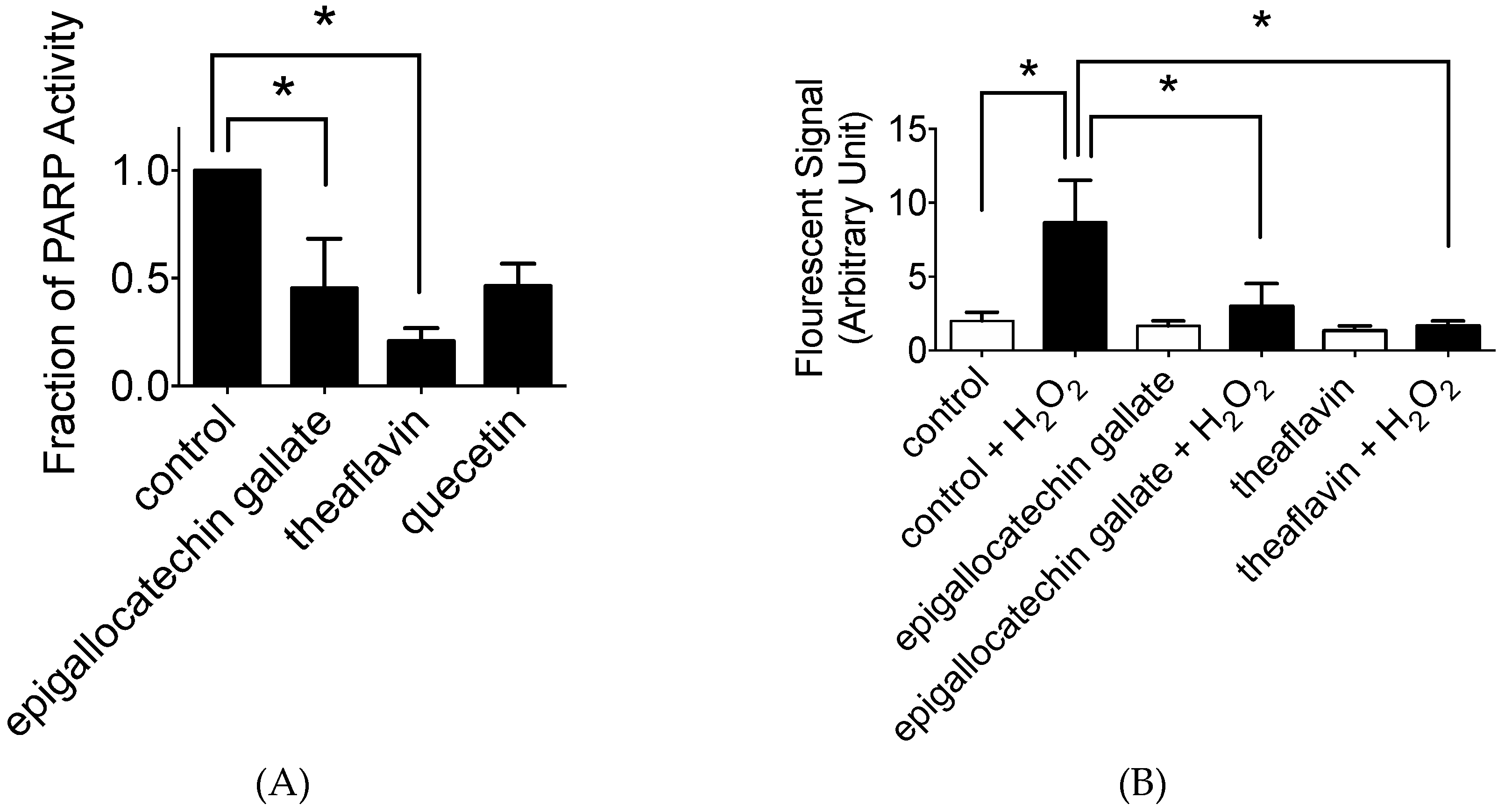
2.3. In Vitro and In Vivo PARP Activity Inhibition
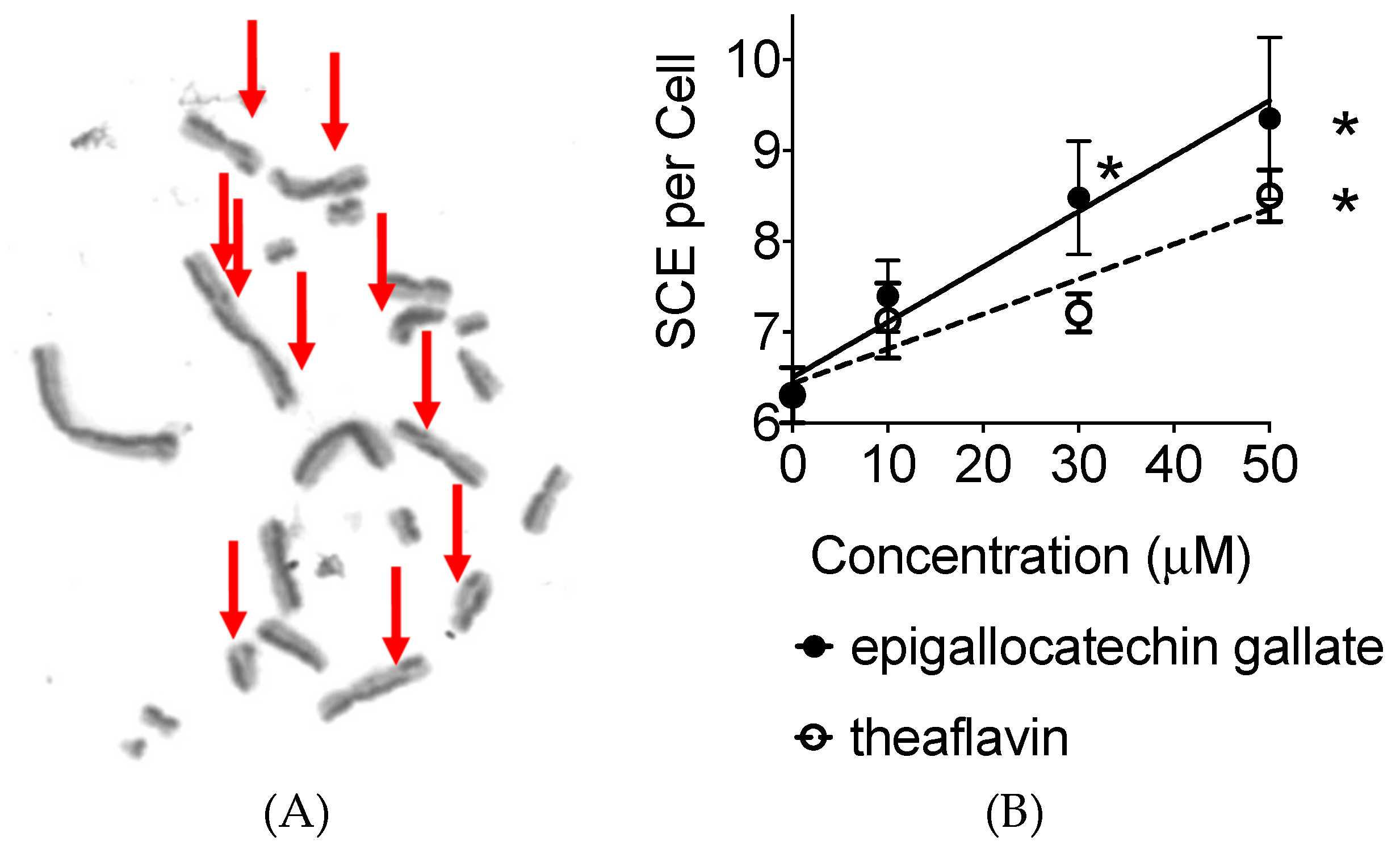
2.4. Sister Chromatid Exchanges
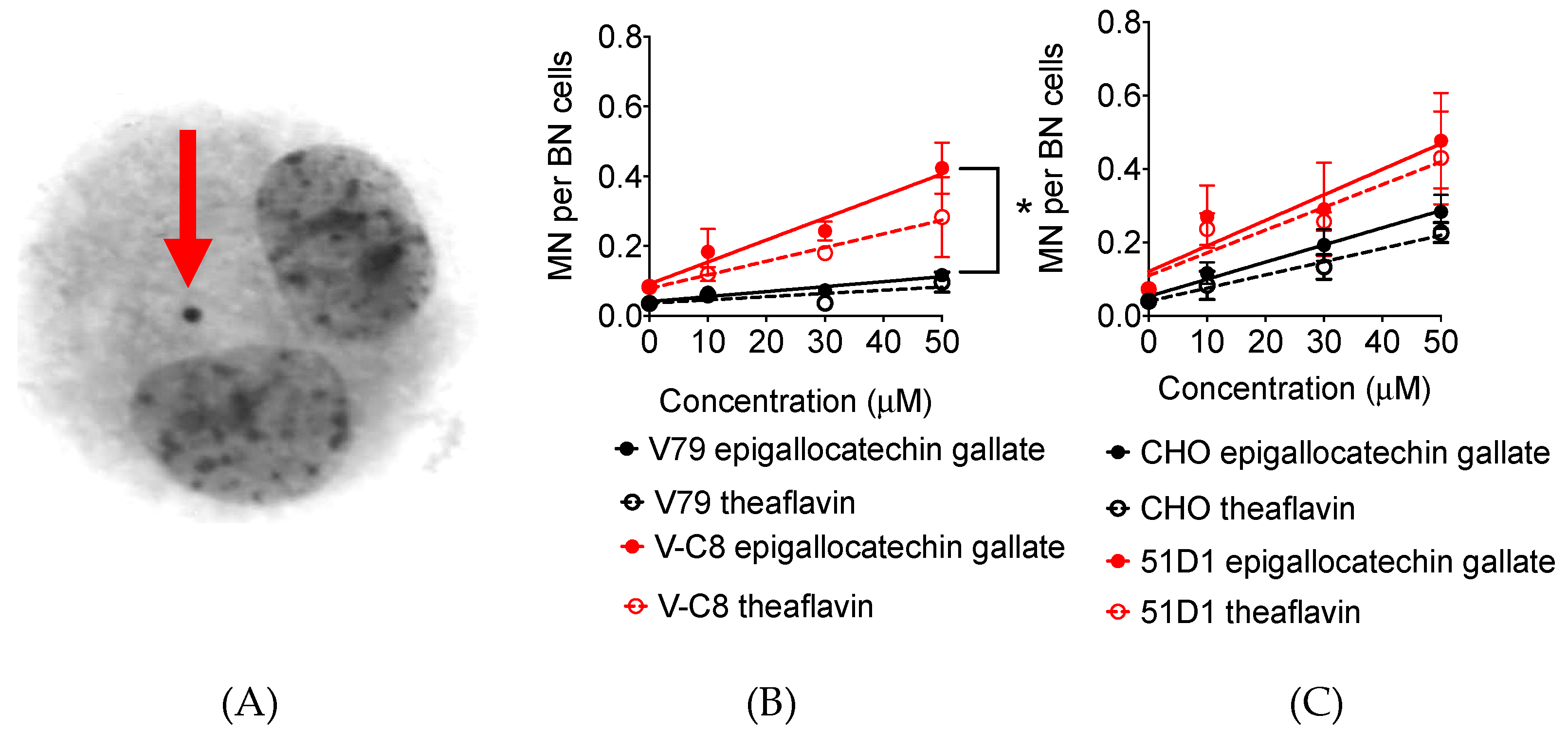
2.5. Micronuclei Formation
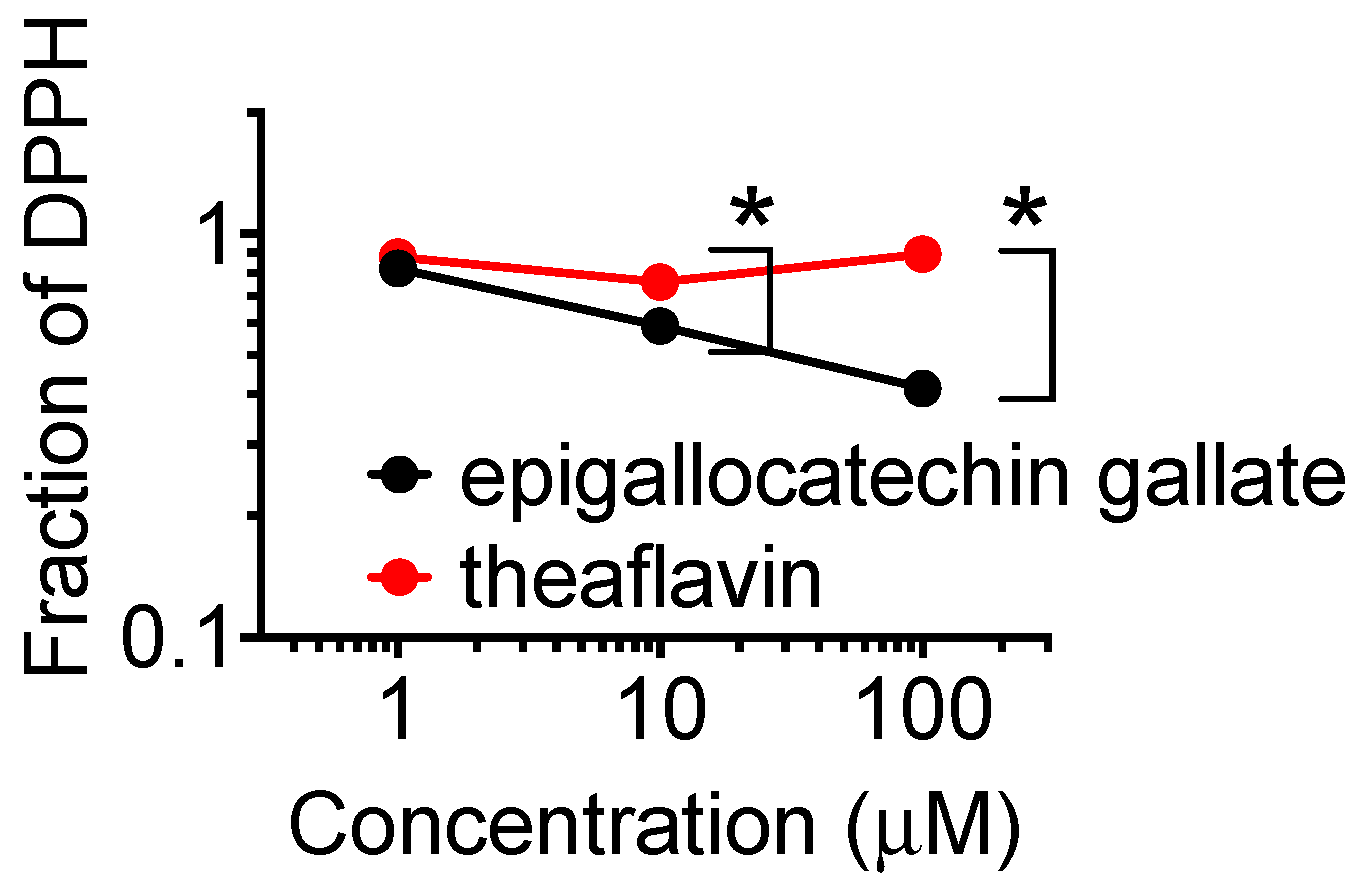
2.6. DPPH Radical Scavenging Capacity
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Chemicals
4.3. Clonogenic Cell Survival
4.4. Growth Inhibition
4.5. In Vitro and In Vivo PARP Activity Inhibition Assay
4.6. Sister Chromatid Exchange
4.7. Micronuclei Formation Assay
4.8. DPPH Analysis
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CHO | Chinese Hamster Ovary |
| BRCA2 | Breast Cancer Associate Gene 2 |
| PARP | Poly (ADP-ribose) polymerase |
| SCE | Sister Chromatid Exchange |
| HR | Homologous Recombination |
| NHEJ | Non-Homologous End Joining |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ANOVA | Analysis of valiance |
References
- Graham, H.N. Green tea composition, consumption, and polyphenol chemistry. Prev. Med. 1992, 21, 334–350. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, M.A.; Mulder, T.P.J.; Molhuizen, H.O.F. Theaflavins from Black Tea, Especially Theaflavin-3-gallate, Reduce the Incorporation of Cholesterol into Mixed Micelles. J. Agric. Food Chem. 2008, 56, 12031–12036. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.J.; Mullen, W.; Crozier, A. On-line high-performance liquid chromatography analysis of the antioxidant activity of phenolic compounds in green and black tea. Mol. Nutr. Food Res. 2005, 49, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Nakachi, K.; Suemasu, K.; Suga, K.; Takeo, T.; Imai, K.; Higashi, Y. Influence of drinking green tea on breast cancer malignancy among Japanese patients. Jpn. J. Cancer res. Gann 1998, 89, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.L.; Yuan, J.M.; Koh, W.P.; Yu, M.C. Green tea, black tea and breast cancer risk: A meta-analysis of epidemiological studies. Carcinogenesis 2006, 27, 1310–1315. [Google Scholar] [CrossRef] [PubMed]
- Romano, B.; Pagano, E.; Montanaro, V.; Fortunato, A.L.; Milic, N.; Borrelli, F. Novel insights into the pharmacology of flavonoids. Phytother. Res. PTR 2013, 27, 1588–1596. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Parshad, R.; Sanford, K.K.; Price, F.M.; Steele, V.E.; Tarone, R.E.; Kelloff, G.J.; Boone, C.W. Protective action of plant polyphenols on radiation-induced chromatid breaks in cultured human cells. Anticancer Res. 1998, 18, 3263–3266. [Google Scholar]
- Maeda, J.; Roybal, E.J.; Brents, C.A.; Uesaka, M.; Aizawa, Y.; Kato, T.A. Natural and glucosyl flavonoids inhibit poly(ADP-ribose) polymerase activity and induce synthetic lethality in BRCA mutant cells. Oncol. Rep. 2014, 31, 551–556. [Google Scholar] [CrossRef]
- Chang, H.L.; Su, J.H.; Yeh, Y.T.; Lee, Y.C.; Chen, H.M.; Wu, Y.C.; Yuan, S.S. Protoapigenone, a novel flavonoid, inhibits ovarian cancer cell growth in vitro and in vivo. Cancer Lett. 2008, 267, 85–95. [Google Scholar] [CrossRef]
- Herceg, Z.; Wang, Z.Q. Functions of poly(ADP-ribose) polymerase (PARP) in DNA repair, genomic integrity and cell death. Mut. Res. 2001, 477, 97–110. [Google Scholar] [CrossRef]
- Helleday, T. The underlying mechanism for the PARP and BRCA synthetic lethality: clearing up the misunderstandings. Mol. Oncol. 2011, 5, 387–393. [Google Scholar] [CrossRef]
- Kaufman, B.; Shapira-Frommer, R.; Schmutzler, R.K.; Audeh, M.W.; Friedlander, M.; Balmaña, J.; Mitchell, G.; Fried, G.; Stemmer, S.M.; Hubert, A.; et al. Olaparib monotherapy in patients with advanced cancer and a germline BRCA1/2 mutation. J. Clin.Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 244–250. [Google Scholar] [CrossRef]
- Thakur, V.S.; Gupta, K.; Gupta, S. Green tea polyphenols causes cell cycle arrest and apoptosis in prostate cancer cells by suppressing class I histone deacetylases. Carcinogenesis 2012, 33, 377–384. [Google Scholar] [CrossRef]
- Su, C.; Haskins, A.H.; Omata, C.; Aizawa, Y.; Kato, T.A. PARP Inhibition by Flavonoids Induced Selective Cell Killing to BRCA2-Deficient Cells. Pharmaceuticals 2017, 10. [Google Scholar] [CrossRef]
- Bryant, H.E.; Schultz, N.; Thomas, H.D.; Parker, K.M.; Flower, D.; Lopez, E.; Kyle, S.; Meuth, M.; Curtin, N.J.; Helleday, T. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature 2005, 434, 913–917. [Google Scholar] [CrossRef]
- Bandele, O.J.; Clawson, S.J.; Osheroff, N. Dietary polyphenols as topoisomerase II poisons: B ring and C ring substituents determine the mechanism of enzyme-mediated DNA cleavage enhancement. Chem. Res. Toxicol. 2008, 21, 1253–1260. [Google Scholar] [CrossRef]
- Bassett, S.A.; Barnett, M.P. The role of dietary histone deacetylases (HDACs) inhibitors in health and disease. Nutrients 2014, 6, 4273–4301. [Google Scholar] [CrossRef]
- Park, S. Polyphenol Compound as a Transcription Factor Inhibitor. Nutrients 2015, 7, 8987–9004. [Google Scholar] [CrossRef] [Green Version]
- Little, J.B.; Nagasawa, H.; Kennedy, A.R. DNA repair and malignant transformation: effect of X irradiation, 12-O-tetradecanoyl-phorbol-13-acetate, and protease inhibitors on transformation and sister-chromatid exchanges in mouse 10T 1/2 cells. Radiat. Res. 1979, 79, 241–255. [Google Scholar] [CrossRef]
- Maeda, J.; Yurkon, C.R.; Fujii, Y.; Fujisawa, H.; Kato, S.; Brents, C.A.; Uesaka, M.; Fujimori, A.; Kitamura, H.; Kato, T.A. Solution Radioactivated by Hadron Radiation Can Increase Sister Chromatid Exchanges. PLoS ONE 2015, 10, e0144619. [Google Scholar] [CrossRef]
- Su, C.; Gius, J.P.; Van Steenberg, J.; Haskins, A.H.; Heishima, K.; Omata, C.; Iwayama, M.; Murakami, M.; Mori, T.; Maruo, K.; et al. Hypersensitivity of BRCA2 deficient cells to rosemary extract explained by weak PARP inhibitory activity. Sci. Rep. 2017, 7, 16704. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Iritani, K.; Yonemori, S.; Oyama, Y.; Takeda, Y. Isolation and antioxidant activity of galloyl flavonol glycosides from the seashore plant, Pemphis acidula. Biosci. Biotechnol. Biochem. 2001, 65, 1302–1309. [Google Scholar] [CrossRef]
- Engen, A.; Maeda, J.; Wozniak, D.E.; Brents, C.A.; Bell, J.J.; Uesaka, M.; Aizawa, Y.; Kato, T.A. Induction of cytotoxic and genotoxic responses by natural and novel quercetin glycosides. Mut. Res. Gen. Toxicol. Environ. Mutagen. 2015, 784–785, 15–22. [Google Scholar] [CrossRef]
- Verhaegh, G.W.; Jongmans, W.; Morolli, B.; Jaspers, N.G.; van der Schans, G.P.; Lohman, P.H.; Zdzienicka, M.Z. A novel type of X-ray-sensitive Chinese hamster cell mutant with radioresistant DNA synthesis and hampered DNA double-strand break repair. Mut. Res. 1995, 337, 119–129. [Google Scholar] [CrossRef]
- Hinz, J.M.; Tebbs, R.S.; Wilson, P.F.; Nham, P.B.; Salazar, E.P.; Nagasawa, H.; Urbin, S.S.; Bedford, J.S.; Thompson, L.H. Repression of mutagenesis by Rad51D-mediated homologous recombination. Nucleic Acids Res. 2006, 34, 1358–1368. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.H.; Read, R.A.; Bedford, J.S. Comparison of synchronized Chinese hamster ovary cells obtained by mitotic shake-off, hydroxyurea, aphidicolin, or methotrexate. Cytometry 1987, 8, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Tobey, R.A.; Ley, K.D. Regulation of initiation of DNA synthesis in Chinese hamster cells. I. Production of stable, reversible G1-arrested populations in suspension culture. J. Cell Biol. 1970, 46, 151–157. [Google Scholar] [CrossRef]
- Cartwright, I.M.; Genet, M.D.; Kato, T.A. A simple and rapid fluorescence in situ hybridization microwave protocol for reliable dicentric chromosome analysis. J. Radiat. Res. 2013, 54, 344–348. [Google Scholar] [CrossRef]
- Perry, P.; Wolff, S. New Giemsa method for the differential staining of sister chromatids. Nature 1974, 251, 156–158. [Google Scholar] [CrossRef]
- Haskins, A.H.; Su, C.; Engen, A.; Salinas, V.A.; Maeda, J.; Uesaka, M.; Aizawa, Y.; Kato, T.A. Data for induction of cytotoxic response by natural and novel quercetin glycosides. Data Brief 2016, 6, 262–266. [Google Scholar] [CrossRef]
- Su, C.; Allum, A.J.; Aizawa, Y.; Kato, T.A. Novel glyceryl glucoside is a low toxic alternative for cryopreservation agent. Biochem. Biophys. Res. Commun. 2016, 476, 359–364. [Google Scholar] [CrossRef]
- Yu, H.; Haskins, J.S.; Su, C.; Allum, A.; Haskins, A.H.; Salinas, V.A.; Sunada, S.; Inoue, T.; Aizawa, Y.; Uesaka, M.; et al. In vitro screening of radioprotective properties in the novel glucosylated flavonoids. Int. J. Mol. Med. 2016, 38, 1525–1530. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqahtani, S.; Welton, K.; Gius, J.P.; Elmegerhi, S.; Kato, T.A. The Effect of Green and Black Tea Polyphenols on BRCA2 Deficient Chinese Hamster Cells by Synthetic Lethality through PARP Inhibition. Int. J. Mol. Sci. 2019, 20, 1274. https://doi.org/10.3390/ijms20061274
Alqahtani S, Welton K, Gius JP, Elmegerhi S, Kato TA. The Effect of Green and Black Tea Polyphenols on BRCA2 Deficient Chinese Hamster Cells by Synthetic Lethality through PARP Inhibition. International Journal of Molecular Sciences. 2019; 20(6):1274. https://doi.org/10.3390/ijms20061274
Chicago/Turabian StyleAlqahtani, Shaherah, Kelly Welton, Jeffrey P. Gius, Suad Elmegerhi, and Takamitsu A. Kato. 2019. "The Effect of Green and Black Tea Polyphenols on BRCA2 Deficient Chinese Hamster Cells by Synthetic Lethality through PARP Inhibition" International Journal of Molecular Sciences 20, no. 6: 1274. https://doi.org/10.3390/ijms20061274
APA StyleAlqahtani, S., Welton, K., Gius, J. P., Elmegerhi, S., & Kato, T. A. (2019). The Effect of Green and Black Tea Polyphenols on BRCA2 Deficient Chinese Hamster Cells by Synthetic Lethality through PARP Inhibition. International Journal of Molecular Sciences, 20(6), 1274. https://doi.org/10.3390/ijms20061274