Oligo-Carrageenan Kappa-Induced Reducing Redox Status and Increase in TRR/TRX Activities Promote Activation and Reprogramming of Terpenoid Metabolism in Eucalyptus Trees

Abstract
:1. Introduction

2. Results and Discussion
2.1. Main Terpenoids in the Leaves of Control Eucalyptus Trees
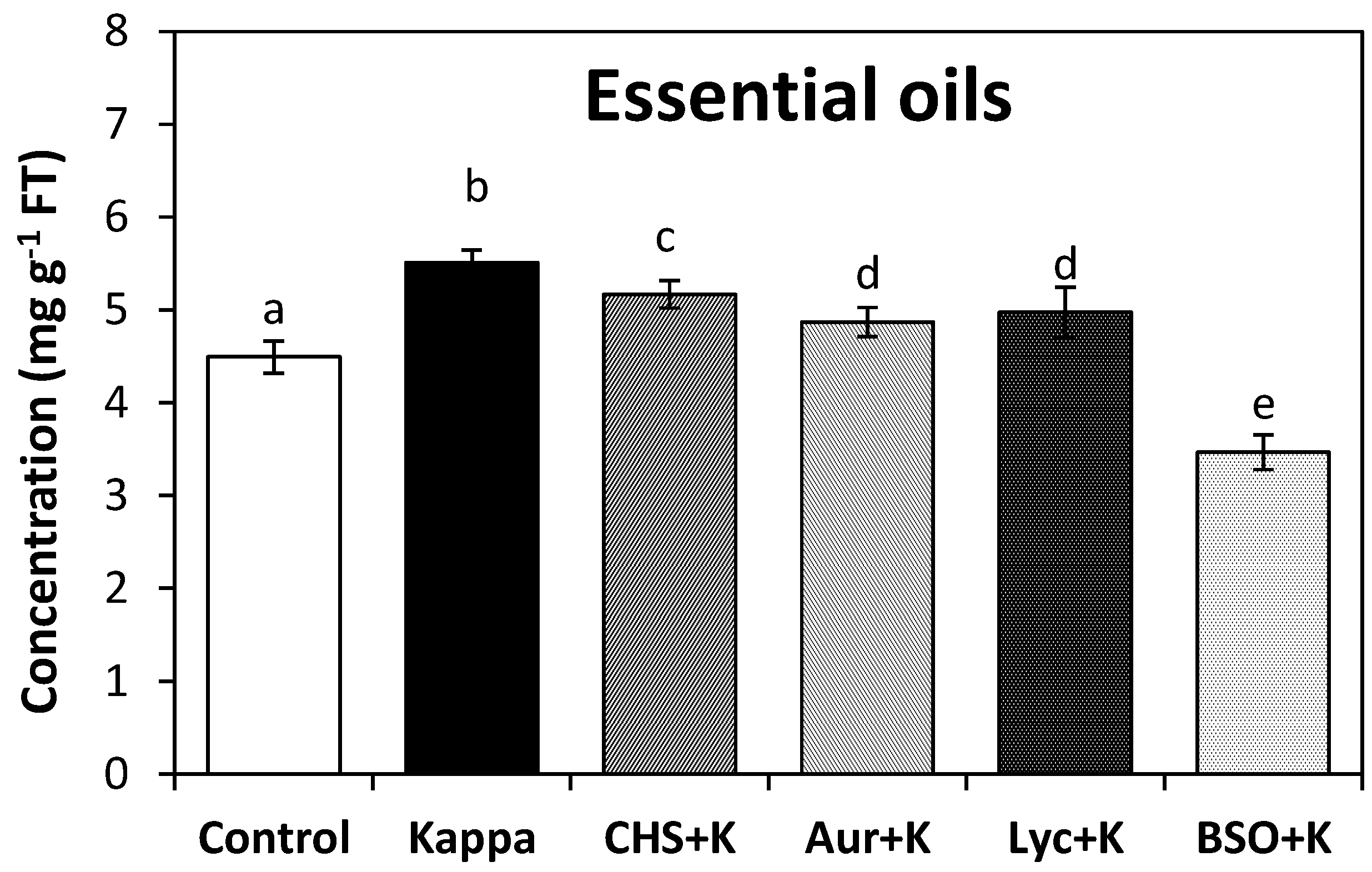
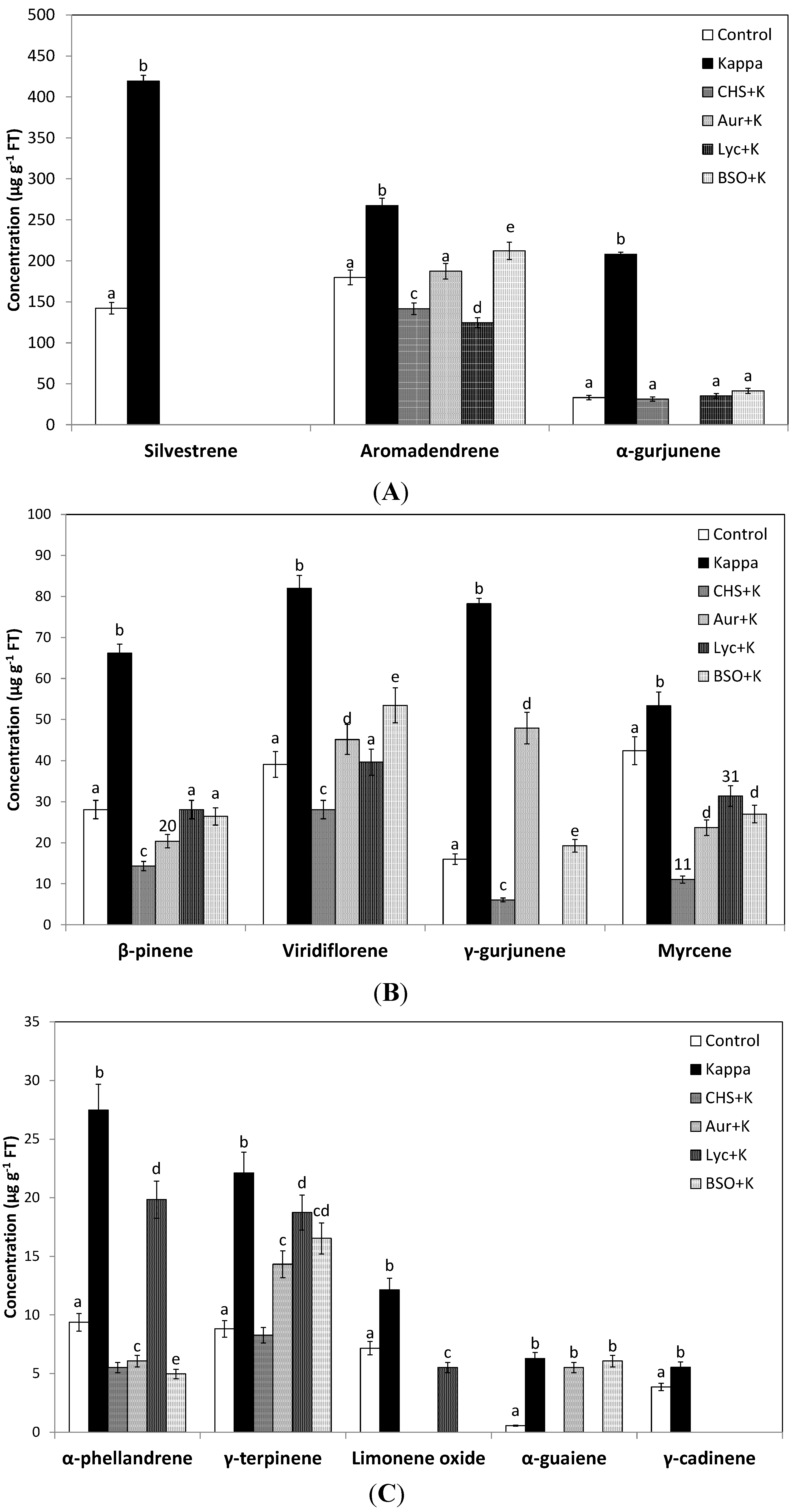
2.2. OC Kappa Induced an Increase in Total Essential Oils and a Reprogramming of Terpenoid Metabolism

{kind=link}
{kind=link}
{kind=link}
| Compound | Level in Control Trees (µg·g−1 FT) | Level in Treated Trees (µg·g−1 FT) | % of Decrease |
|---|---|---|---|
| Eucalyptol | 3757 ± 188 | 3485 ± 174 | 7.2 |
| Sabinene | 101 ± 8.1 | 42 ± 3.3 | 59 |
| α-Terpineol | 45 ± 3.6 | 39 ± 3.1 | 13 |
| α-Pinene | 499 ± 25 | 457 ± 23 | 8 |
| δ-Cadinene | 29 ± 2.3 | 14 ± 1.1 | 52 |
| Isoledene | 26 ± 2.1 | 11 ± 0.9 | 59 |
| Compound | Level in Control Trees (µg·g−1 FT) | Level in Treated Trees (µg·g−1 FT) | % of Increase |
|---|---|---|---|
| Silvestrene | 142 ± 7 | 419 ± 21 | 195 |
| α-Phellandrene | 9 ± 0.7 | 27 ± 2.2 | 193 |
| γ-Terpinene | 9 ± 0.7 | 22 ± 1.8 | 151 |
| Limonene oxide | 7 ± 0.6 | 12 ± 1 | 70 |
| β-Pinene | 28 ± 2.2 | 66 ± 5.3 | 135 |
| γ-Cadinene | 4 ± 0.3 | 5 ± 0.4 | 44 |
| Aromadendrene | 180 ± 9 | 267 ± 13.4 | 49 |
| Viridiflorene | 39 ± 3.1 | 82 ± 6.6 | 110 |
| α-Gurjunene | 33 ± 2.6 | 208 ± 17 | 529 |
| γ-Gurjunene | 16 ± 1.3 | 78 ± 6.3 | 390 |
| α-Guaiene | 1 ± 0.04 | 6 ± 0.5 | 1041 |
| Myrcene | 42 ± 3.4 | 53 ± 4.3 | 26 |
| Compound | Type | Level in Treated Trees (µg·g−1 FT) |
|---|---|---|
| Carene | Monoterpene | 4 ± 0.3 |
| α-Terpinene | Monoterpene | 3 ± 0.3 |
| α-Fenchene | Monoterpene | 8 ± 0.6 |
| γ-Maaliene | Sesquiterpene | 9 ± 0.8 |
| Spathulenol | Sesquiterpene | 5 ± 0.4 |
| α-Camphenolic aldehyde | Monoterpene | 6 ± 0.4 |
2.3. OC-Kappa Induced Reducing Redox Status Change the Level of Total Essential Oils and Induced Terpenoid Synthesis Reprogramming


3. Experimental
3.1. Preparation of OC Kappa
3.2. Treatment of Trees with OC Kappa and Inhibitors/OC Kappa
3.3. Distillation of Total Essential Oils
3.4. Analysis of Terpenoids by GC-MS
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aharoni, A.; Jongsma, M.A.; Kim, T.Y.; Ri, M.B.; Giri, A.P.; Verstappen, F.W.A.; Schwab, W.; Bouwmeester, H.J. Metabolic engineering of terpenoid biosynthesis in plants. Phytochem. Rev. 2006, 5, 49–58. [Google Scholar] [CrossRef]
- Kuzuyama, T. Mevalonate and nonmevalonate pathways for the biosynthesis of isoprene units. Biosci. Biotechnol. Biochem. 2002, 66, 1619–1627. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Structure and dynamics of the isoprenoid pathway network. Mol. Plant 2012, 5, 318–333. [Google Scholar] [CrossRef]
- Picherky, E.; Geherson, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr.Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Nagegowda, D.A.; Gutensohn, M.; Wilkerson, C.; Dudareva, N. Two nearly identical terpene synthases catalyze the formation of nerolidol and linalool in snapdragon flowers. Plant J. 2008, 55, 224–239. [Google Scholar] [CrossRef]
- Australian Native Plant Society. Available online: http://www.anpsa.org.au (accessed on 20 March 2014).
- Song, A.; Wang, Y.; Liu, Y. Study on the chemical constituents of the essential oil of the leaves of Eucaliptus globulus Labill from China. Asian J. Trad. Med. 2009, 4, 134–140. [Google Scholar]
- Singh, H.P.; Mittal, S.; Kaur, S.; Batish, D.R.; Kohli, R.K. Characterization and antioxidant activity of essential oils from fresh and decaying leaves of Eucalyptus tereticornis. J. Agric. FoodChem. 2009, 57, 6962–6996. [Google Scholar]
- Tohidpour, A.; Sattari, M.; Ornidbaigi, R.; Yadegar, A.; Nazerri, J. Antibacterial effect of essential oils from two medicinal plants against methicillin-resistant Staphylococcus aureus (MRSA). Phytomedicine 2010, 17, 142–145. [Google Scholar] [CrossRef]
- Schnitzler, P.; Shön, K.; Reichling, J. Antiviral activity of Australian tree tea oil and eucalyptus oil against herpes simplex virus in cell culture. Pharmazie 2001, 56, 343–347. [Google Scholar]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Pharm. Biol. 2011, 49, 893–899. [Google Scholar] [CrossRef]
- Mulyaningsih, S.; Sporer, F.; Reichling, J.; Wink, M. Antibacterial activity of essential oils from Eucalyptus and of selected components against multidrug-resistance bacterial pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef]
- Yáñez-Rueda, X.; Pérez, O.G.; Meza, H. Larvicidal activity of essential oil isolated from Colombian Eucalyptus globulus against Aedes aegypti Linnaeus. Rev. Bistua 2010, 8, 71–77. [Google Scholar]
- Hofstetter, R.W.; Chen, Z.; Gaylor, M.L.; McMillin, J.D.; Wagner, M.R. Synergistic effect of α-pinene and exo-bervicomin on pine bark beetle and associated insects in Arizona. J. Appl. Entomol. 2008, 132, 387–397. [Google Scholar] [CrossRef]
- Cheng, S.S.; Huang, C.G.; Chen, Y.J.; Yu, J.J.; Chen, W.J.; Chang, S.T. Chemical composition and larvicidal activities of leaf essential oils from two eucalyptus species. Bioresour.Technol. 2009, 100, 452–456. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; González, A.; Moenne, A. Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef]
- González, A.; Contreras, R.A.; Moenne, A. Oligo-carrageenans enhance growth and contents of cellulose, essential oils and polyphenolic compounds in Eucalyptus globulus trees. Molecules 2013, 18, 840–851. [Google Scholar]
- González, A.; Moenne, F.; Contreras, R.A.; Gómez, M.; Moenne, A. Cross-talk among NADPH, ascorbate and glutathione synthesis and TRR/TRX activities enhances photosynthesis, basal metabolism, and growth in Eucalyptus globulus trees treated with OC kappa. Front. Plant Sci. 2014. submitted for publication. [Google Scholar]
- Ait-Ouazzou, A.; Lorán, S.; Bakkali, M.; Laglaoui, A.; Rota, C.; Herrera, A.; Pagán, R.; Conchello, P. Chemical composition and antimicrobial activity of essential oils of Thymus algeriensis, Eucalyptus globulus and Rosmarinus officinalis from Morocco. J. Sci. Food Agric. 2011, 91, 2643–2651. [Google Scholar] [CrossRef]
- Kim, S.I.; Yoon, J.S.; Jung, J.W.; Hong, K.B.; Ahn, Y.J.; Kwon, H.W. Toxicity and repellency of origanum essential oil and its components against Tribolium castaneum (Coleoptera: Tenebrionidae) adults. J. Asia-Pacif. Entomol. 2010, 13, 369–373. [Google Scholar] [CrossRef]
- Lee, S.E.; Lee, B.H.; Choi, W.S.; Park, B.S.; Kim, J.G.; Campbell, B.C. Fumigant toxicity of volatile natural products from Korean spices and medicinal plants towards the rice weevil Sitophilus oryzae (L.). Pest Manag. Sci. 2001, 57, 548–553. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, Y.; Lei, C.L. Evaluation of monoterpenes for the control of Triboilum castaneum (Herbst) and Sitophulus zeamaise Motschulsky. Nat. Prod. Res. 2009, 23, 1080–1088. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Klun, J.A.; Bryson, C.T.; Kobaisy, M.; Duke, S.O. Isolation and identification of mosquito bite deterrent terpenoids from leaves of American (Callirpa americana) and Japanese (Callirpa japonica) beautyberry. J. Agric. Food Chem. 2005, 53, 5948–5953. [Google Scholar] [CrossRef]
- Balmer, Y.; Koller, A.; del Val, G.; Manieri, W.; Schürmann, P.; Buchanan, B.B. Proteomics give insight into the regulatory function of chloroplast thioredoxins. Proc. Natl. Acad. Sci. USA 2003, 100, 370–375. [Google Scholar] [CrossRef]
- Olessen, U.H.; Petersen, J.G.; Garten, A.; Kiess, W.; Yoshino, J.; Imai, S.I.; Christensen, M.K.; Fistrup, P.; Thougaard, A.V.; Björkling, F.; et al. Target enzyme mutations are the molecular basis for resistance toward pharmacological inhibition of nicotinamide phosphoribosyl transferase. BMC Cancer 2010, 10, 677–690. [Google Scholar] [CrossRef]
- Arrigoni, O.; Pacciolla, C.; de Gara, L. Inhibition of galactonolactone dehydrogenase activity by lycorine. Boll. Soc. Ital. Biol. Sper. 1996, 72, 37–43. [Google Scholar]
- Griffith, O.W.; Meister, A. Potent and specific inhibition of glutathione synthesis by buthionine sulfoximine (S-n-butyl homocysteine sulfoximine). J. Biol. Chem. 1979, 254, 7558–7560. [Google Scholar]
- Gromer, S.; Arscott, L.D.; Williams, C.H.; Shirmer, R.H.; Becker, K. Human placenta thioredoxin reductase: Isolation of the selenoenzyme, steady sate kinetics, and inhibition by therapeutic gold compounds. J. Biol. Chem. 1998, 273, 20096–20101. [Google Scholar]
- Zar, J. Biostatistical Analysis; Prentice Hall/Pearson Inc.: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
González, A.; Gutiérrez-Cutiño, M.; Moenne, A. Oligo-Carrageenan Kappa-Induced Reducing Redox Status and Increase in TRR/TRX Activities Promote Activation and Reprogramming of Terpenoid Metabolism in Eucalyptus Trees. Molecules 2014, 19, 7356-7367. https://doi.org/10.3390/molecules19067356
González A, Gutiérrez-Cutiño M, Moenne A. Oligo-Carrageenan Kappa-Induced Reducing Redox Status and Increase in TRR/TRX Activities Promote Activation and Reprogramming of Terpenoid Metabolism in Eucalyptus Trees. Molecules. 2014; 19(6):7356-7367. https://doi.org/10.3390/molecules19067356
Chicago/Turabian StyleGonzález, Alberto, Marlen Gutiérrez-Cutiño, and Alejandra Moenne. 2014. "Oligo-Carrageenan Kappa-Induced Reducing Redox Status and Increase in TRR/TRX Activities Promote Activation and Reprogramming of Terpenoid Metabolism in Eucalyptus Trees" Molecules 19, no. 6: 7356-7367. https://doi.org/10.3390/molecules19067356
APA StyleGonzález, A., Gutiérrez-Cutiño, M., & Moenne, A. (2014). Oligo-Carrageenan Kappa-Induced Reducing Redox Status and Increase in TRR/TRX Activities Promote Activation and Reprogramming of Terpenoid Metabolism in Eucalyptus Trees. Molecules, 19(6), 7356-7367. https://doi.org/10.3390/molecules19067356