Terpenoid-Induced Feeding Deterrence and Antennal Response of Honey Bees

 , and
, and 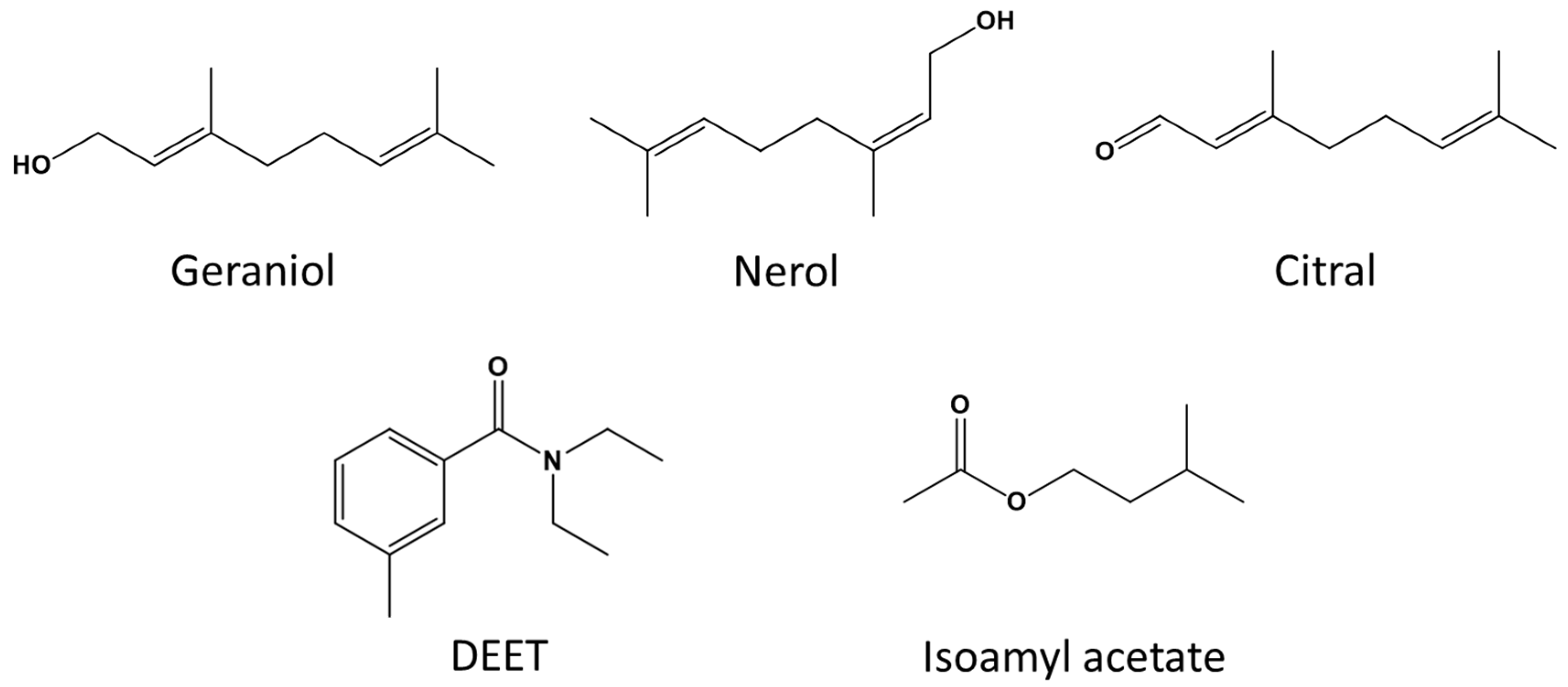{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Compounds
2.3. Video-Tracking Recordings
2.4. Electroantennogram (EAG) Recordings
3. Results
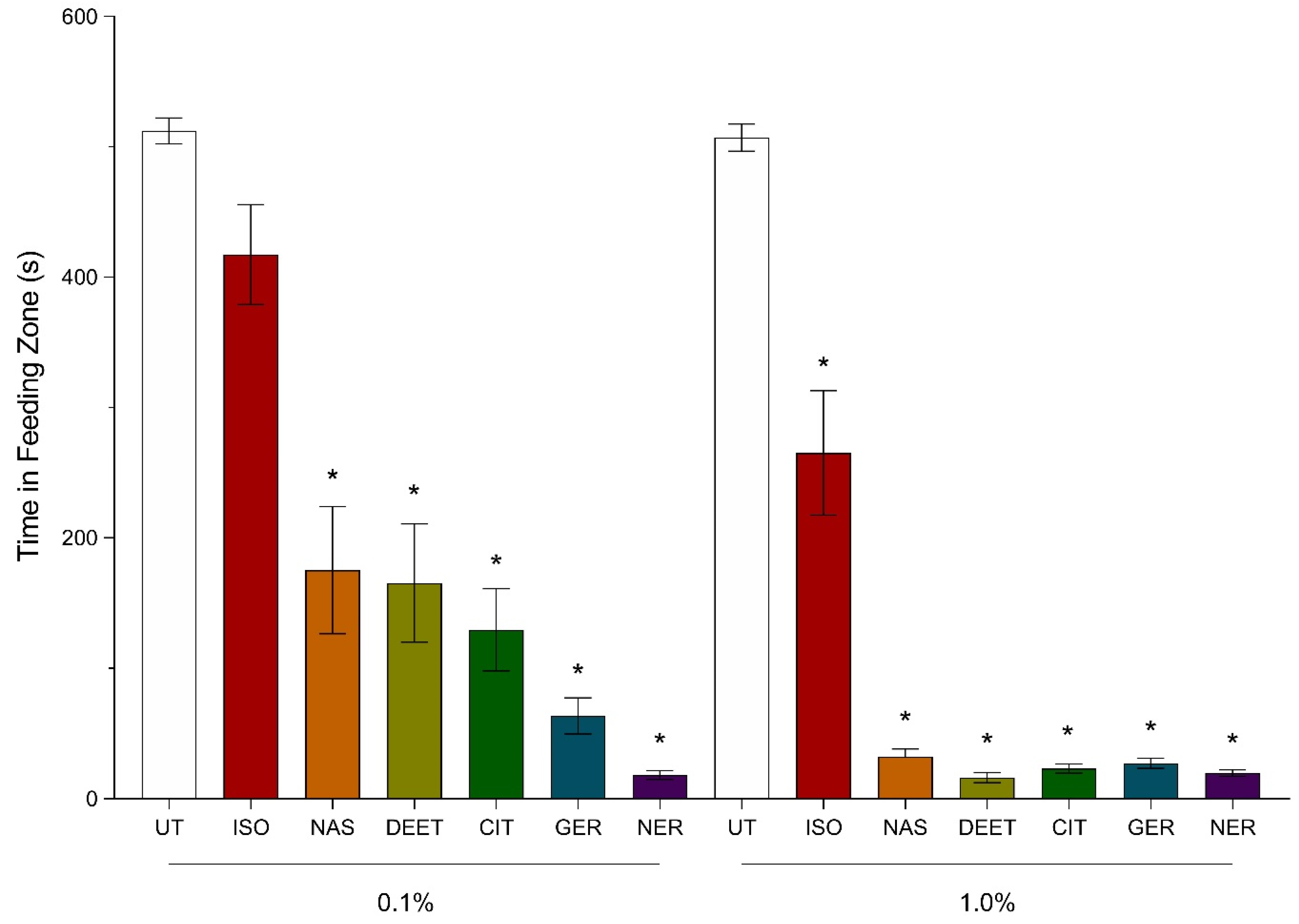
3.1. Video-Tracking Recordings
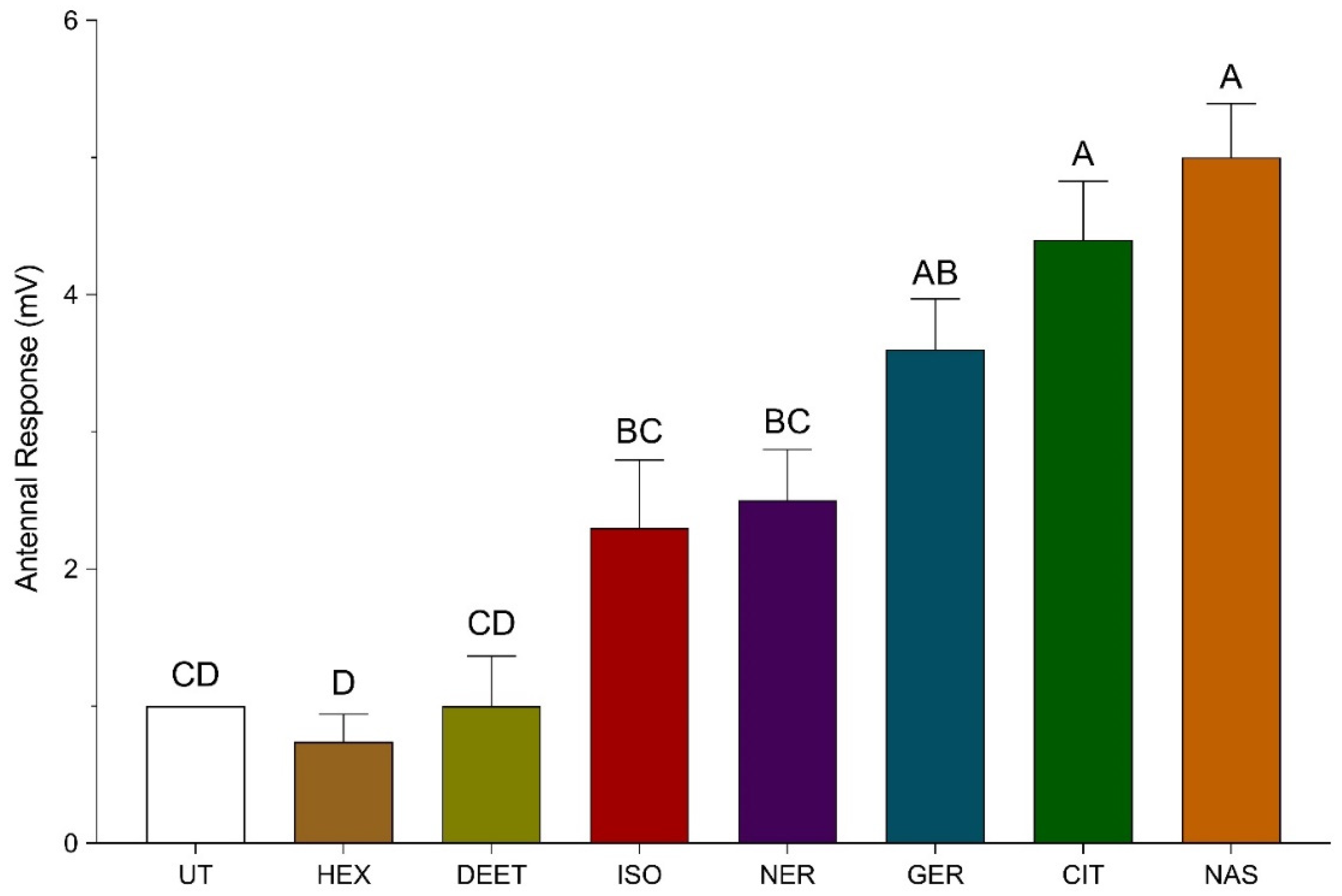
3.2. Electroantennogram (EAG) Recordings
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissiere, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/en/ (accessed on 1 May 2019).
- Van Engelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, S.T.; Anderson, T.D.; Wu-Smart, J.Y. Interactions between pesticides and pathogen susceptibility in honey bees. Curr. Opin. Insect Sci. 2018, 26, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, A.M.; O’Neal, S.T.; Fell, R.D.; Brewster, C.C.; Anderson, T.D. In-hive acaricides alter biochemical and morphological indicators of honey bee nutrition, immunity, and development. J. Insect Sci. 2018, 18, 8. [Google Scholar] [CrossRef]
- O’Neal, S.T.; Reeves, A.M.; Fell, R.D.; Brewster, C.C.; Anderson, T.D. Chlorothalonil exposure alters virus susceptibility and markers of immunity, nutrition, and development in honey bees. J. Insect Sci. 2019, 19. [Google Scholar] [CrossRef]
- Tison, L.; Hahn, M.-L.; Holtz, S.; Rößner, A.; Greggers, U.; Bischoff, G.; Menzel, R. Honey bees’ behavior is impaired by chronic exposure to the neonicotinoid thiacloprid in the field. Environ. Sci. Technol. 2016, 50, 7218–7227. [Google Scholar] [CrossRef]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honey bees (Apis mellifera L.). J. Insect Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef]
- Brandt, A.; Grikscheit, K.; Siede, R.; Grosse, R.; Meixner, M.D.; Büchler, R. Immunosuppression in honeybee queens by the neonicotinoids thiacloprid and clothianidin. Sci. Rep. UK 2017, 7, 4673. [Google Scholar] [CrossRef] [Green Version]
- Siede, R.; Meixner, M.D.; Almanza, M.T.; Schöning, R.; Maus, C.; Büchler, R. A long-term field study on the effects of dietary exposure of clothianidin to varroosis-weakened honey bee colonies. Ecotoxicology 2018, 27, 772–783. [Google Scholar] [CrossRef] [Green Version]
- Colin, T.; Meikle, W.G.; Paten, A.M.; Barron, A.B. Long-term dynamics of honey bee colonies following exposure to chemical stress. Sci. Total Environ. 2019, 677, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.A.D.; Mulhauser, B.; Mulot, M.; Mutabazi, A.; Glauser, G.; Aebi, A. A worldwide survey of neonicotinoids in honey. Science 2017, 358, 109–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostiguy, N.; Drummond, F.A.; Aronstein, K.; Eitzer, B.; Ellis, J.D.; Spivak, M.; Sheppard, W.S. Honey bee exposure to pesticides: A four-year nationwide study. Insects 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Jones, G.D.G. The responses of the honey-bee to repellent chemicals. J. Exp. Biol. 1952, 29, 372–386. [Google Scholar]
- Woodrow, A.W.; Green, N.; Tucker, H.; Schonhorst, M.H.; Hamilton, K.C. Evaluation of chemicals as honey bee attractants and repellents. J. Econ. Entomol. 1965, 58, 1094–1102. [Google Scholar] [CrossRef]
- Atkins, E.L.; Macdonald, R.L.; Greywood-Hale, E.A. Repellent additives to reduce pesticide hazards to honey bees: Field tests. Environ. Entomol. 1975, 4, 207–210. [Google Scholar] [CrossRef]
- Katz, T.M.; Miller, J.H.; Hebert, A.A. Insect repellents: Historical perspectives and new developments. J. Am. Acad. Dermatol. 2008, 58, 865–871. [Google Scholar] [CrossRef]
- Free, J.B.; Pickett, J.A.; Ferguson, A.W.; Simpkins, J.R.; Smith, M.C. Repelling foraging honeybees with alarm pheromones. J. Agric. Sci. 1985, 105, 255–260. [Google Scholar] [CrossRef]
- Vallet, A.; Cassier, P.; Lensky, Y. Ontogeny of the fine structure of the mandibular glands of the honeybee (Apis mellifera L.) workers and the pheromonal activity of 2-heptanone. J. Insect Physiol. 1991, 37, 789–804. [Google Scholar] [CrossRef]
- Jay, S.C. Spatial Management of Honey-Bees on Crops. Annu. Rev. Entomol. 1986, 31, 49–65. [Google Scholar] [CrossRef]
- Larson, N.R.; Anderson, T.D. Video tracking protocol to screen deterrent chemistries for honey bees. JoVE 2017, e55603. [Google Scholar] [CrossRef] [PubMed]
- Pickett, J.A.; Williams, I.H.; Martin, A.P.; Smith, M.C. Nasonov pheromone of the honey bee, Apis mellifera L. (Hymenoptera: Apidae). 1. Chemical characterization. J. Chem. Ecol. 1980, 6, 425–434. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A. Identification of nerolic and geranic acids in the Nassanoff pheromone of the honey bee. Nature 1964, 202, 320–321. [Google Scholar] [CrossRef]
- Shearer, D.A.; Boch, R. Citral in the Nassanoff pheromone of the honey bee. J. Insect Physiol. 1966, 12, 1513–1521. [Google Scholar] [CrossRef]
- Tangtrakulwanich, K.; Chen, H.; Baxendale, F.; Brewer, G.; Zhu, J.J. Characterization of olfactory sensilla of Stomoxys calcitrans and electrophysiological responses to odorant compounds associated with hosts and oviposition media. Med. Vet. Entomol. 2011, 25, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.O. Attractant or pheromone: The case of Nasonov secretion and honeybee swarms. J. Chem. Ecol. 1999, 25, 2051–2056. [Google Scholar] [CrossRef]
- Schmidt, J.O.; Hurley, R. Selection of nest cavities by Africanized and European honey bees. Apidologie 1995, 26, 467–475. [Google Scholar] [CrossRef]
- Free, J.B.; Ferguson, A.W.; Pickett, J.A.; Williams, I.H. Use of unpurified Nasonov pheromone components to attract clustering honeybees. J. Apic. Res. 1982, 21, 26–29. [Google Scholar] [CrossRef]
- Free, J.B.; Ferguson, A.W.; Pickett, J.A. Evaluation of the various components of the Nasonov pheromone used by clustering honeybees. Physiol. Entomol. 1981, 6, 263–268. [Google Scholar] [CrossRef]
- Williams, I.H.; Pickett, J.A.; Martin, A.P. The Nasonov pheromone of the honeybee Apis mellifera L. (Hymenoptera, Apidae). Part II. Bioassay of the components using foragers. J. Chem. Ecol. 1981, 7, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects—Finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambolena, J.S.; Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Areco, V.A.; Zygadlo, J.A. Terpenes: Natural products for controlling insects of importance to human health—A structure-activity relationship study. Psyche 2016, 2016, 17. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, H.; Ritter, W. The influence of the Nasonov pheromone on the recognition of house bees and foragers by Varroa jacobsoni. Apidologie 1988, 19, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Price, D.N.; Berry, M.S. Neurophysiological effects of naturally occurring defensive compounds on the freshwater snail Planorbis corneus: Comparison with effects in insects. J. Chem. Ecol. 2008, 34, 994. [Google Scholar] [CrossRef] [PubMed]
- Barnard, D.R.; Xue, R.-D. Laboratory evaluation of mosquito repellents against Aedes albopictus, Culex nigripalpus, and Ochlerotatus triseriatus (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 726–730. [Google Scholar] [CrossRef] [Green Version]
- Müller, G.C.; Junnila, A.; Kravchenko, V.D.; Revay, E.E.; Butler, J.; Schlein, Y. Indoor protection against mosquito and sand fly bites: A comparison between citronella, linalool, and geraniol candles. J. Am. Mosq. Control Assoc. 2008, 24, 150–153, 154. [Google Scholar]
- Wirtz, R.A.; Turrentine, J.D., Jr.; Rutledge, L.C. Mosquito area repellents: Laboratory testing of candidate materials against Aedes aegypti. Mosq. News 1980, 40, 432–439. [Google Scholar]
- Chadha, M.S.; Eisner, T.; Monro, A.; Meinwald, J. Defence Mechanisms of Arthropods .7. Citronellal and Citral in the Mandibular Gland Secretion of the Ant Acanthomyops-Claviger (Roger). J. Insect Physiol. 1962, 8, 175. [Google Scholar] [CrossRef]
- Butler, C.G.; Calam, D.H. Pheromones of the honey bee—The secretion of the Nassanoff gland of the worker. J. Insect Physiol. 1969, 15, 237–244. [Google Scholar] [CrossRef]
- Johnson, L.K.; Wiemer, D.F. Nerol: An alarm substance of the stingless bee, Trigona fulviventris (Hymenoptera: Apidae). J. Chem. Ecol. 1982, 8, 1167–1181. [Google Scholar] [CrossRef] [PubMed]
- Lwande, W.; Ndakala, A.J.; Hassanali, A.; Moreka, L.; Nyandat, E.; Ndungu, M.; Amiani, H.; Gitu, P.M.; Malonza, M.M.; Punyua, D.K. Gynandropsis gynandra essential oil and its constituents as tick (Rhipicephalus appendiculatus) repellents. Phytochemistry 1999, 50, 401–405. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, H.-P.; Wei, J.-Y.; Yang, K.; Tian, Z.-F.; Li, X.-L.; Wang, P.-J.; Wang, C.-F.; Du, S.-S.; Cai, Q. Repellent constituents of essential oil from Citrus wilsonii stem barks against Tribolium castaneum. Nat. Prod. Commun. 2014, 9. [Google Scholar] [CrossRef]
- Naik, D.G.; Vaidya-Kannur, H.; Deshpande, P.V.; Dandge, C.N.; Reddy, G.V.P. Potential use of an essential oil from the flower of Swertia densifolia as a repellent for Apis florea (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 2015, 108, 18–25. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A.; Stone, B.C. Identification of iso-amyl acetate as an active component in the sting pheromone of the honey bee. Nature 1962, 195, 1018–1020. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A.; Petrasovits, A. Efficacies of two alarm substances of the honey bee. J. Insect Physiol. 1970, 16, 17–24. [Google Scholar] [CrossRef]
- Free, J.B. The stimuli releasing the stinging response of honeybees. Anim. Behav. 1961, 9, 193–196. [Google Scholar] [CrossRef]
- Ghent, R.L.; Gary, N.E. A chemical alarm releaser in honey bee stings (Apis Mellifera L.). Psyche 1962, 69, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Free, J.B.; Simpson, J. The alerting pheromones of the honeybee. Z. Für Vgl. Physiol. 1968, 61, 361–365. [Google Scholar] [CrossRef]
- Williams, I.H.; Pickett, J.A.; Martin, A.P. Nasonov pheromone of the honeybee Apis mellifera L. (Hymenoptera, Apidae): IV. Comparative electroantennogram responses. J. Chem. Ecol. 1982, 8, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Akers, R.P.; Getz, W.M. A test of identified response classes among olfactory receptor neurons in the honey-bee worker. Chem. Senses 1992, 17, 191–209. [Google Scholar] [CrossRef]
- Akers, R.P.; Getz, W.M. Response of olfactory receptor neurons in honeybees to odorants and their binary mixtures. J. Comp. Physiol. A 1993, 173, 169–185. [Google Scholar] [CrossRef]
- Simpson, R.F. Bioassay of pine oil components as attractants for Sirex noctilio (Hymenoptera: Siricidae) using electroantennogram techniques. Entomol. Exp. Appl. 1976, 19, 11–18. [Google Scholar] [CrossRef]
- Fonta, C.; Masson, C. Comparative study by electrophysiology of olfactory responses in bumblebees (Bombus hypnorum and Bombus terrestris). J. Chem. Ecol. 1984, 10, 1157–1168. [Google Scholar] [CrossRef]
- Masson, C.; Arnold, G. Ontogeny, maturation and plasticity of the olfactory system in the workerbee. J. Insect Physiol. 1984, 30, 7–14. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larson, N.R.; O’Neal, S.T.; Bernier, U.R.; Bloomquist, J.R.; Anderson, T.D. Terpenoid-Induced Feeding Deterrence and Antennal Response of Honey Bees. Insects 2020, 11, 83. https://doi.org/10.3390/insects11020083
Larson NR, O’Neal ST, Bernier UR, Bloomquist JR, Anderson TD. Terpenoid-Induced Feeding Deterrence and Antennal Response of Honey Bees. Insects. 2020; 11(2):83. https://doi.org/10.3390/insects11020083
Chicago/Turabian StyleLarson, Nicholas R., Scott T. O’Neal, Ulrich R. Bernier, Jeffrey R. Bloomquist, and Troy D. Anderson. 2020. "Terpenoid-Induced Feeding Deterrence and Antennal Response of Honey Bees" Insects 11, no. 2: 83. https://doi.org/10.3390/insects11020083
APA StyleLarson, N. R., O’Neal, S. T., Bernier, U. R., Bloomquist, J. R., & Anderson, T. D. (2020). Terpenoid-Induced Feeding Deterrence and Antennal Response of Honey Bees. Insects, 11(2), 83. https://doi.org/10.3390/insects11020083