Anti-Obesity Effects of Lactobacillus fermentum CQPC05 Isolated from Sichuan Pickle in High-Fat Diet-Induced Obese Mice through PPAR-α Signaling Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Strain
2.2. Establishment of Animal Model
2.3. Determination of Alanine Aminotransferase (ALT), Aspartate Aminotransaminase (AST), Total Cholesterol (TC), Triglyceride (TG), High-Density Lipoprotein Cholesterol (HDL-C), and Low-Density Lipoprotein Cholesterol (LDL-C) Levels in Serum and Liver Samples
2.4. Pathological Observation of Liver Tissue and Epididymal Adipose Tissue
2.5. Quantitative PCR (qPCR) Assay
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
3.1. Resistance of Lactic Acid Bacteria to Artificial Gastric Juice and Bile Salt
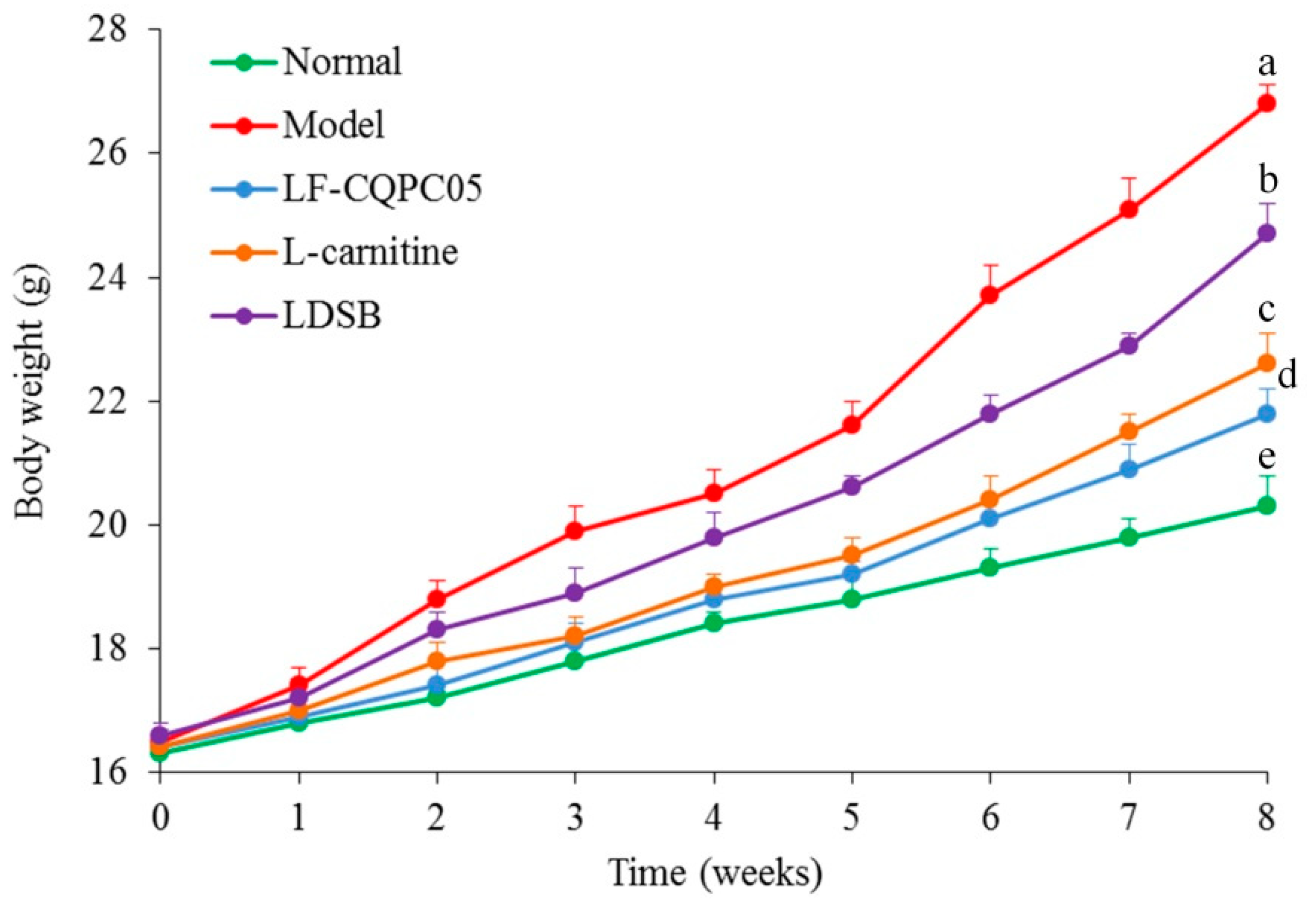
3.2. Body Weight of Mice
3.3. Organ Indices of Mice
3.4. Levels of ALT, AST, TC, TG, HDL-C, and LDL-C in the Serum of Mice
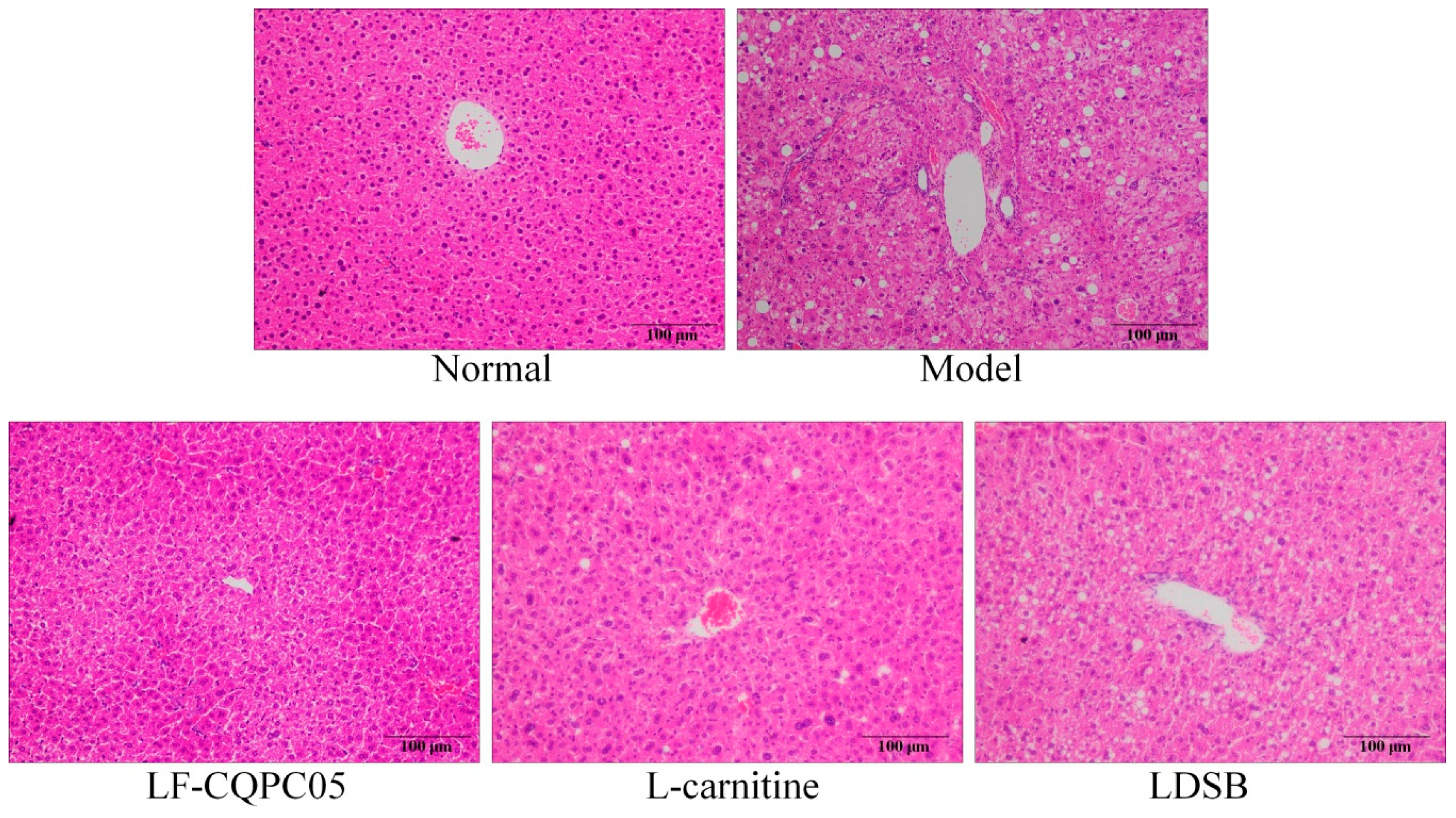
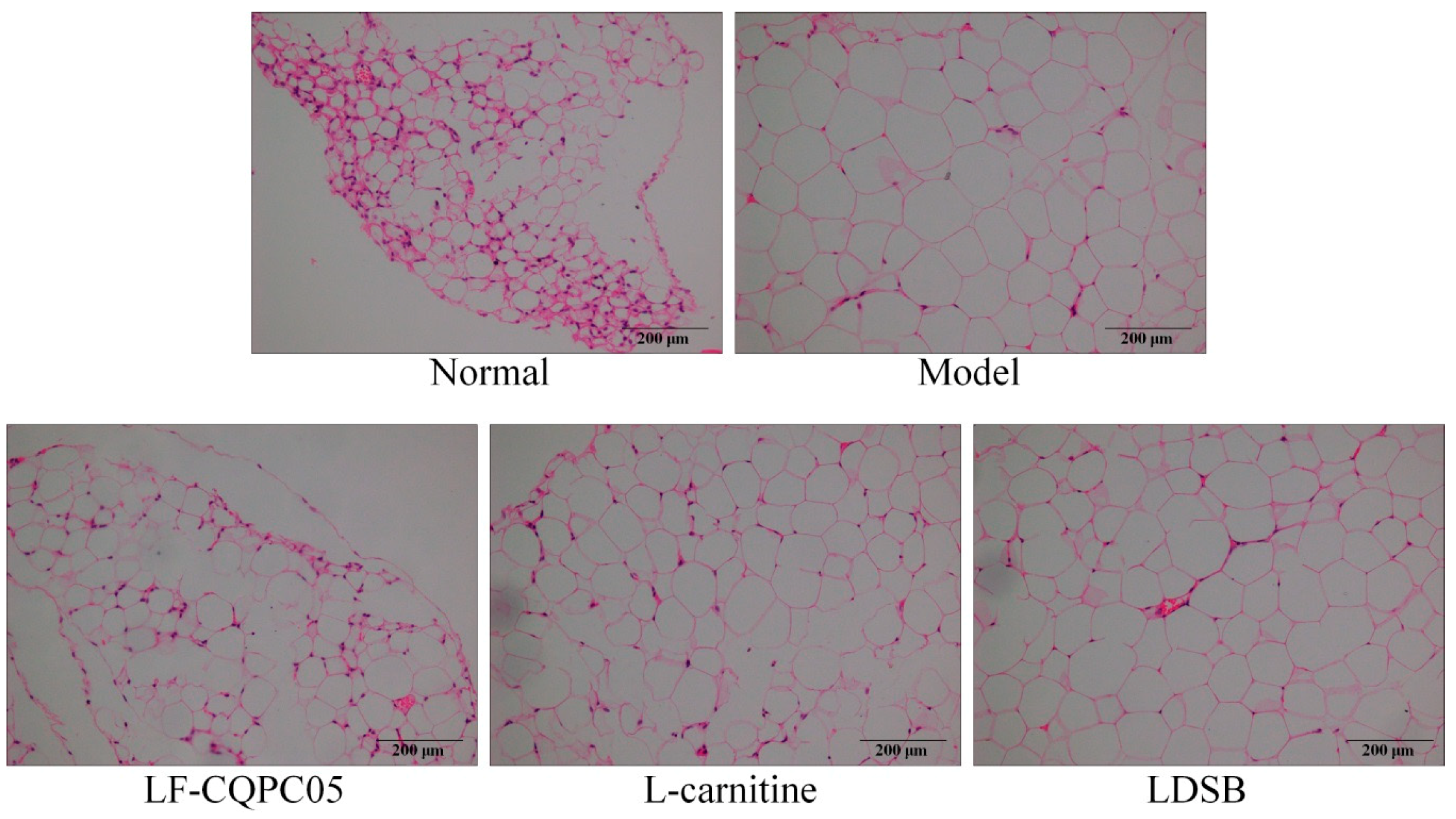
3.5. Pathological Observation of Liver and Epididymal Adipose Tissue of Mice
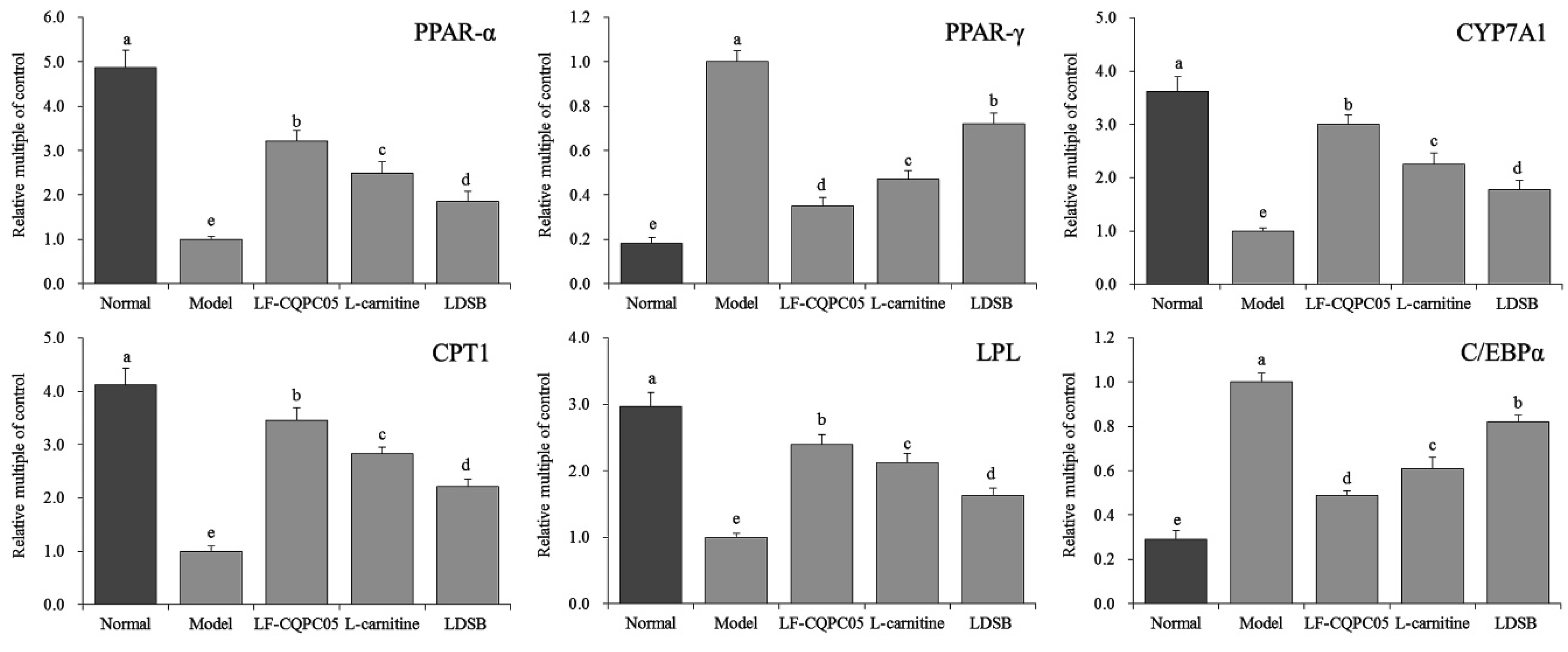
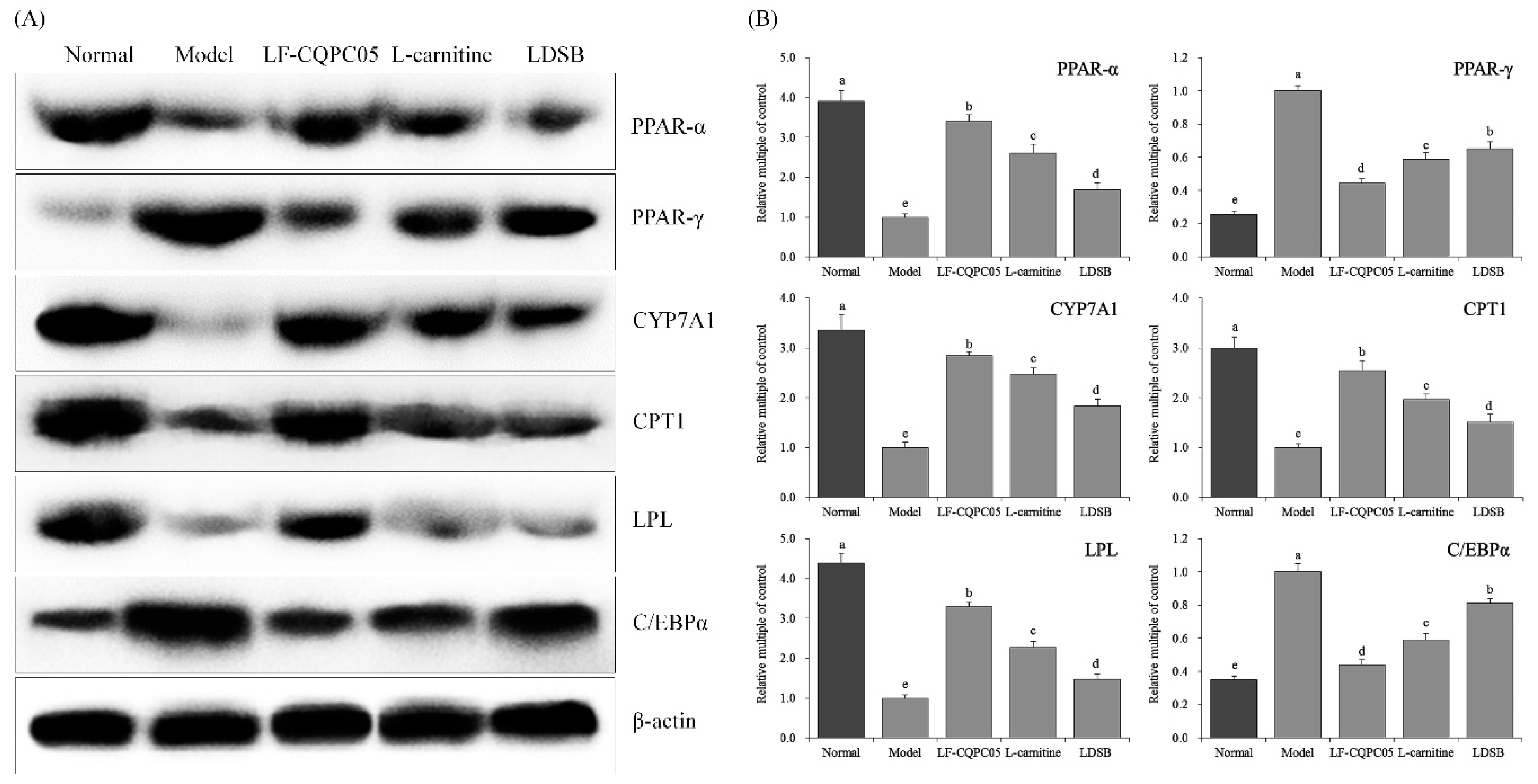
3.6. Expression of RNA and Protein in Mouse Liver Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liu, B.H.; Yang, X.; Guo, L.J.; Zhang, J.; Zhou, X.R.; Yi, R.K.; Zhao, X. Inhibitory effect of Lactobacillus plantarum CQPC02 isolated from Chinese Sichuan pickles (Paocai) on constipation in mice. J. Food Qual. 2019, 2019, 9781914. [Google Scholar] [CrossRef]
- Yang, J.X.; Zhang, L.L.; Jiang, H.Y.; He, Z.F. Isolation and identification of lactic bacteria in pickled vegetavles from Meishan city. Food Sci. 2015, 36, 158–163. [Google Scholar]
- Liang, H.; Yin, L.; Zhang, Y.; Chang, C.; Zhang, W. Dynamics and diversity of a microbial community during the fermentation of industrialized Qingcai paocai, a traditional Chinese fermented vegetable food, as assessed by Illumina MiSeq sequencing, DGGE and qPCR assay. Ann. Microbiol. 2018, 68, 111–122. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Yi, R.; Mu, J.; Zhao, X.; Yang, Z. Hepatoprotective effects of Lactobacillus on carbon tetrachloride-induced acute liver injury in mice. Int. J. Mol. Sci. 2018, 19, 2212. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Díaz-Ropero, M.A.; Gómez, N.; Lara-Villoslada, F.; Sierra, S.; Maldonado, J.A.; Martín, R.; López-Huertas, E.; Rodríguez, J.M.; Xaus, J. Oral administration of two probiotic strains, Lactobacillus gasseri CECT5714 and Lactobacillus coryniformis CECT5711, enhances the intestinal function of healthy adults. Int. J. Food Microbiol. 2006, 107, 104–111. [Google Scholar] [CrossRef]
- Qian, Y.; Song, J.-L.; Yi, R.; Li, G.; Sun, P.; Zhao, X.; Huo, G. Preventive effects of Lactobacillus plantarum YS4 on constipation induced by activated carbon in mice. Appl. Sci. 2018, 8, 363. [Google Scholar] [CrossRef]
- Chen, G.; Yu, W.H.; Zhang, Q.S.; Song, P.; Zhang, B.B.; Liu, Z.; You, J.G.; Li, H. Research of Sichuan Paocai and Lactic Acid Bacteria. Adv. J. Food Sci. Technol. 2014, 6, 1–5. [Google Scholar]
- Huang, Y.; Luo, Y.B.; Zhai, Z.Y.; Zhang, H.X.; Yang, C.X.; Tian, H.T.; Li, Z.; Feng, J.N.; Liu, H.; Hao, Y.L. Characterization and application of an anti-Listeria bacteriocin produced by Pediococcus pentosaceus 05–10 isolated from Sichuan Pickle, a traditionally fermented vegetable product from China. Food Control. 2009, 20, 1030–1035. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, X.; Wang, H.; Yang, Z.; Li, J.; Suo, H. Prevent effects of Lactobacillus fermentum HY01 on dextran sulfate sodium-induced colitis in mice. Nutrients 2017, 9, 545. [Google Scholar] [CrossRef]
- Chen, X.; Tan, F.; Yi, R.; Mu, J.; Zhao, X.; Yang, Z. Effects of Lactobacillus on mice with diabetes induced by high-fat diet with streptozotocin (STZ). Appl. Sci. 2018, 8, 1249. [Google Scholar] [CrossRef]
- Ren, D.Y.; Li, C.; Qin, Y.Q.; Jin, N.Y. Development review on healthy function and potential mechanisms of lactic acid bacteria. Chin. J. Vet. Drug 2011, 45, 47–50. [Google Scholar]
- Chen, D.W.; Yang, Z.Q.; Chen, X.; Huang, Y.J.; Yin, B.X.; Guo, F.X.; Zhao, H.Q.; Zhao, T.Y.; Qu, H.X.; Huang, J.D.; et al. The effect of Lactobacillus rhamnosus hsryfm 1301 on the intestinal microbiota of a hyperlipidemic rat model. BMC Complement. Altern. Med. 2014, 14, 386. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, X.Q.; Chen, C.C.; Wang, J.L.; Zou, X.; Li, C.W.; Xu, Z.C.; Yang, X.Y.; Shi, W.B.; Zeng, C.Y. Oxidative stress causes imbalance of renal renin angiotensin system (RAS) components and hypertension in obese Zucker rats. J. Am. Heart Assoc. 2015, 4, e001559. [Google Scholar] [CrossRef] [PubMed]
- Alisi, A.; Bedogni, G.; Baviera, G.; Giorgio, V.; Porro, E.; Paris, C.; Giammaria, P.; Reali, L.; Anania, F.; Nobili, V. Randomised clinical trial: The beneficial effects of VSL#3 in obese children with non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2014, 39, 1276–1285. [Google Scholar] [PubMed]
- Clément, K.; Ferré, P. Genetics and the pathophysiology of obesity. Pediatr. Res. 2003, 53, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Dayan, P.H.; Sforzo, G.; Boisseau, N.; Pereira-Lancha, L.O.; Lancha, A.H., Jr. A new clinical perspective: Treating obesity with nutritional coaching versus energy-restricted diets. Nutrition 2019, 60, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Finicelli, M.; Squillaro, T.; Di Cristo, F.; Di Salle, A.; Melone, M.A.B.; Galderisi, U.; Peluso, G. Metabolic syndrome, Mediterranean diet, and polyphenols: Evidence and perspectives. J. Cell Physiol. 2019, 234, 5807–5826. [Google Scholar] [CrossRef] [PubMed]
- Asano, M.; Iwagaki, Y.; Sugawara, S.; Kushida, M.; Okouchi, R.; Yamamoto, K.; Tsuduki, T. Effects of Japanese diet in combination with exercise on visceral fat accumulation. Nutrition 2019, 57, 173–182. [Google Scholar] [CrossRef]
- Appiakannan, H.S.; Kestyus, D.R.; Weber, E.T. Effects of high fat diet and chronic circadian challenge on glucocorticoid regulation in C57BL/6J mice. Physiol. Behav. 2019, 204, 100–105. [Google Scholar] [CrossRef]
- Corrêa, T.A.; Rogero, M.M. Polyphenols regulating microRNAs and inflammation biomarkers in obesity. Nutrition 2019, 59, 150–157. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeong, D.; Kang, I.B.; Kim, H.; Song, K.Y.; Seo, K.H. Dual function of Lactobacillus kefiri DH5 in preventing high-fat-diet-induced obesity: Direct reduction of cholesterol and upregulation of PPAR-α in adipose tissue. Mol. Nutr. Food Res. 2017, 61, 1700252. [Google Scholar] [CrossRef] [PubMed]
- Maleki Kakelar, H.; Barzegari, A.; Hanifian, S.; Barar, J.; Omidi, Y. Isolation and molecular identification of Lactobacillus with probiotic potential from abomasums driven rennet. Food Chem. 2019, 272, 709–714. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, X.; Chen, B.; Long, X.; Mu, J.; Pan, Y.; Song, J.L.; Zhao, X.; Yang, Z. Preventive effect of Lactobacillus plantarum CQPC10 on activated carbon induced constipation in Institute of Cancer Research (ICR) Mice. Appl. Sci. 2018, 8, 1498. [Google Scholar] [CrossRef]
- Tanida, M.; Shen, J.; Maeda, K.; Horii, Y.; Yamano, T.; Fukushima, Y.; Nagai, K. High-fat diet-induced obesity is attenuated by probiotic strain Lactobacillus paracasei ST11 (NCC2461) in rats. Obes Res. Clin. Pract. 2008, 2, 159–169. [Google Scholar] [CrossRef]
- Rather, S.A.; Pothuraju, R.; Sharma, R.K.; De, S.; Mir, N.A.; Jangra, S. Anti-obesity effect of feeding probiotic dahi containing Lactobacillus casei NCDC 19 in high fat diet-induced obese mice. Int. J. Dairy Technol. 2014, 67, 504–509. [Google Scholar] [CrossRef]
- Toral, M.; Gómez-Guzmán, M.; Jiménez, R.; Romero, M.; Sánchez, M.; Utrilla, M.P.; Garrido-Mesa, N.; Rodríguez-Cabezas, M.E.; Olivares, M.; Gálvez, J.; et al. The probiotic Lactobacillus coryniformis CECT5711 reduces the vascular pro-oxidant and pro-inflammatory status in obese mice. Clin. Sci. 2014, 127, 33–45. [Google Scholar] [CrossRef]
- Thornton, B.; Basu, C. Real-time PCR (qPCR) primer design using free online software. Biochem. Mol. Biol. Educ. 2011, 39, 145–154. [Google Scholar] [CrossRef]
- Bustina, S.; Huggett, J. qPCR primer design revisited. Biomol. Detect. Quantif. 2017, 14, 19–28. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, J.; Zhou, X.; Yi, R.; Mu, J.; Long, X.; Pan, Y.; Zhao, X.; Liu, W. Lactobacillus plantarum CQPC11 isolated from sichuan pickled cabbages antagonizes d-galactose-induced oxidation and aging in mice. Molecules 2018, 23, 3026. [Google Scholar] [CrossRef]
- Pan, Y.; Long, X.; Yi, R.; Zhao, X. Polyphenols in Liubao tea can prevent CCl4-induced hepatic damage in mice through its antioxidant capacities. Nutrients 2018, 10, 1280. [Google Scholar] [CrossRef]
- Al Zarzour, R.H.; Ahmad, M.; Asmawi, M.Z.; Kaur, G.; Saeed, M.A.A.; Al-Mansoub, M.A.; Saghir, S.A.M.; Usman, N.S.; Al-Dulaimi, D.W.; Yam, M.F. Phyllanthus Niruri standardized extract alleviates the progression of non-alcoholic fatty liver disease and decreases atherosclerotic risk in Sprague-Dawley Rats. Nutrients 2017, 9, 766. [Google Scholar] [CrossRef] [PubMed]
- Panelli, M.F.; Pierine, D.T.; De Souza, S.L.B.; Ferron, A.J.T.; Garcia, J.L.; Santos, K.C.; Belin, M.A.F.; Lima, G.P.P.; Borguini, M.G.; Minatel, I.O.; et al. Bark of Passiflora edulis treatment stimulates antioxidant capacity, and reduces dyslipidemia and body fat in db/db mice. Antioxidants 2018, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, J.; Ye, J.; Guo, Q.; Wang, W.; Sun, Y.; Zeng, Q. Separate and combined associations of physical activity and obesity with lipid-related indices in non-diabetic and diabetic patients. Lipids Health Dis. 2019, 18, 49. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.X.; Sun, Y.; Dai, X.L.; Zhan, J.X.; Long, T.T.; Xiong, M.X.; Li, H.H.; Kuang, Q.; Tang, T.T.; Qin, Y.T.; et al. Fisetin attenuates high fat diet-triggered hepatic lipid accumulation: A mechanism involving liver inflammation overload associated TACE/TNF-α pathway. J. Funct. Foods 2019, 53, 7–21. [Google Scholar]
- Lee, S.; Han, D.; Kang, H.; Jeong, S.J.; Jo, J.E.; Shin, J.; Kim, D.K.; Park, H.W. Intravenous sustained-release nifedipine ameliorates nonalcoholic fatty liver disease by restoring autophagic clearance. Biomaterials 2019, 197, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Khajebishak, Y.; Payahoo, L.; Alivand, M.; Alipour, B. Punicic acid: A potential compound of pomegranate seed oil in Type 2 diabetes mellitus management. J. Cell. Physiol. 2019, 234, 2112–2120. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Kim, K.K.; Son, B.Y.; Nam, S.W.; Shin, P.G.; Kim, G.D. The anti-adipogenic activity of a new cultivar, Pleurotus eryngii var. ferulae ’Beesan No. 2’, through down-regulation of PPAR γ and C/EBP α in 3T3-L1 cells. J. Microbiol. Biotechnol. 2016, 26, 1836–1844. [Google Scholar] [CrossRef]
- Kim, H.J.; You, M.K.; Lee, Y.H.; Kim, H.J.; Adhikari, D.; Kim, H.A. Red pepper seed water extract inhibits preadipocyte differentiation and induces mature adipocyte apoptosis in 3T3-L1 cells. Nutr. Res. Pract. 2018, 12, 494–502. [Google Scholar] [CrossRef]
- Lundsgaard, A.M.; Fritzen, A.M.; Kiens, B. Molecular regulation of fatty acid oxidation in skeletal muscle during aerobic exercise. Trends Endocrinol. Metab. 2018, 29, 18–30. [Google Scholar] [CrossRef]
- Colom, C.; Viladés, D.; Pérez-Cuellar, M.; Leta, R.; Rivas-Urbina, A.; Carreras, G.; Ordóñez-Llanos, J.; Pérez, A.; Sánchez-Quesada, J.L. Associations between epicardial adipose tissue, subclinical atherosclerosis and high-density lipoprotein composition in type 1 diabetes. Cardiovasc. Diabetol. 2018, 17, 156. [Google Scholar] [CrossRef]
- Su, X.; Kong, Y.; Peng, D. New insights into apolipoprotein A5 in controlling lipoprotein metabolism in obesity and the metabolic syndrome patients. Lipids Health Dis. 2018, 17, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruciani-Guglielmacci, C.; Magnan, C. Brain lipoprotein lipase as a regulator of energy balance. Biochimie 2017, 143, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Hu, C.; Deng, X.; Bai, Y.; Cao, H.; Guo, J.; Su, Z. Therapeutic effect of chitooligosaccharide tablets on lipids in high-fat diets induced hyperlipidemic rats. Molecules 2019, 24, 514. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Ikeda, T.; Yamamoto, K.; Masuda, S.; Ogawa, H.; Kamisako, T. Gender-divergent expression of lipid and bile acid metabolism-related genes in adult mice offspring of dams fed a high-fat diet. J. Biosci. 2018, 43, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Stephens, R.W.; Arhire, L.; Covasa, M. Gut microbiota: From microorganisms to metabolic organ influencing obesity. Obesity 2018, 26, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Hegde, V.; Dhurandhar, N.V. Microbes and obesity--interrelationship between infection, adipose tissue and the immune system. Clin. Microbiol. Infect. 2013, 19, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Roselli, M.; Devirgiliis, C.; Zinno, P.; Guantario, B.; Finamore, A.; Rami, R.; Perozzi, G. Impact of supplementation with a food-derived microbial community on obesity-associated inflammation and gut microbiota composition. Genes Nutr. 2017, 12, 25. [Google Scholar] [CrossRef]
- Le Barz, M.; Anhê, F.F.; Varin, T.V.; Desjardins, V.; Levy, E.; Roy, D.; Urdaci, M.C.; Marette, A. Probiotics as complementary treatment for metabolic disorders. Diabetes Metab. J. 2015, 39, 291–303. [Google Scholar] [CrossRef]
- Anhê, F.F.; Nachbar, R.T.; Varin, T.V.; Trottier, J.; Dudonné, S.; Le Barz, M.; Feutry, P.; Pilon, G.; Barbier, O.; Desjardins, Y.; et al. Treatment with camu camu (Myrciaria dubia) prevents obesity by altering the gut microbiota and increasing energy expenditure in diet-induced obese mice. Gut 2019, 68, 453–464. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Ingredients | 10% Fat Mixed Diet (%) | 45% Fat Mixed Diet (%) |
|---|---|---|
| Corn flour | 24.89 | 7.79 |
| Bran | 15.00 | 7.00 |
| Wheatmeal | 7.00 | |
| Soybean meal | 18.50 | 22.00 |
| Sucrose | 16.38 | 20.00 |
| Lard | 1.02 | 19.50 |
| Premix | 17.21 | 23.71 |
| Gene Name | Sequence |
|---|---|
| PPAR-α | Forward: 5′-CCTCAGGGTACCACTACGGAGT-3′ |
| Reverse: 5′-GCCGAATAGTTCGCCGAA-3′ | |
| PPAR-γ | Forward: 5′-AGGCCGAGAAGGAGAAGCTGTTG-3′ |
| Reverse: 5′-TGGCCACCTCTTTGCTGTGCTC-3′ | |
| CYP7A1 | Forward: 5′-AGCAACTAAACAACCTGCCAGTACTA-3′ |
| Reverse: 5′-GTCCGGATATTCAAGGATGCA-3′ | |
| CPT1 | Forward: 5′-AAAGATCAATCGGACCCTAGACA-3′ |
| Reverse: 5′-CAGCGAGTAGCGCATAGTCA-3′ | |
| LPL | Forward: 5′-AGGGCTCTGCCTGAGTTGTA-3′ |
| Reverse: 5′-AGAAATCTCGAAGGCCTGGT-3′ | |
| C/EBPα | Forward: 5′-TGGACAAGAACAGCAACGAGTAC-3′ |
| Reverse: 5′-GCAGTTGCCCATGGCCTTGAC-3′ | |
| GAPDH | Forward: 5′-ACCCAGAAGACTGTGGATGG-3′ |
| Reverse: 5′-ACACATTGGGGGTAGGAACA-3′ |
| Treatment | Survival Rate in Artificial Gastric Juice at pH 3.0 (%) | Survival Rate in 0.3% Bile Salt (%) |
|---|---|---|
| LF-CQPC05 | 83.22 ± 3.61 | 13.37 ± 1.17 |
| LDSB | 41.33 ± 2.74 | 8.65 ± 0.56 |
| Group | 1st Week | 2nd Week | 3rd Week | 4th Week | 5th Week | 6th Week | 7th Week | 8th Week |
|---|---|---|---|---|---|---|---|---|
| Normal | 3.25 ± 0.11 a | 3.32 ± 0.14 a | 3.62 ± 0.13 a | 3.92 ± 0.15 a | 4.12 ± 0.19 a | 4.33 ± 0.22 a | 4.55 ± 0.26 a | 4.88 ± 0.27 a |
| Model | 3.29 ± 0.08 a | 3.33 ± 0.16 a | 3.59 ± 0.12 a | 3.89 ± 0.18 a | 4.15 ± 0.22 a | 4.36 ± 0.24 a | 4.61 ± 0.28 a | 4.92 ± 0.23 a |
| LF-CQPC05 | 3.22 ± 0.10 a | 3.35 ± 0.15 a | 3.63 ± 0.17 a | 3.95 ± 0.15 a | 4.21 ± 0.16 a | 3.29 ± 0.23 a | 4.59 ± 0.18 a | 4.93 ± 0.25 a |
| l-carnitine | 3.26 ± 0.12 a | 3.32 ± 0.14 a | 3.66 ± 0.16 a | 3.93 ± 0.16 a | 4.23 ± 0.23 a | 3.35 ± 0.25 a | 4.66 ± 0.19 a | 5.02 ± 0.27 a |
| LDSB | 3.31 ± 0.14 a | 3.33 ± 0.17 a | 3.61 ± 0.19 a | 3.94 ± 0.16 a | 4.19 ± 0.20 a | 3.32 ± 0.24 a | 4.60 ± 0.25 a | 5.05 ± 0.22 a |
| Group | Hepatosomatic Index | Epididymal Adipose Index | Perirenal Adipose Index |
|---|---|---|---|
| Normal | 32.38 ± 0.71 e | 10.30 ± 0.53 e | 1.56 ± 0.17 e |
| Model | 48.32 ± 1.20 a | 28.16 ± 2.08 a | 12.13 ± 1.26 a |
| LF-CQPC05 | 35.36 ± 1.03 d | 14.25 ± 1.81 d | 3.41 ± 0.48 d |
| l-carnitine | 38.23 ± 1.12 c | 18.33 ± 1.65 c | 5.20 ± 0.62 c |
| LDSB | 42.39 ± 1.06 c | 23.11 ± 1.76 b | 8.39 ± 0.54 b |
| Group | ALT (U/L) | AST (U/L) | HDL-C (mmol/L) | LDL-C (mmol/L) | TC (mmol/L) | TG (mmol/L) |
|---|---|---|---|---|---|---|
| Normal | 17.08 ± 1.33 e | 11.02 ± 0.89 e | 1.08 ± 0.12 a | 0.48 ± 0.06 e | 1.62 ± 0.21 e | 0.47 ± 0.04 e |
| Model | 60.35 ± 3.87 a | 51.88 ± 2.13 a | 0.28 ± 0.06 e | 1.97 ± 0.32 a | 5.86 ± 0.52 a | 1.70 ± 0.11 a |
| LF-CQPC05 | 29.82 ± 1.86 d | 18.01 ± 1.12 d | 0.83 ± 0.06 b | 0.71 ± 0.08 d | 2.21 ± 0.24 d | 0.68 ± 0.08 d |
| l-carnitine | 35.17 ± 1.38 c | 25.62 ± 1.53 c | 0.62 ± 0.05 c | 0.93 ± 0.07 c | 3.03 ± 0.25 c | 0.93 ± 0.10 c |
| LDSB | 48.71 ± 2.02 b | 34.19 ± 1.66 b | 0.49 ± 0.05 d | 1.34 ± 0.16 b | 4.18 ± 0.22 b | 1.36 ± 0.12 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.; Tan, F.; Mu, J.; Yi, R.; Zhou, X.; Zhao, X. Anti-Obesity Effects of Lactobacillus fermentum CQPC05 Isolated from Sichuan Pickle in High-Fat Diet-Induced Obese Mice through PPAR-α Signaling Pathway. Microorganisms 2019, 7, 194. https://doi.org/10.3390/microorganisms7070194
Zhu K, Tan F, Mu J, Yi R, Zhou X, Zhao X. Anti-Obesity Effects of Lactobacillus fermentum CQPC05 Isolated from Sichuan Pickle in High-Fat Diet-Induced Obese Mice through PPAR-α Signaling Pathway. Microorganisms. 2019; 7(7):194. https://doi.org/10.3390/microorganisms7070194
Chicago/Turabian StyleZhu, Kai, Fang Tan, Jianfei Mu, Ruokun Yi, Xianrong Zhou, and Xin Zhao. 2019. "Anti-Obesity Effects of Lactobacillus fermentum CQPC05 Isolated from Sichuan Pickle in High-Fat Diet-Induced Obese Mice through PPAR-α Signaling Pathway" Microorganisms 7, no. 7: 194. https://doi.org/10.3390/microorganisms7070194
APA StyleZhu, K., Tan, F., Mu, J., Yi, R., Zhou, X., & Zhao, X. (2019). Anti-Obesity Effects of Lactobacillus fermentum CQPC05 Isolated from Sichuan Pickle in High-Fat Diet-Induced Obese Mice through PPAR-α Signaling Pathway. Microorganisms, 7(7), 194. https://doi.org/10.3390/microorganisms7070194