Streptococcus mutans and Actinomyces naeslundii Interaction in Dual-Species Biofilm

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of Single- and Dual-Species Biofilms
2.2. Stress Response of Single and Dual-Species Biofilms Using Drip-Flow Reactor
2.3. Data Analysis
3. Results
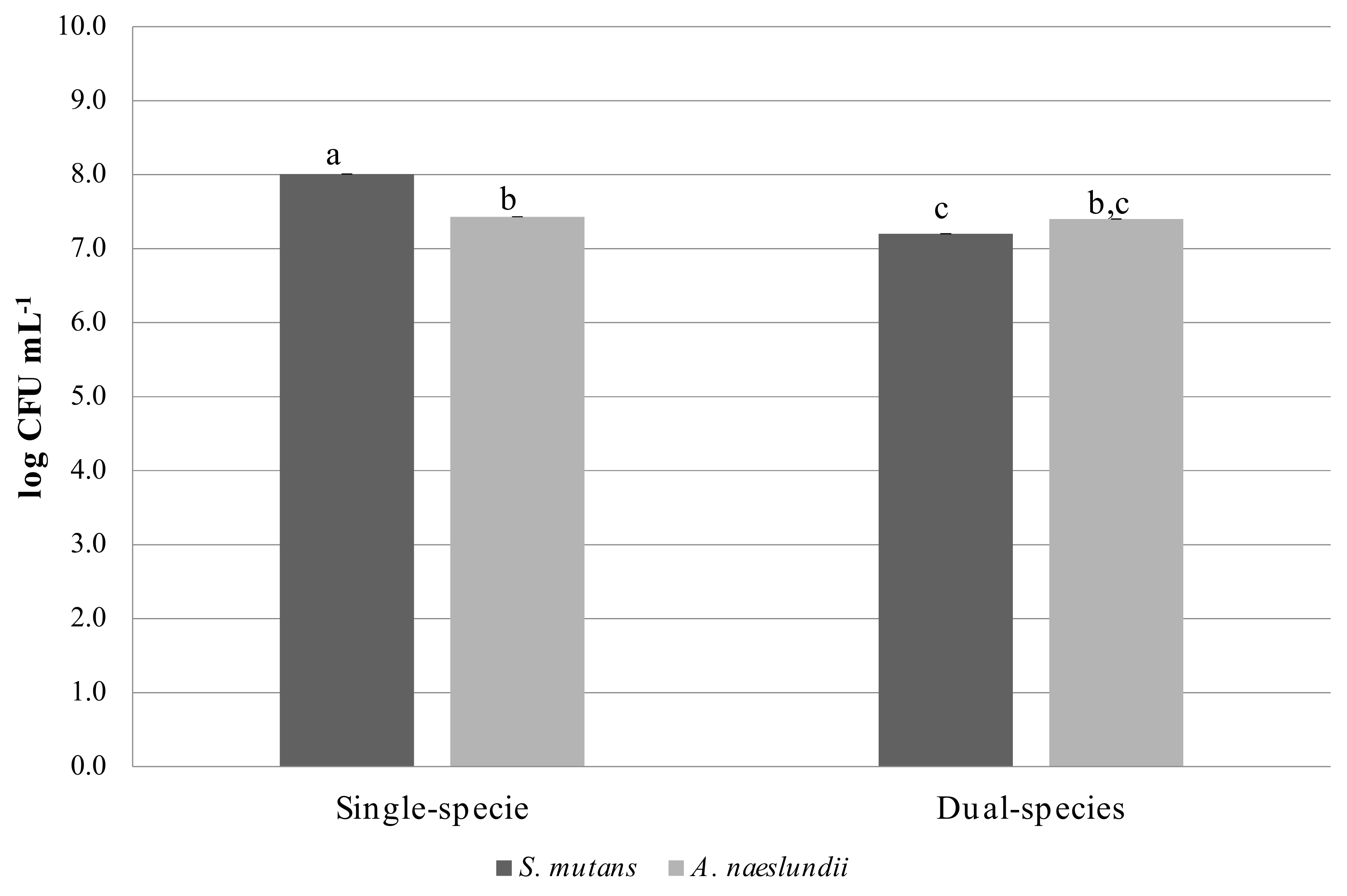
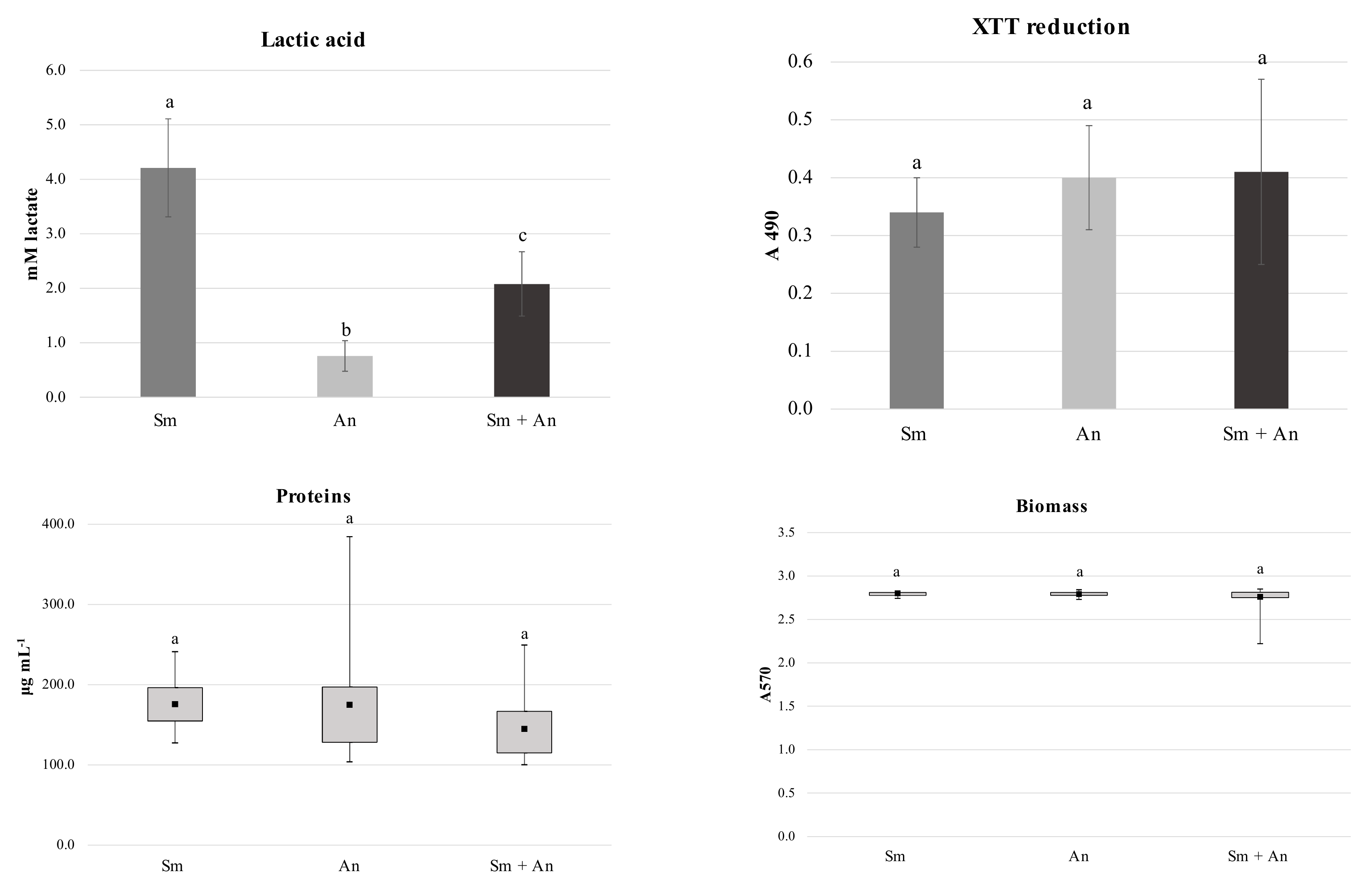
3.1. Characterization of Single and Dual Species Biofilms
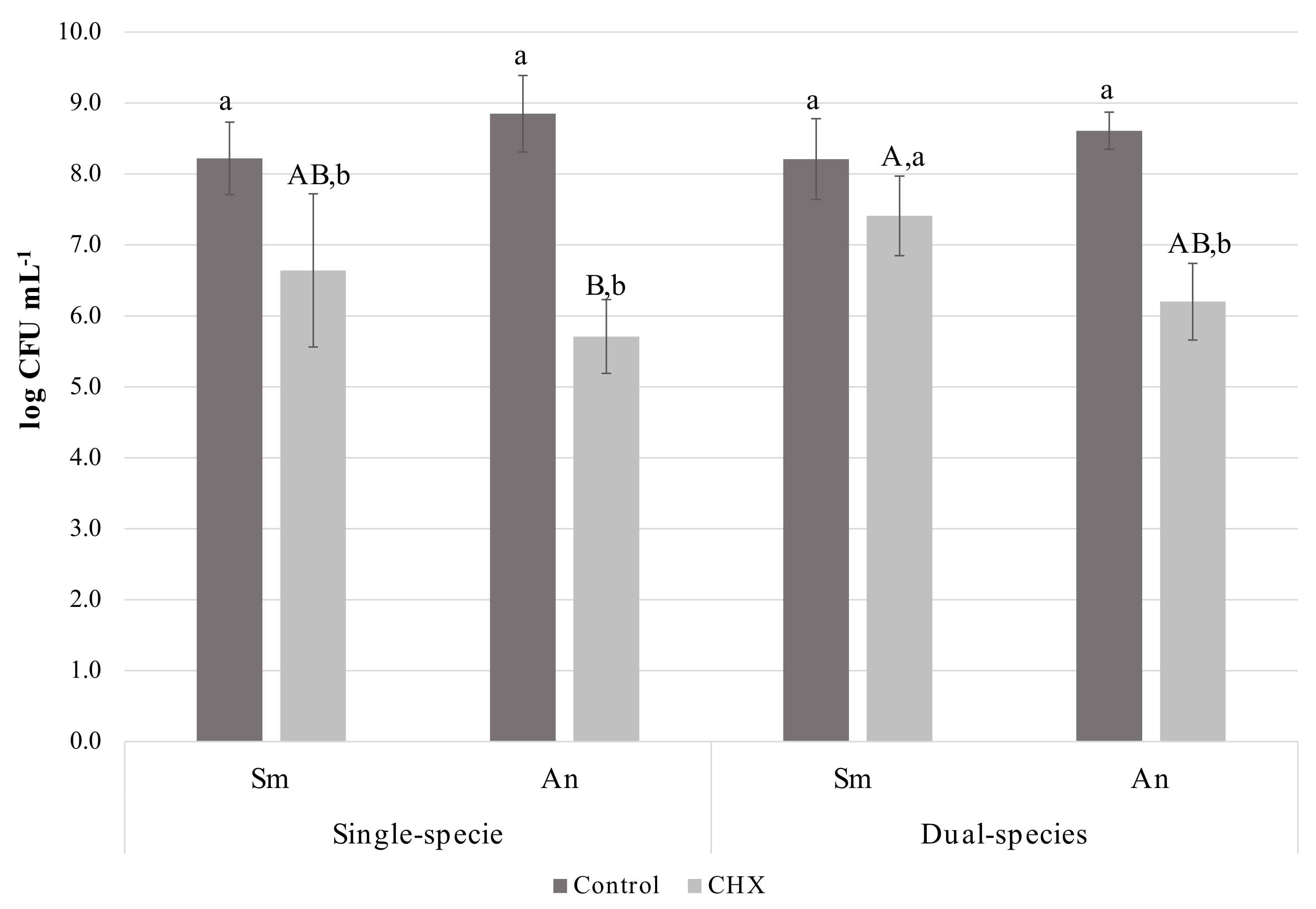
3.2. Stress Response of Single- and Dual-Species Biofilms Using Drip-Flow Reactor
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loesche, W.J. Chemotherapy of dental plaque infections. Oral Sci. Rev. 1976, 9, 65–107. [Google Scholar]
- Kolenbrander, P.E.; Palmer, R.J.; Rickard, A.H.; Jakubovics, N.S.; Chalmers, N.I.; Diaz, P.I. Bacterial interactions and successions during plaque development. Periodontol 2000 2006, 42, 47–79. [Google Scholar] [CrossRef]
- Darveau, R.P.; Hajishengallis, G.; Curtis, M.A. Porphyromonas gingivalis as a potential community activist for disease. J. Dent. Res. 2012, 91, 816–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Kistler, J.O.; Pesaro, M.; Wade, W.G. Development and pyrosequencing analysis of an in-vitro oral biofilm model. BMC Microbiol. 2015, 15, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, P.D. Dental plaque: Biological significance of a biofilm and community life-style. J. Clin. Periodontol. 2005, 32, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E. Multispecies communities: Interspecies interactions influence growth on saliva as sole nutritional source. Int. J. Oral Sci. 2011, 3, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial coaggregation: An integral process in the development of multi-species biofilms. Trends Microbiol. 2003, 11, 94–100. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. Caries ecology revisited: Microbial dynamics and the caries process. Caries Res. 2008, 42, 409–418. [Google Scholar] [CrossRef]
- Bradshaw, D.J.; Marsh, P.D.; Watson, G.K.; Allison, C. Oral anaerobes cannot survive oxygen stress without interacting with facultative/aerobic species as a microbial community. Lett. Appl. Microbiol. 1997, 25, 385–387. [Google Scholar] [CrossRef]
- Jakubovics, N.S.; Gill, S.R.; Iobst, S.E.; Vickerman, M.M.; Kolenbrander, P.E. Regulation of gene expression in a mixed-genus community: Stabilized arginine biosynthesis in Streptococcus gordonii by coaggregation with Actinomyces naeslundii. J. Bacteriol. 2008, 190, 3646–3657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Kim, Y.J. A comparative study of the effect of probiotics on cariogenic biofilm model for preventing dental caries. Arch. Microbiol. 2014, 196, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E.; Andersen, R.N.; Moore, L.V.H. Coaggregation of Fusobacterium nucleatum, Selenomonas flueggei, Selenomonas infelix, Selenomonas noxia, and Selenomonas sputigena with strains from 11 genera of oral bacteria. Infect. Immun. 1989, 57, 3194–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreth, J.; Merritt, J.; Shi, W.; Qi, F. Competition and coexistence between Streptococcus mutans and Streptococcus sanguinis in the dental biofilm. J. Bacteriol. 2005, 187, 7193–7203. [Google Scholar] [CrossRef] [Green Version]
- Iwami, Y.; Yamada, T. Rate-limiting steps of the glycolytic pathway in the oral bacteria Streptococcus mutans and Streptococcus sanguis and the influence of acidic pH on the glucose metabolism. Arch. Oral Biol. 1980, 25, 163–169. [Google Scholar] [CrossRef]
- Ahmed, A.; Dachang, W.; Lei, Z.; Jianjun, L.; Juanjuan, Q.; Yi, X. Effect of Lactobacillus species on Streptococcus mutans biofilm formation. Pak. J. Pharm. Sci. 2014, 27, 1523–1528. [Google Scholar]
- Wen, Z.T.; Yates, D.; Ahn, S.J.; Burne, R.A. Biofilm formation and virulence expression by Streptococcus mutans are altered when grown in dual-species model. BMC Microbiol. 2010, 10, 111. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, A.; Ansai, T.; Takehara, T.; Kuramitsu, H.K. LuxS-based signaling affects Streptococcus mutans biofilm formation. Appl. Environ. Microbiol. 2005, 71, 2372–2380. [Google Scholar] [CrossRef] [Green Version]
- Dame-Teixeira, N.; Parolo, C.C.F.; Maltz, M.; Tugnait, A.; Devine, D.; Do, T. Actinomyces spp. gene expression in root caries lesions. J. Oral Microbiol. 2016, 8, 32383. [Google Scholar] [CrossRef] [Green Version]
- Garcia, S.S.; Du, Q.; Wu, H. Streptococcus mutans copper chaperone, CopZ, is critical for biofilm formation and competitiveness. Mol. Oral Microbiol. 2016, 31, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Kawada-Matsuo, M.; Oogai, Y.; Komatsuzawa, H. Sugar allocation to metabolic pathways is tightly regulated and affects the virulence of Streptococcus mutans. Genes 2017, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, W.; Ren, B.; Dai, H.; Zhang, L.; Zhang, Q.; Zhou, X.; Li, Y. Clotrimazole and econazole inhibit Streptococcus mutans biofilm and virulence in vitro. Arch. Oral Biol. 2017, 73, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Brighenti, F.L.; Luppens, S.B.I.; Delbem, A.C.B.; Deng, D.M.; Hoogenkamp, M.A.; Gaetti-Jardim Jr., E.; Dekker, H.L.; Crielaard, W.; ten Cate, J.M. Effect of psidium cattleianum leaf extract on Streptococcus mutans viability, protein expression and acid production. Caries Res. 2008, 42, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Mattos-Graner, R.O.; Klein, M.I.; Smith, D.J. Lessons Learned from Clinical Studies: Roles of Mutans Streptococci in the Pathogenesis of Dental Caries. Curr. Oral Health Rep. 2014, 1, 70. [Google Scholar] [CrossRef]
- Kara, D.; Luppens, S.B.I.; ten Cate, J.M. Differences between single- and dual-species biofilms of Streptococcus mutans and Veillonella parvula in growth, acidogenicity and susceptibility to chlorhexidine. Eur. J. Oral Sci. 2006, 114, 58–63. [Google Scholar] [CrossRef]
- Kara, D.; Luppens, S.B.I.; Van Marle, J.; Özok, R.; ten Cate, J.M. Microstructural differences between single-species and dual-species biofilms of Streptococcus mutans and Veillonella parvula, before and after exposure to chlorhexidine. FEMS Microbiol. Lett. 2007, 271, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luppens, S.B.I.; Kara, D.; Bandounas, L.; Jonker, M.J.; Wittink, F.R.A.; Bruning, O.; Breit, T.M.; ten Cate, J.M.; Crielaard, W. Effect of Veillonella parvula on the antimicrobial resistance and gene expression of Streptococcus mutans grown in a dual-species biofilm. Oral Microbiol. Immunol. 2008, 23, 183–189. [Google Scholar] [CrossRef]
- De Matos, B.M.; Brighenti, F.L.; Do, T.; Beighton, D.; Koga-Ito, C.Y. Acidogenicity of dual-species biofilms of bifidobacteria and Streptococcus mutans. Clin. Oral Investig. 2017, 21, 1769–1776. [Google Scholar] [CrossRef]
- Lobo, C.I.V.; Rinaldi, T.B.; Christiano, C.M.S.; de Sales Leite, L.; Barbugli, P.A.; Klein, M.I. Dual-species biofilms of Streptococcus mutans and Candida albicans exhibit more biomass and are mutually beneficial compared with single-species biofilms. J. Oral Microbiol. 2019, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, F.L.; Salvador, M.J.; Delbem, A.C.B.; Delbem, Á.C.B.; Oliveira, M.A.C.; Soares, C.P.; Freitas, L.S.F.; Koga-Ito, C.Y. Systematic screening of plant extracts from the brazilian pantanal with antimicrobial activity against bacteria with cariogenic relevance. Caries Res. 2014, 48, 353–360. [Google Scholar] [CrossRef]
- Liu, Y.L.; Nascimento, M.; Burne, R.A. Progress toward understanding the contribution of alkali generation in dental biofilms to inhibition of dental caries. Int. J. Oral Sci. 2012, 4, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, M.M.; Gordan, V.V.; Garvan, C.W.; Browngardt, C.M.; Burne, R.A. Correlations of oral bacterial arginine and urea catabolism with caries experience. Oral Microbiol. Immunol. 2009, 24, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N.; Yamada, T. Glucose and lactate metabolism by Actinomyces naeslundii. Crit. Rev. Oral Biol. Med. 1999, 10, 487–503. [Google Scholar] [CrossRef] [PubMed]
- Alves, F.R.F.; Silva, M.G.; Rôças, I.N.; Siqueira, J.F. Biofilm biomass disruption by natural substances with potential for endodontic use. Braz. Oral Res. 2013, 27, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Filoche, S.K.; Soma, K.J.; Sissons, C.H. Caries-related plaque microcosm biofilms developed in microplates. Oral Microbiol. Immunol. 2007, 22, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Paes Leme, A.F.; Bellato, C.M.; Bedi, G.; Cury, A.A.D.B.; Koo, H.; Cury, J.A. Effects of sucrose on the extracellular matrix of plaque-like biofilm formed in vivo, studied by proteomic analysis. Caries Res. 2008, 42, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Paes Leme, A.F.; Koo, H.; Bellato, C.M.; Bedi, G.; Cury, J.A. The role of sucrose in cariogenic dental biofilm formation--new insight. J. Dent. Res. 2006, 85, 878–887. [Google Scholar] [CrossRef]
- Moi, G.P.; Cury, J.A.; Dombroski, T.C.D.; Pauletti, B.A.; Paes Leme, A.F. Proteomic analysis of matrix of dental biofilm formed under dietary carbohydrate exposure. Caries Res. 2012, 46, 339–345. [Google Scholar] [CrossRef]
- Bowden, G.H.; Li, Y.H. Nutritional influences on biofilm development. Adv. Dent. Res. 1997, 11, 81–99. [Google Scholar] [CrossRef]
- Arias, L.S.; Delbem, A.C.B.; Fernandes, R.A.; Barbosa, D.B.; Monteiro, D.R. Activity of tyrosol against single and mixed-species oral biofilms. J. Appl. Microbiol. 2016, 120, 1240–1249. [Google Scholar] [CrossRef] [Green Version]
- Exterkate, R.A.M.; Crielaard, W.; ten Cate, J.M. Different response to amine fluoride by Streptococcus mutans and polymicrobial biofilms in a novel high-throughput active attachment model. Caries Res. 2010, 44, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, Y.E.; Danelon, M.; Salvador, M.J.; Koga-Ito, C.Y.; Botazzo Delbem, A.C.; Ramirez-Rueda, R.Y.; Lacerda Gontijo, A.V.; Brighenti, F.L. Mouthwash containing Croton doctoris essential oil: In vitro study using a validated model of caries induction. Future Microbiol. 2018, 13, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Klein, M.I.; Falsetta, M.L.; Lu, B.; Delahunty, C.M.; Yates, J.R.; Heydorn, A.; Koo, H. The exopolysaccharide matrix modulates the interaction between 3D architecture and virulence of a mixed-species oral biofilm. PLoS Pathog. 2012, 8, e1002623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans and Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Merritt, J.H.; Kadouri, D.E.; O’toole, G.A. Growing and Analyzing Static Biofilms HHS Public Access. Curr. Protoc. Microbiol. 2005. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, F.L.; Medeiros, A.C.; Matos, B.M.; Ribeiro, Z.E.A.; Koga-Ito, C.Y. Evaluation of caries-associated virulence of biofilms from Candida albicans isolated from saliva of pediatric patients with sickle-cell anemia. J. Appl. Oral Sci. 2014, 22, 484–489. [Google Scholar] [CrossRef]
- Gutmann, I.; Wahlefeld, W. L-(+)-Lactate. Determination with lactate dehydrogenase and NAD. Methods of Enzym. Anal. 1974, 3, 1464–1472. [Google Scholar]
- Cheng, X.; Liu, J.; Li, J.; Zhou, X.; Wang, L.; Liu, J.; Xu, X. Comparative effect of a stannous fluoride toothpaste and a sodium fluoride toothpaste on a multispecies biofilm. Arch. Oral Biol. 2017, 74, 5–11. [Google Scholar] [CrossRef]
- Duckworth, R.M.; Jones, S. On the relationship between the rate of salivary flow and salivary fluoride clearance. Caries Res. 2015, 49, 141–146. [Google Scholar] [CrossRef]
- Cousido, M.C.; Carmona, I.T.; García-Caballero, L.; Limeres, J.; Álvarez, M.; Diz, P. In vivo substantivity of 0.12% and 0.2% chlorhexidine mouthrinses on salivary bacteria. Clin. Oral Investig. 2010, 14, 397–402. [Google Scholar] [CrossRef]
- Sánchez, M.C.; Llama-Palacios, A.; Blanc, V.; León, R.; Herrera, D.; Sanz, M. Structure, viability and bacterial kinetics of an in vitro biofilm model using six bacteria from the subgingival microbiota. J. Periodontal Res. 2011, 46, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Dige, I.; Raarup, M.K.; Nyengaard, J.R.; Kilian, M.; Nyvad, B. Actinomyces naeslundii in initial dental biofilm formation. Microbiology 2009, 155, 2116–2126. [Google Scholar] [CrossRef] [Green Version]
- Doel, J.J.; Benjamin, N.; Hector, M.P.; Rogers, M.; Allaker, R.P. Evaluation of bacterial nitrate reduction in the human oral cavity. Eur. J. Oral Sci. 2005, 113, 14–19. [Google Scholar] [CrossRef]
- Klein, M.I.; Hwang, G.; Santos, P.H.S.; Campanella, O.H.; Koo, H. Streptococcus mutans-derived extracellular matrix in cariogenic oral biofilms. Front. Cell. Infect. Microbiol. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Mothey, D.; Buttaro, B.A.; Piggot, P.J. Mucin can enhance growth, biofilm formation, and survival of Streptococcus mutans. FEMS Microbiol. Lett. 2014, 350, 161–167. [Google Scholar] [CrossRef]
- Bauer, J.; Siala, W.; Tulkens, P.M.; Van Bambeke, F. A combined pharmacodynamic quantitative and qualitative model reveals the potent activity of daptomycin and delafloxacin against Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 2013, 57, 2726–2737. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, M.E.; Schellmann, D.; Brunhofer, G.; Erker, T.; Busygin, I.; Leino, R.; Vuorela, P.M.; Fallarero, A. Pros and cons of using resazurin staining for quantification of viable Staphylococcus aureus biofilms in a screening assay. J. Microbiol. Methods 2009, 78, 104–106. [Google Scholar] [CrossRef]
- Welch, K.; Cai, Y.; Strømme, M. A Method for Quantitative Determination of Biofilm Viability. J. Funct. Biomater. 2012, 3, 418–431. [Google Scholar] [CrossRef] [Green Version]
- Alves, C.; Brandão, M.; Andion, J.; Menezes, R. Use of graduated syringes for measuring salivary flow rate: A pilot study. Braz. Dent. J. 2010, 21, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Blanc, V.; Isabal, S.; Sánchez, M.C.; Llama-Palacios, A.; Herrera, D.; Sanz, M.; León, R. Characterization and application of a flow system for in vitro multispecies oral biofilm formation. J. Periodontal Res. 2014, 49, 323–332. [Google Scholar] [CrossRef]
- Sawant, S.N.; Selvaraj, V.; Prabhawathi, V.; Doble, M. Antibiofilm Properties of Silver and Gold Incorporated PU, PCLm, PC and PMMA Nanocomposites under Two Shear Conditions. PLoS ONE 2013, 8, e63311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ASTM. International Standard Test Method for Quantification of Pseudomonas aeruginosa Biofilm Grown with High Shear and Continuous Flow using CDC Biofilm; ASTM E2562-17; ASTM Int.: Montgomery County, PA, USA, 2017. [Google Scholar]
- Brambilla, E.; Ionescu, A.; Cazzaniga, G.; Edefonti, V.; Gagliani, M. The influence of antibacterial toothpastes on in vitro Streptococcus mutans biofilm formation: A continuous culture study. Am. J. Dent. 2014, 27, 160–166. [Google Scholar] [PubMed]
- Hahnel, S.; Ionescu, A.C.; Cazzaniga, G.; Ottobelli, M.; Brambilla, E. Biofilm formation and release of fluoride from dental restorative materials in relation to their surface properties. J. Dent. 2017, 60, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, A.C.; Brambilla, E.; Travan, A.; Marsich, E.; Donati, I.; Gobbi, P.; Turco, G.; Di Lenarda, R.; Cadenaro, M.; Paoletti, S.; et al. Silver-polysaccharide antimicrobial nanocomposite coating for methacrylic surfaces reduces Streptococcus mutans biofilm formation in vitro. J. Dent. 2015, 43, 1483–1490. [Google Scholar] [CrossRef]
- Williams, D.L.; Epperson, R.T.; DeGrauw, J.P.; Nielsen, M.B.; Taylor, N.B.; Jolley, R.D. Effect of silver-loaded PMMA on Streptococcus mutans in a drip flow reactor. J. Biomed. Mater. Res. Part A 2017, 105, 2632–2639. [Google Scholar] [CrossRef]
- Guggenheim, B.; Meier, A. In vitro effect of chlorhexidine mouth rinses on polyspecies biofilms. Schweiz. Monatsschr. Zahnmed. 2011, 121, 432–441. [Google Scholar]
- García-Caballero, L.; Quintas, V.; Prada-López, I.; Seoane, J.; Donos, N.; Tomás, I. Chlorhexidine substantivity on salivary flora and plaque-like biofilm: An In situ model. PLoS ONE 2013, 8, e83522. [Google Scholar] [CrossRef] [Green Version]
- Arzmi, M.H.; Alnuaimi, A.D.; Dashper, S.; Cirillo, N.; Reynolds, E.C.; McCullough, M. Polymicrobial biofilm formation by Candida albicans, Actinomyces naeslundii, and Streptococcus mutans is Candida albicans strain and medium dependent. Med. Mycol. 2016, 54, 856–864. [Google Scholar] [CrossRef]
- Bermejo, P.; Sánchez, M.C.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Topographic characterization of multispecies biofilms growing on dental implant surfaces: An in vitro model. Clin. Oral Implants Res. 2019, 30, 229–241. [Google Scholar] [CrossRef]
- Kneist, S.; Kubieziel, H.; Kupper, H.; Willershausen, B.; Callaway, A. Modeling of S. mutans and A. naeslundii acid production in vitro with caries incidence of low- and high-risk children. Quintessence Int. 2012, 43, 413–420. [Google Scholar]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Filoche, S.K.; Zhu, M.; Wu, C.D. In situ biofilm formation by multi-species oral bacteria under flowing and anaerobic conditions. J. Dent. Res. 2004, 83, 802–806. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, R.V.D.; Bonafé, F.S.S.; Spolidorio, D.M.P.; Koga-Ito, C.Y.; de Farias, A.L.; Kirker, K.R.; James, G.A.; Brighenti, F.L. Streptococcus mutans and Actinomyces naeslundii Interaction in Dual-Species Biofilm. Microorganisms 2020, 8, 194. https://doi.org/10.3390/microorganisms8020194
de Oliveira RVD, Bonafé FSS, Spolidorio DMP, Koga-Ito CY, de Farias AL, Kirker KR, James GA, Brighenti FL. Streptococcus mutans and Actinomyces naeslundii Interaction in Dual-Species Biofilm. Microorganisms. 2020; 8(2):194. https://doi.org/10.3390/microorganisms8020194
Chicago/Turabian Stylede Oliveira, Rosa Virginia Dutra, Fernanda Salloume Sampaio Bonafé, Denise Madalena Palomari Spolidorio, Cristiane Yumi Koga-Ito, Aline Leite de Farias, Kelly R. Kirker, Garth A. James, and Fernanda Lourenção Brighenti. 2020. "Streptococcus mutans and Actinomyces naeslundii Interaction in Dual-Species Biofilm" Microorganisms 8, no. 2: 194. https://doi.org/10.3390/microorganisms8020194