Metabolism of the Cyanogenic Glucosides in Developing Flax: Metabolic Analysis, and Expression Pattern of Genes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
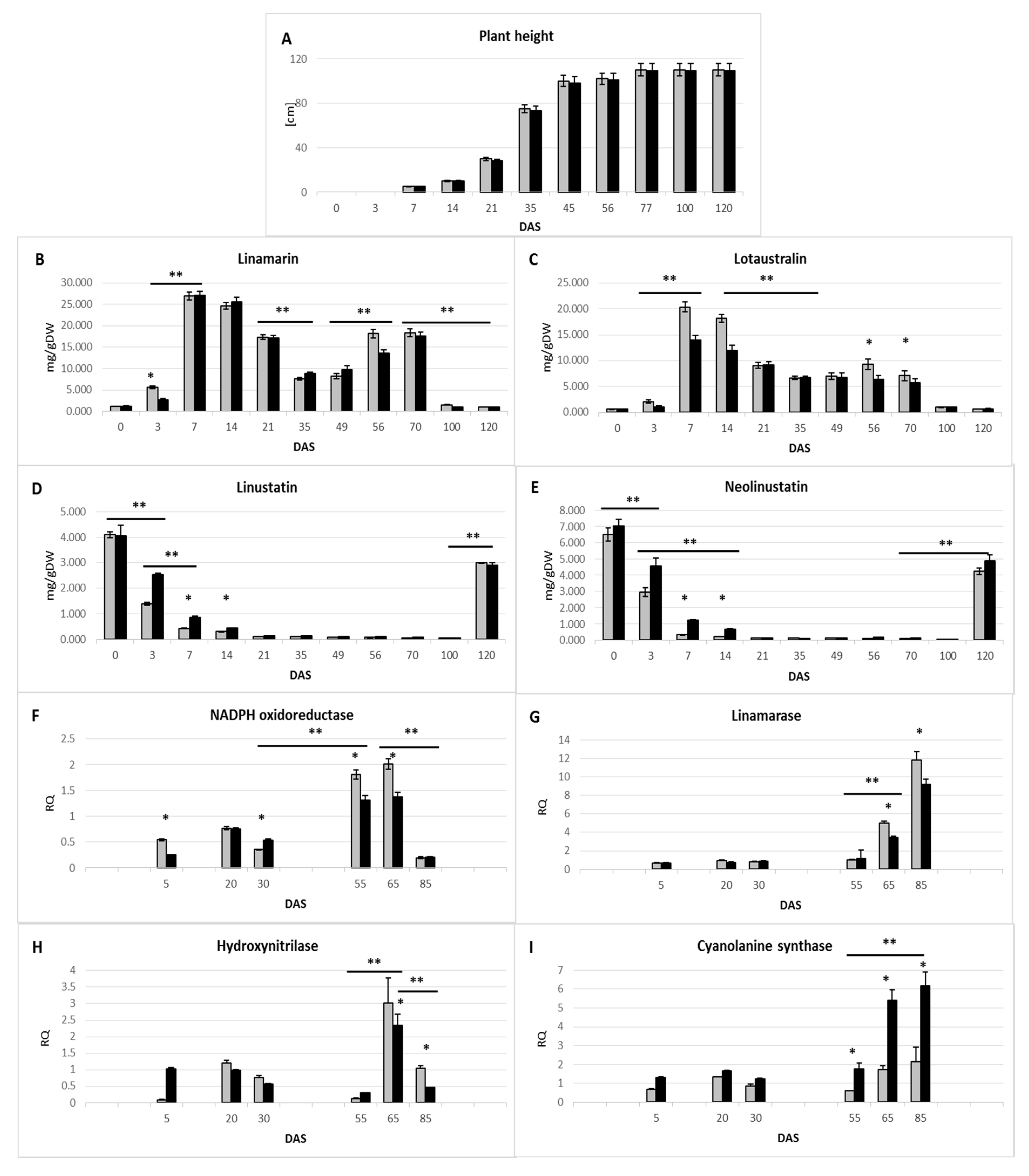
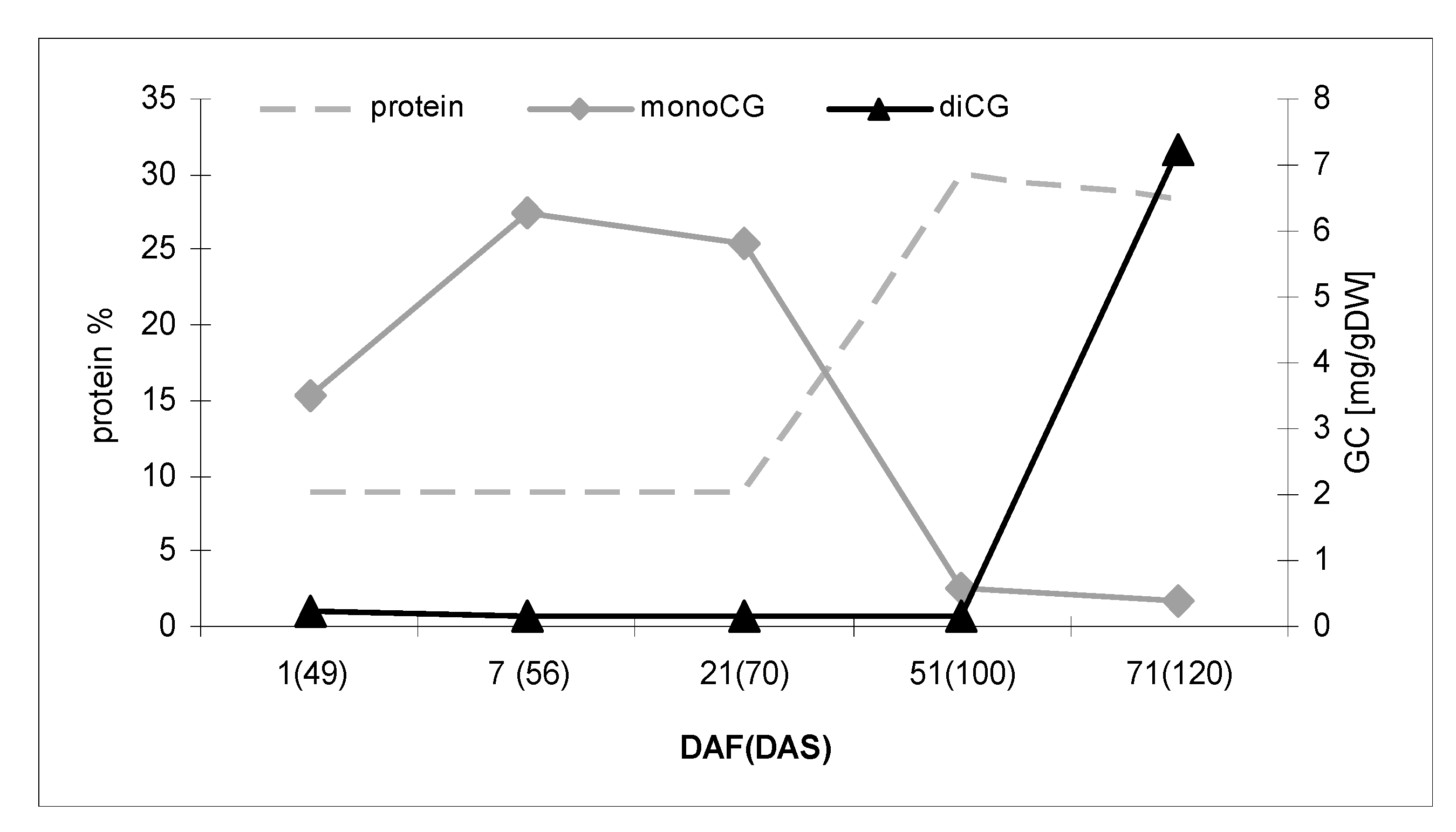
2.1. Assessment of Changes in Cyanogenic Glycoside Levels during Flax Growth
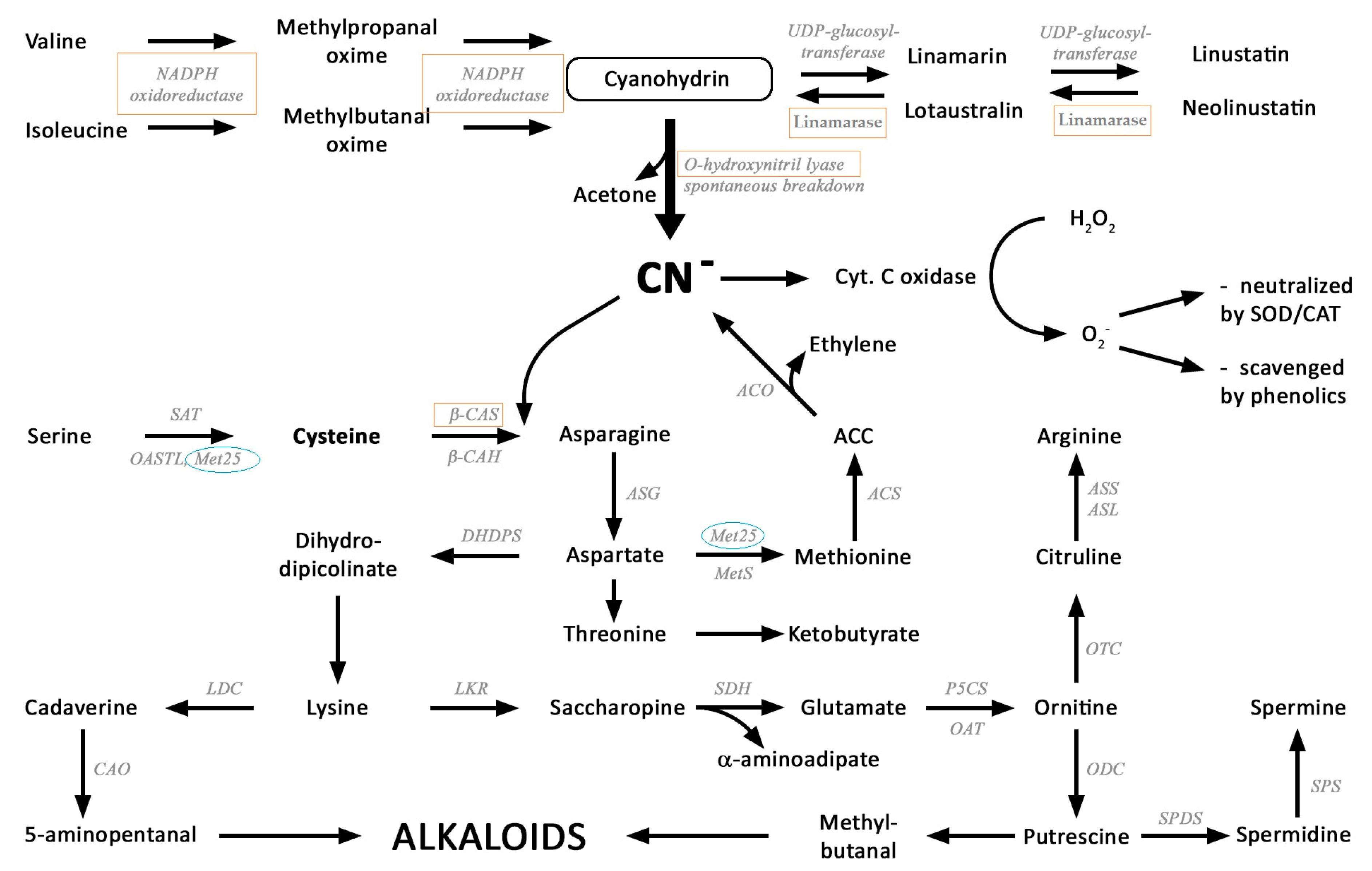
2.2. Identification of Key Enzymes for the Metabolism of Cyanogenic Glycosides
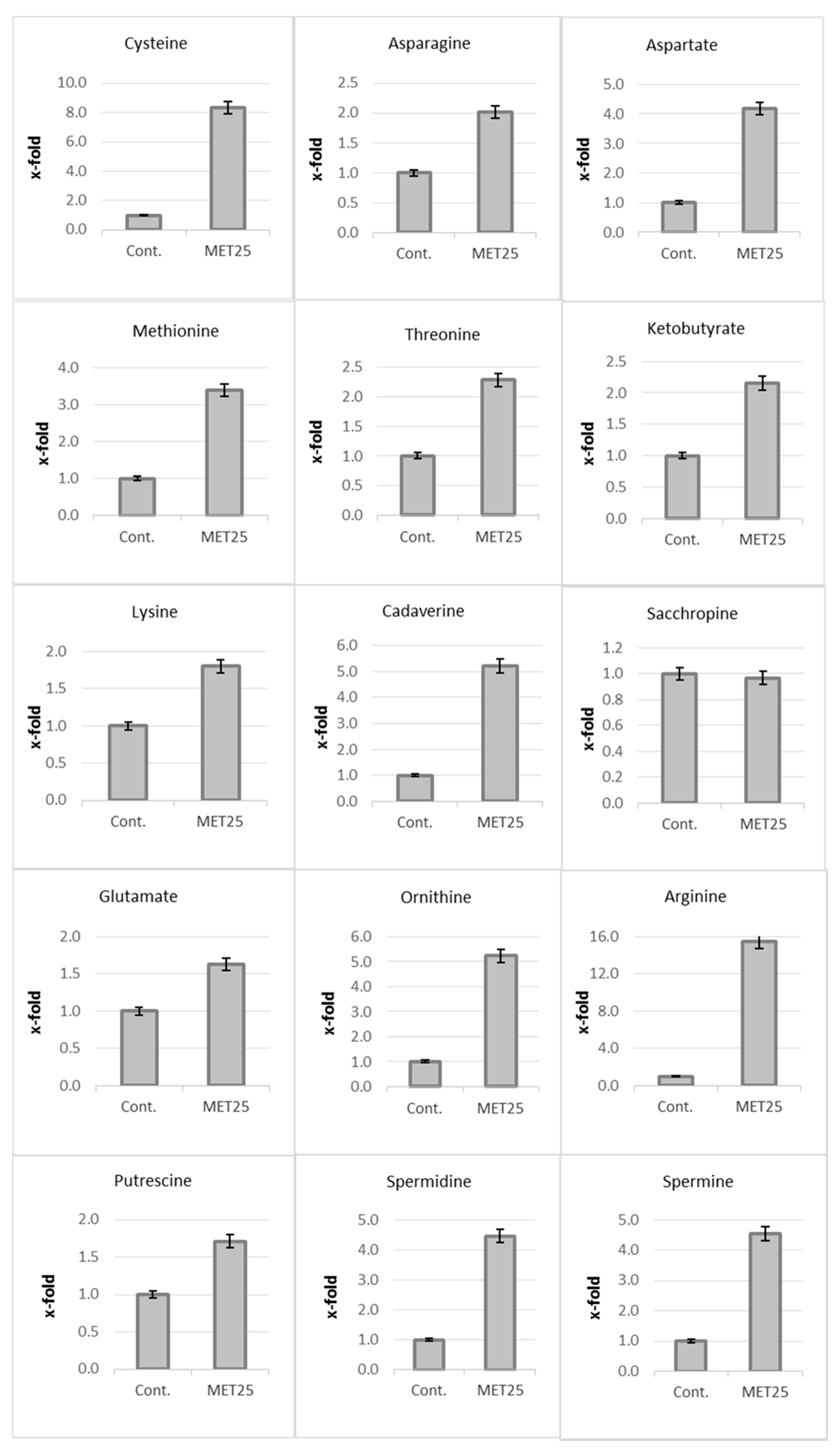
2.3. Assimilation of CN− into Amino Acids and Diamines
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Cyanogenic Glycosides (CG) Extraction
4.3. Separation and Identification Method for Cyanogenic Glycosides (CG)
4.4. Extraction of Metabolites for GC-MS Analysis
4.5. GC-MS Metabolite Profiling
4.6. Gene Expression Analysis with Quantitative PCR
4.7. Statistic Analysis
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cressey, P.; Reeve, J. Metabolism of cyanogenic glycosides: A review. Food Chem. Toxicol. 2019, 125, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Ganjewala, D.; Kumar, S.; Devi, A.; Ambika, K. Advances in cyanogenic glycosides biosynthesis and analyses in plants: A review. Acta Biol. Szeged. 2010, 54, 1–14. [Google Scholar]
- Davis, R.H. Cyanogens; Elsevier: New York, NY, USA, 1991; pp. 202–225. [Google Scholar]
- Vetter, J. Plant cyanogenic glycosides. Toxicon 2000, 38, 11–36. [Google Scholar] [CrossRef]
- Møller, B.L. Functional diversifications of cyanogenic gluco-sides. Curr. Opin. Plant Biol. 2010, 13, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Gleadow, R.M.; Woodrow, I.E. Constrains on effectiveness of cyanogenic glycosides in herbivore defense. J. Chem. Ecol. 2002, 28, 1301–1313. [Google Scholar] [CrossRef]
- Sanchez-Perez, R.; Jorgensen, K.; Motawia, M.S.; Dicenta, F.; Møller, B.L. Tissue and cellular localization of individual beta-glycosidases using a substrate specific sugar reducing assay. Plant J. 2009, 60, 894–906. [Google Scholar] [CrossRef]
- Siegień, I.; Adamczuk, A.; Wróblewska, K. Light affects in vitro organogenesis of Linum usitatissimum L. and its cyanogenic potential. Acta Physiol. Plant. 2012, 35, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Siritunga, D.; Sayre, R. Generation of cyanogen-free transgenic cassava. Planta 2003, 217, 367–373. [Google Scholar] [CrossRef]
- Czuj, T.; Zuk, M.; Starzycki, M.; Amir, R.; Szopa, J. Engineering increases in sulfur amino acid contents in flax by overexpressing the yeast met25 gene. Plant Sci. 2009, 177, 584–592. [Google Scholar] [CrossRef]
- Goudenhooft, C.; Bourmaud, A.; Baley, C. Flax (Linum usitatissimum L.) Fibers for Composite Reinforcement: Exploring the Link Between Plant Growth, Cell Walls Development, and Fiber Properties. Front. Plant Sci. 2019, 10, 411. [Google Scholar] [CrossRef] [Green Version]
- Dalisay, D.; Kim, K.W.; Lee, C.; Yang, H.; Rübel, O.; Bowen, B.P.; Davin, L.B.; Lewis, N.G. Dirigent Protein-Mediated Lignan and Cyanogenic Glucoside Formation in Flax Seed: Integrated Omics and MALDI Mass Spectrometry Imaging. J. Nat. Prod. 2015, 78, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Krech, M.J.; Fieldes, M.A. Analysis of the developmental regulation of the cyanogenic compounds in seedlings of two lines of Linum usitatissimum L. Can. J. Bot. 2003, 81, 1029–1038. [Google Scholar] [CrossRef] [Green Version]
- Hills, M.J. Control of storage-product synthesis in seeds. Curr. Opin. Plant. Boil. 2004, 7, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K. A role for cyanide, derived from ethylene biosynthesis, in the development of stress symptoms. Physiol. Plant. 1996, 97, 772–775. [Google Scholar] [CrossRef]
- Caraparo, A.; Freund, R.; Gustafson, T.A. 14-3-3 (epsilon) interacts with insulin-like growth factor i and insulin receptor substrat iin a phosphoserine-dependent manner. J. Biol. Chem. 1997, 272, 11663–11669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.; Mitsuhara, I.; Feng, J.; Iwai, T.; Hasegawa, M.; Ohashi, Y. Cyanide, a coproduct of plant hormone ethylene biosynthesis, contributes to the resistance of rice to blast fungus. Plant. Physiol. 2010, 155, 502–514. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Zhang, D.-W.; Zhu, F.; Tang, H.; Lv, X.; Cheng, J.; Xie, H.-F.; Lin, H.-H. A novel role for cyanide in the control of cucumber (Cucumis sativus L.) seedlings response to environmental stress. Plant. Cell Environ. 2012, 35, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Herchi, W.; Bouali, I.; Bahashwan, S.; Rochut, S.; Boukhchina, S.; Kallel, H.; Pepe, C. Changes in phospholipid composition, protein content and chemical properties of flaxseed oil during development. Plant. Physiol. Biochem. 2012, 54, 1–5. [Google Scholar] [CrossRef]
- Siegień, I.; Bogatek, R. Cyanide action in plants—From toxic to regulatory. Acta Physiol. Plant. 2006, 28, 483–497. [Google Scholar] [CrossRef]
- Yu, L.; Liu, Y.; Xu, F. Comparative transcriptome analysis reveals significant differences in the regulation of gene expression between hydrogen cyanide- and ethylene-treated Arabidopsis thaliana. BMC Plant. Boil. 2019, 19, 92. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Juergens, M.; Jez, J.M. Structure of soybean beta-cyanoalanine synthase and the molecular basis for cyanide detoxification in plants. Plant Cell. 2012, 24, 2696–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, D.; Ravasz, D.; Chinopoulos, C. The effect of 2-ketobutyrate on mitochondrial substrate-level phosphorylation. Neurochem. Res. 2019, 44, 2301–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Żuk, M.; Szperlik, J.; Hnitecka, A.; Szopa, J. Temporal biosynthesis of flavone constituents in flax growth stages. Plant. Physiol. Biochem. 2019, 142, 234–245. [Google Scholar] [CrossRef] [PubMed]
- García, I.; Arenas-Alfonseca, L.; Moreno, I.; Gotor, C.; Romero, L.C. Hcn regulates cellular processes through posttranslational modification of proteins by S-cyanylation. Plant Physiol. 2019, 179, 107–123. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuk, M.; Pelc, K.; Szperlik, J.; Sawula, A.; Szopa, J. Metabolism of the Cyanogenic Glucosides in Developing Flax: Metabolic Analysis, and Expression Pattern of Genes. Metabolites 2020, 10, 288. https://doi.org/10.3390/metabo10070288
Zuk M, Pelc K, Szperlik J, Sawula A, Szopa J. Metabolism of the Cyanogenic Glucosides in Developing Flax: Metabolic Analysis, and Expression Pattern of Genes. Metabolites. 2020; 10(7):288. https://doi.org/10.3390/metabo10070288
Chicago/Turabian StyleZuk, Magdalena, Katarzyna Pelc, Jakub Szperlik, Agnieszka Sawula, and Jan Szopa. 2020. "Metabolism of the Cyanogenic Glucosides in Developing Flax: Metabolic Analysis, and Expression Pattern of Genes" Metabolites 10, no. 7: 288. https://doi.org/10.3390/metabo10070288
APA StyleZuk, M., Pelc, K., Szperlik, J., Sawula, A., & Szopa, J. (2020). Metabolism of the Cyanogenic Glucosides in Developing Flax: Metabolic Analysis, and Expression Pattern of Genes. Metabolites, 10(7), 288. https://doi.org/10.3390/metabo10070288