
In Silico Analyses and Cytotoxicity Study of Asiaticoside and Asiatic Acid from Malaysian Plant as Potential mTOR Inhibitors

 , ,
, ,
Abstract
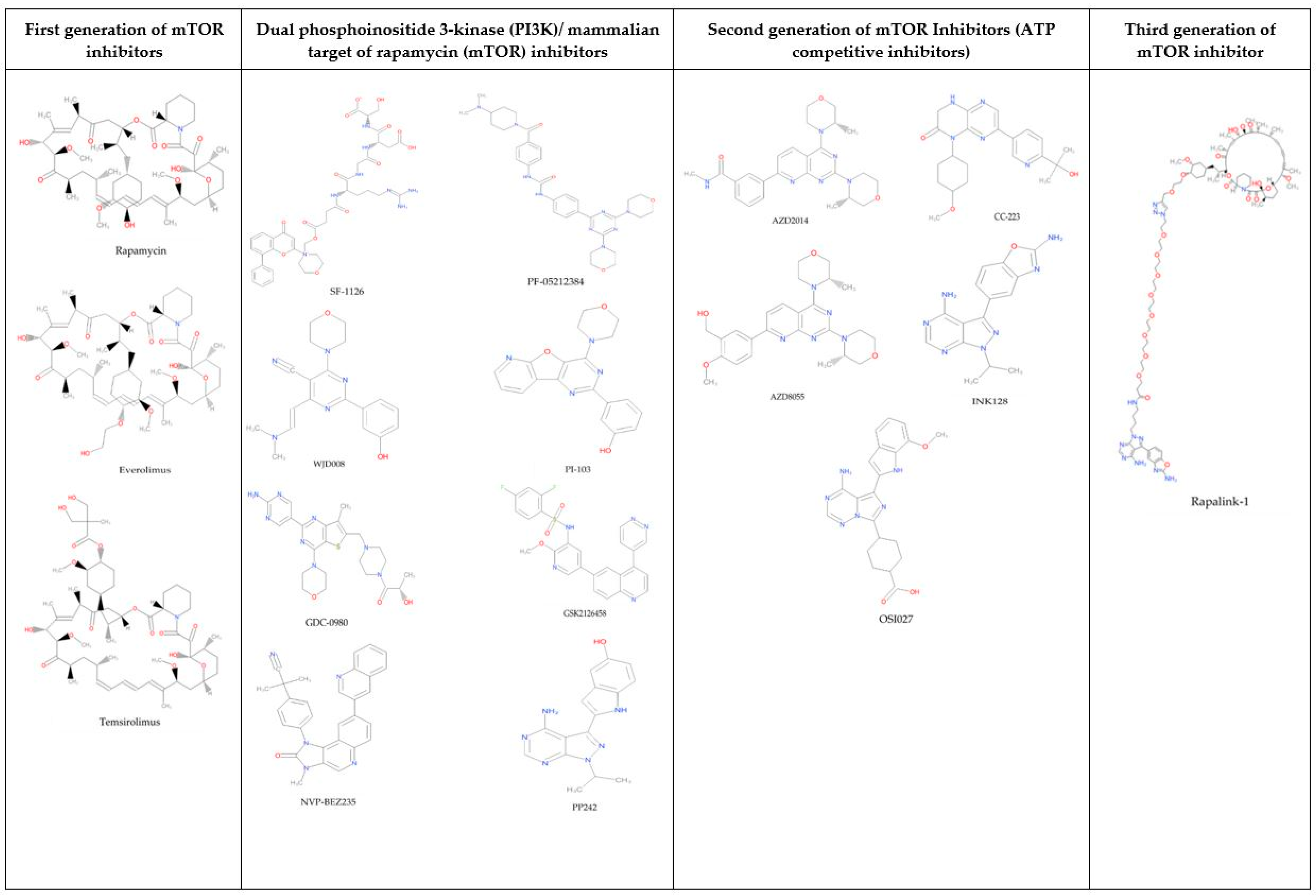
:1. Introduction
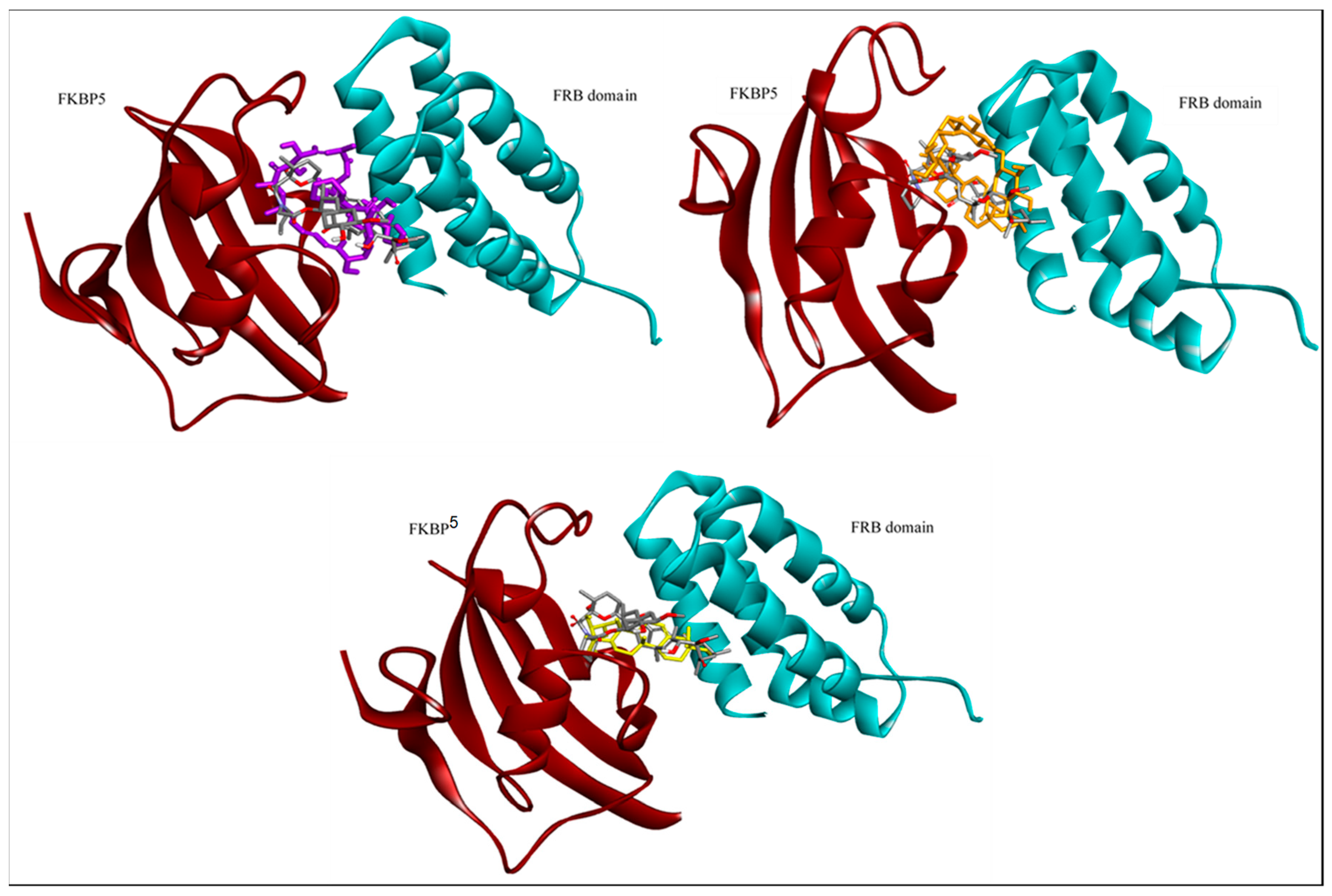
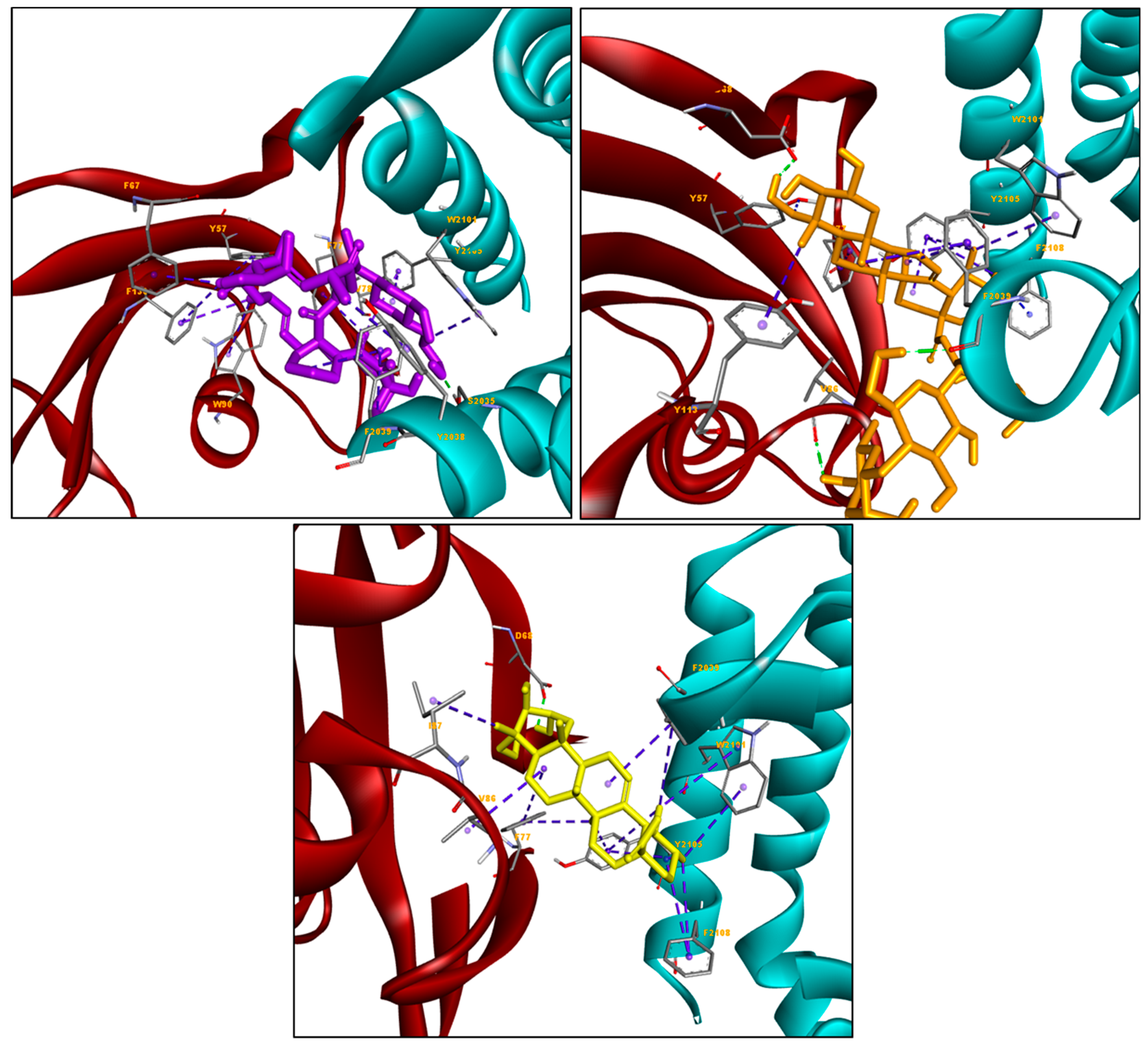
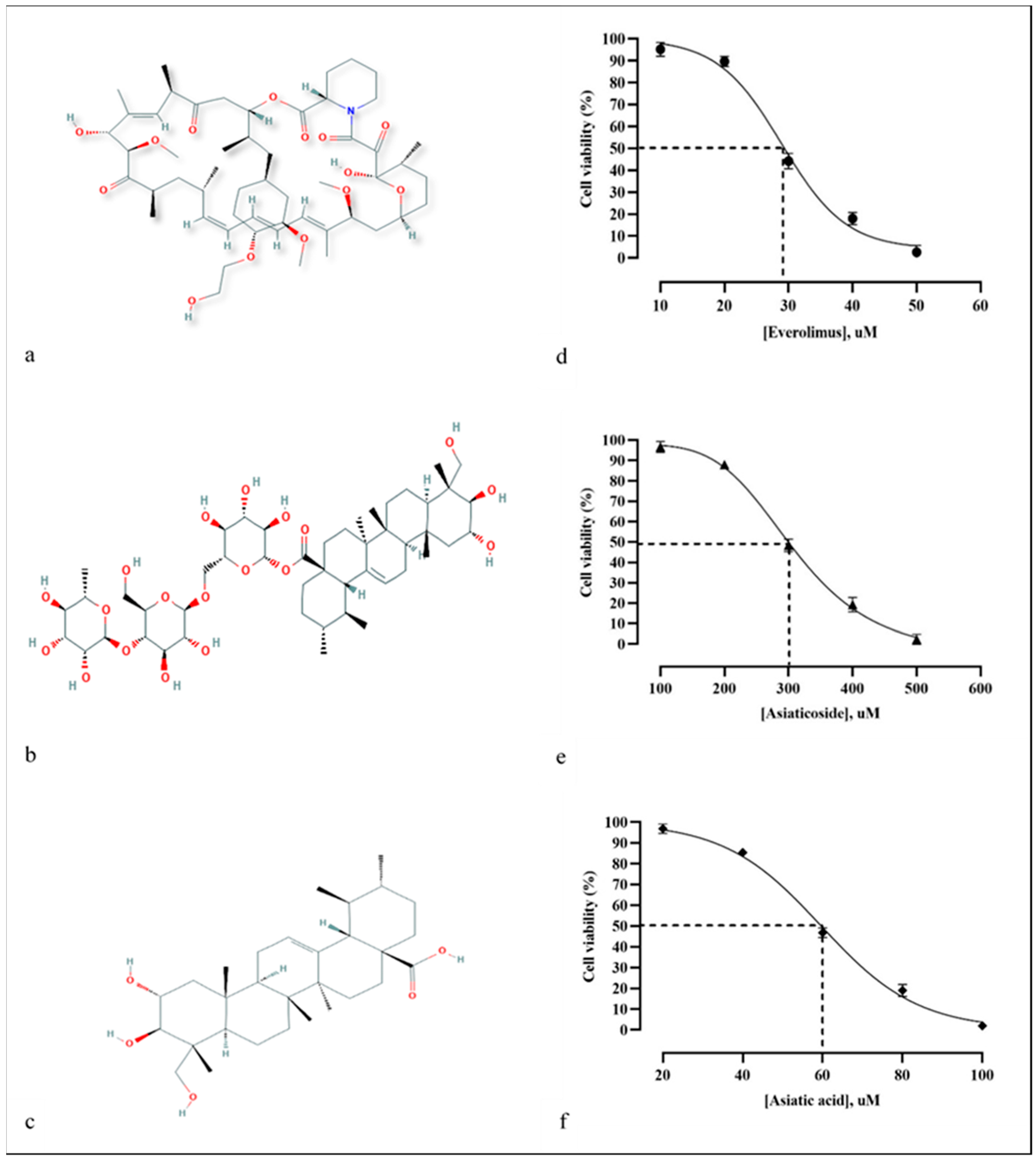
2. Results and Discussion
3. Materials and Methods
3.1. Accession of the Target Protein
3.2. Ligand Selection
3.3. Analysis of Target Active Binding Sites
3.4. Control Docking
3.5. Virtual Screening
3.6. Molecular Docking
3.7. Cell Culture and Treatment
3.8. Cytotoxicity Assay
- Absorbance of treated cells: Cells + Media + Drug concentration + DMSO.
- Absorbance of controlled cells: Cells + Media + DMSO.
- Absorbance of blank cells: Media.
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rosset, C.; Vairo, F.; Bandeira, I.C.; Correia, R.L.; De Goes, F.V.; Da Silva, R.T.B.; Bueno, L.S.M.; de Miranda Gomes, M.C.S.; Galvão, H.D.C.R.; Neri, J.I.; et al. Molecular analysis of TSC1 and TSC2 genes and phenotypic correlations in Brazilian families with tuberous sclerosis. PLoS ONE 2017, 12, e0185713. [Google Scholar] [CrossRef] [PubMed]
- Hyman, M.H.; Whittemore, V.H. National Institutes of Health consensus conference: Tuberous sclerosis complex. Arch. Neurol. 2000, 57, 662–665. [Google Scholar] [CrossRef]
- Astrinidis, A.; Cash, T.P.; Hunter, D.S.; Walker, C.L.; Chernoff, J.; Henske, E.P. Tuberin, the tuberous sclerosis complex 2 tumor suppressor gene product, regulates Rho activation, cell adhesion and migration. Oncogene 2002, 21, 8470–8476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napolioni, V.; Curatolo, P. Genetics and molecular biology of tuberous sclerosis complex. Curr. Genom. 2008, 9, 475–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wang, X.; Proud, C.G. mTOR inhibitors in cancer therapy. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Blagosklonny, M.V.; Hall, M.N. Growth and aging: A common molecular mechanism. Aging 2009, 1, 357. [Google Scholar] [CrossRef] [Green Version]
- Hands, S.L.; Proud, C.G.; Wyttenbach, A. mTOR’s role in ageing: Protein synthesis or autophagy? Aging 2009, 1, 586. [Google Scholar] [CrossRef] [Green Version]
- Antikainen, H.; Driscoll, M.; Haspel, G.; Dobrowolski, R. TOR-mediated regulation of metabolism in aging. Aging Cell 2017, 16, 1219–1233. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, X.; Zhang, J. mTOR signaling in cancer and mtor inhibitors in solid tumor targeting therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viana, S.D.; Reis, F.; Alves, R. Therapeutic use of mTOR inhibitors in renal diseases: Advances, drawbacks, and challenges. Oxid. Med. Cell. Longev. 2018, 2018, 3693625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carew, J.S.; Kelly, K.R.; Nawrocki, S.T. Mechanisms of mTOR inhibitor resistance in cancer therapy. Targeted Oncol. 2011, 6, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.M.; Mita, A.C.; Chu, Q.S.; Rowinsky, E.K.; Fetterly, G.J.; Goldston, M.; Patnaik, A.; Mathews, L.; Ricart, A.D.; Mays, T.; et al. Phase I trial of the novel mammalian target of rapamycin inhibitor deforolimus (AP23573; MK-8669) administered intravenously daily for 5 days every 2 weeks to patients with advanced malignancies. J. Clin. Oncol. 2008, 26, 361–367. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, A.; Faivre, S.; Burris III, H.A.; Rea, D.; Papadimitrakopoulou, V.; Shand, N.; Lane, H.A.; Hazell, K.; Zoellner, U.; Kovarik, J.M.; et al. Phase I pharmacokinetic and pharmacodynamic study of the oral mammalian target of rapamycin inhibitor everolimus in patients with advanced solid tumors. J. Clin. Oncol. 2008, 26, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Raymond, E.; Alexandre, J.; Faivre, S.; Vera, K.; Materman, E.; Boni, J.; Leister, C.; Korth-Bradley, J.; Hanauske, A.; Armand, J.P. Safety and pharmacokinetics of escalated doses of weekly intravenous infusion of CCI-779, a novel mTOR inhibitor, in patients with cancer. J. Clin. Oncol. 2004, 22, 2336–2347. [Google Scholar] [CrossRef]
- Tabernero, J.; Rojo, F.; Calvo, E.; Burris, H.; Judson, I.; Hazell, K.; Martinelli, E.; Cajal, S.R.Y.; Jones, S.; Vidal, L.; et al. Dose-and schedule-dependent inhibition of the mammalian target of rapamycin pathway with everolimus: A phase I tumor pharmacodynamic study in patients with advanced solid tumors. J. Clin. Oncol. 2008, 26, 1603–1610. [Google Scholar] [CrossRef]
- Switon, K.; Kotulska, K.; Janusz-Kaminska, A.; Zmorzynska, J.; Jaworski, J. Tuberous sclerosis complex: From molecular biology to novel therapeutic approaches. IUBMB Life 2016, 68, 955–962. [Google Scholar] [CrossRef]
- Mingarelli, A.; Vignoli, A.; La Briola, F.; Peron, A.; Giordano, L.; Banderali, G.; Canevini, M.P. Dramatic relapse of seizures after everolimus withdrawal. Eur. J. Paediatr. Neurol. 2018, 22, 203–206. [Google Scholar] [CrossRef]
- Martelli, A.M.; Buontempo, F.; McCubrey, J.A. Drug discovery targeting the mTOR pathway. Clin. Sci. 2018, 132, 543–568. [Google Scholar] [CrossRef]
- Herschbein, L.; Liesveld, J.L. Dueling for dual inhibition: Means to enhance effectiveness of PI3K/Akt/mTOR inhibitors in AML. Blood Rev. 2018, 32, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Wise-Draper, T.M.; Moorthy, G.; Salkeni, M.A.; Karim, N.A.; Thomas, H.E.; Mercer, C.A.; Beg, M.S.; O’Gara, S.; Olowokure, O.; Fathallah, H.; et al. A phase Ib study of the dual PI3K/mTOR inhibitor dactolisib (BEZ235) combined with everolimus in patients with advanced solid malignancies. Target. Oncol. 2017, 12, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, G.I.; Bell-McGuinn, K.M.; Molina, J.R.; Bendell, J.; Spicer, J.; Kwak, E.L.; Pandya, S.S.; Millham, R.; Borzillo, G.; Pierce, K.J.; et al. First-in-human study of PF-05212384 (PKI-587), a small-molecule, intravenous, dual inhibitor of PI3K and mTOR in patients with advanced cancer. Clin. Cancer Res. 2015, 21, 1888–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Proud, C.G. Signaling crosstalk between the mTOR complexes. Translation 2014, 2, e28174. [Google Scholar] [CrossRef]
- Ghosh, A.P.; Marshall, C.B.; Coric, T.; Shim, E.H.; Kirkman, R.; Ballestas, M.E.; Ikura, M.; Bjornsti, M.A.; Sudarshan, S. Point mutations of the mTOR-RHEB pathway in renal cell carcinoma. Oncotarget 2015, 6, 17895. [Google Scholar] [CrossRef] [Green Version]
- Rodrik-Outmezguine, V.S.; Okaniwa, M.; Yao, Z.; Novotny, C.J.; McWhirter, C.; Banaji, A.; Won, H.; Wong, W.; Berger, M.; de Stanchina, E.; et al. Overcoming mTOR resistance mutations with a new-generation mTOR inhibitor. Nature 2016, 534, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Barlow, A.D.; Xie, J.; Moore, C.E.; Campbell, S.C.; Shaw, J.A.M.; Nicholson, M.L.; Herbert, T.P. Rapamycin toxicity in MIN6 cells and rat and human islets is mediated by the inhibition of mTOR complex 2 (mTORC2). Diabetologia 2012, 55, 1355–1365. [Google Scholar] [CrossRef] [Green Version]
- Mohamad, S.; Zin, N.M.; Wahab, H.A.; Ibrahim, P.; Sulaiman, S.F.; Zahariluddin, A.S.M.; Noor, S.S.M. Antituberculosis potential of some ethnobotanically selected Malaysian plants. J. Ethnopharmacol. 2011, 133, 1021–1026. [Google Scholar] [CrossRef]
- Ikram, N.K.K.; Durrant, J.D.; Muchtaridi, M.; Zalaludin, A.S.; Purwitasari, N.; Mohamed, N.; Rahim, A.S.A.; Lam, C.K.; Normi, Y.M.; Rahman, N.A.; et al. A virtual screening approach for identifying plants with anti H5N1 neuraminidase activity. J. Chem. Inf. Model. 2015, 55, 308–316. [Google Scholar] [CrossRef]
- Hariono, M.; Abdullah, N.; Damodaran, K.V.; Kamarulzaman, E.E.; Mohamed, N.; Hassan, S.S.; Shamsuddin, S.; Wahab, H.A. Potential new H1N1 neuraminidase inhibitors from ferulic acid and vanillin: Molecular modelling, synthesis and in vitro assay. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.V.; Choi, S.B.; Wahab, H.A.; Lim, T.S.; Choong, Y.S. Applications of ensemble docking in potential inhibitor screening for Mycobacterium tuberculosis isocitrate lyase using a local plant database. J. Chem. Inf. Model. 2019, 59, 2487–2495. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Wachtel-Galor, S. Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Nisar, B.; Sultan, A.; Rubab, S.L. Comparison of Medicinally Important Natural Products versus Synthetic Drugs—A Short Commentary. Nat. Prod. Chem. Res. 2018, 6, 308. [Google Scholar] [CrossRef]
- Karimi, A.; Majlesi, M.; Rafieian-Kopaei, M. Herbal versus synthetic drugs; beliefs and facts. J. Nephropharmacol. 2015, 4, 27. [Google Scholar] [PubMed]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pingaew, R.; Mandi, P.; Prachayasittikul, V.; Prachayasittikul, S.; Ruchirawat, S.; Prachayasittikul, V. Synthesis, molecular docking, and QSAR study of sulfonamide-based indoles as aromatase inhibitors. Eur. J. Med. Chem. 2018, 143, 1604–1615. [Google Scholar] [CrossRef]
- Rangaraju, A.; Rao, A.V. A review on molecular docking: Novel tool in drug design and analysis. J. Harmon Res. Pharm. 2013, 2, 215–221. [Google Scholar]
- Yingchun, L.; Huihan, W.; Rong, Z.; Guojun, Z.; Ying, Y.; Zhuogang, L. Antitumor activity of asiaticoside against multiple myeloma drug-resistant cancer cells is mediated by autophagy induction, activation of effector caspases, and inhibition of cell migration, invasion, and STAT-3 signaling pathway. Med. Sci. Monit. 2019, 25, 1355–1361. [Google Scholar] [CrossRef]
- Al-Saeedi, F.J. Study of the cytotoxicity of asiaticoside on rats and tumour cells. BMC Cancer 2014, 14, 220. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Cao, Q.X.; Zhai, F.R.; Yang, S.Q.; Zhang, H.X. Asiatic acid exerts anticancer potential in human ovarian cancer cells via suppression of PI3K/Akt/mTOR signalling. Pharm. Biol. 2016, 54, 2377–2382. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Y.; Xu, Q.W.; Xu, H.; Huang, Z.H. Asiatic acid promotes p21 (WAF1/CIP1) protein stability through attenuation of NDR1/2 dependent phosphorylation of p21 (WAF1/CIP1) in HepG2 human hepatoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 963–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.L.; Yang, X.Y.; Jung, H.J.; Kim, S.Y.; Jung, S.Y.; Choi, D.Y.; Park, W.C.; Park, H. Asiatic acid induces colon cancer cell growth inhibition and apoptosis through mitochondrial death cascade. Biol. Pharm. Bull. 2009, 32, 1399–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.L.; Kuo, P.L.; Lin, L.T.; Lin, C.C. Asiatic acid, a triterpene, induces apoptosis and cell cycle arrest through activation of extracellular signal-regulated kinase and p38 mitogen-activated protein kinase pathways in human breast cancer cells. J. Pharmacol. Exp. Ther. 2005, 313, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Mannangatti, P.; Naidu, K.N. Indian herbs for the treatment of neurodegenerative disease. Adv. Neurobiol. 2016, 12, 323–336. [Google Scholar] [PubMed]
- Kim, S.R.; Koo, K.A.; Lee, M.K.; Park, H.G.; Jew, S.S.; Cha, K.H.; Kim, Y.C. Asiatic acid derivatives enhance cognitive performance partly by improving acetylcholine synthesis. J. Pharm. Pharmacol. 2004, 56, 1275–1282. [Google Scholar] [CrossRef]
- Nasir, M.N.; Habsah, M.; Zamzuri, I.; Rammes, G.; Hasnan, J.; Abdullah, J. Effects of asiatic acid on passive and active avoidance task in male Spraque–Dawley rats. J. Ethnopharmacol. 2011, 134, 203–209. [Google Scholar] [CrossRef]
- Lee, J.; Jung, E.; Kim, Y.; Park, J.; Park, J.; Hong, S.; Kim, J.; Hyun, C.; Kim, Y.S.; Park, D. Asiaticoside induces human collagen I synthesis through TGFβ receptor I kinase (TβRI kinase)-independent Smad signaling. Planta Med. 2006, 72, 324–328. [Google Scholar] [CrossRef]
- Chao, P.C.; Yin, M.C.; Mong, M.C. Anti-apoptotic and anti-glycative effects of asiatic acid in the brain of D-galactose treated mice. Food Funct. 2015, 6, 542–548. [Google Scholar] [CrossRef]
- Bonte, F.; Dumas, M.; Chaudagne, C.; Meybeck, A. Influence of asiatic acid, madecassic acid, and asiaticoside on human collagen I synthesis. Planta Med. 1994, 60, 133–135. [Google Scholar] [CrossRef]
- Ganesan, A. The impact of natural products upon modern drug discovery. Curr. Opin. Chem. Biol. 2008, 12, 306–317. [Google Scholar] [CrossRef]
- Arnott, J.A.; Planey, S.L. The influence of lipophilicity in drug discovery and design. Expert. Opin. Drug Discov. 2012, 7, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Huang, F.; Zhang, J.; Zhu, G.; Yang, M.; Xiao, P. Cytotoxic cycloartane triterpene saponins from Actaea asiatica. J. Nat. Prod. 2006, 69, 500–1502. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Zhang, S.H.; Zhen, R.X.; Xu, X.D.; Zhen, Y.S. Asiaticoside inducing apoptosis of tumor cells and enhancing anti-tumor activity of vincristine. Chin. J. Canc. 2004, 23, 1599–1604. [Google Scholar]
- Kist, R.; Caceres, R.A. New potential inhibitors of mTOR: A computational investigation integrating molecular docking, virtual screening and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2017, 35, 3555–3568. [Google Scholar] [CrossRef]
- Palavra, F.; Robalo, C.; Reis, F. Recent advances and challenges of mTOR inhibitors use in the treatment of patients with tuberous sclerosis complex. Oxid. Med. Cell. Longev. 2017, 2017, 9820181. [Google Scholar] [CrossRef]
- Shim, P.J.; Park, J.H.; Chang, M.S.; Lim, M.J.; Kim, D.H.; Jung, Y.H.; Jew, S.S.; Park, E.H.; Kim, H.D. Asiaticoside mimetics as wound healing agent. Bioorganic Med. Chem. Lett. 1996, 6, 2937–2940. [Google Scholar] [CrossRef]
- Rush, W.R.; Murray, G.R.; Graham, D.J.M. The comparative steady-state bioavailability of the active ingredients of Madecassol. Eur. J. Drug Metab. Pharmacokinet. 1993, 18, 323–326. [Google Scholar] [CrossRef]
- Zheng, X.C.; Wang, S.H. Determination of asiatic acid in beagle dog plasma after oral administration of Centella asiatica extract by precolumn derivatization RP-HPLC. J. Chromatogr. B 2009, 877, 477–481. [Google Scholar] [CrossRef]
- Gill, R.; Tripathy, A.; Bairy, K.L. Current status of mTOR inhibitors as novel therapeutic agents. J. Glob. Pharma Technol. 2009, 6, 14–34. [Google Scholar]
- Lichota, A.; Gwozdzinski, K. Anticancer activity of natural compounds from plant and marine environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef] [Green Version]
- Meeran, N.; Fizur, M.; Goyal, S.N.; Suchal, K.; Sharma, C.; Patil, C.R.; Ojha, S.K. Pharmacological properties, molecular mechanisms, and pharmaceutical development of asiatic acid: A pentacyclic triterpenoid of therapeutic promise. Front. Pharmacol. 2018, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Cancer 2002, 2, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Brinkhaus, B.; Lindner, M.; Schuppan, D.; Hahn, E.G. Chemical, pharmacological and clinical profile of the East Asian medical plant Centella aslatica. Phytomedicine 2000, 7, 427–448. [Google Scholar] [CrossRef]
- Loiseau, A.; Mercier, M. Centella asiatica and skin care. Cosmet. Toilet. 2000, 115, 63–67. [Google Scholar]
- Ling, A.P.K.; Marziah, M.; Tan, S.E. Triterpenoids distribution in whole plant and callus cultures of Centella asiatica accessions. In Interdisciplinary Approaches in Natural Products Research, Proceedings of the 16th National Seminar on Natural Products, Serdang, Malaysia, 24–25 October 2000; Department of Chemistry, University Putra Malaysia: Seri Kembangan, Malaysia, 2001. [Google Scholar]
- Ling, A.P.K. Triterpene Production in Centella asiatica (L.) Urban (Pegaga) Callus and Cell Suspension Cultures. Ph.D. Thesis, Universiti Putra Malaysia, Seri Kembangan, Malaysia, 2004. [Google Scholar]
- Oyedeji, O.A.; Afolayan, A.J. Chemical composition and antibacterial activity of the essential oil of Centella asiatica. Growing in South Africa. Pharm. Biol. 2005, 43, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Mook-Jung, I.; Shin, J.E.; Yun, S.H.; Huh, K.; Koh, J.Y.; Park, H.K.; Jew, S.S.; Jung, M.W. Protective effects of asiaticoside derivatives against beta-amyloid neurotoxicity. J. Neurosci. Res. 1999, 58, 417–425. [Google Scholar] [CrossRef]
- Shukla, A.; Rasik, A.M.; Dhawan, B.N. Asiaticoside-induced elevation of antioxidant levels in healing wounds. Phytother. Res. 1999, 13, 50–54. [Google Scholar] [CrossRef]
- Cheng, C.L.; Guo, J.S.; Luk, J.; Koo, M.W.L. The healing effects of Centella extract and asiaticoside on acetic acid induced gastric ulcers in rats. Life Sci. 2004, 74, 2237–2249. [Google Scholar] [CrossRef]
- Guo, J.S.; Cheng, C.L.; Koo, M.W.L. Inhibitory effects of Centella asiatica water extract and asiaticoside on inducible nitric oxide synthase during gastric ulcer healing in rats. Planta Med. 2004, 70, 1150–1154. [Google Scholar] [CrossRef]
- Wijeweera, P.; Arnason, J.T.; Koszycki, D.; Merali, Z. Evaluation of anxiolytic properties of Gotukola-(Centella asiatica) extracts and asiaticoside in rat behavioral models. Phytomedicine 2006, 13, 668–676. [Google Scholar] [CrossRef]
- Dong, M.S.; Jung, S.H.; Kim, H.J.; Kim, J.R.; Zhao, L.X.; Lee, E.S.; Lee, E.J.; Yi, J.B.; Lee, N.; Cho, Y.B.; et al. Structure-related cytotoxicity and anti-hepatofibric effect of asiatic acid derivatives in rat hepatic stellate cell-line, HSC-T6. Arch. Pharm. Res. 2004, 27, 512–517. [Google Scholar] [CrossRef]
- Suguna, L.; Sivakumar, P.; Chandrakasan, G. Effects of Centella asiatica extract on dermal wound healing in rats. Indian J. Exp. Biol. 1996, 34, 1208–1211. [Google Scholar] [PubMed]
- Komarcević, A. The modern approach to wound treatment. Med. Pregl. 2000, 53, 363–368. [Google Scholar] [PubMed]
- Appa Rao, M.V.R.; Srinivasan, K.; Rao, K.T.; Rao, T.; Rao, K.; Appa, R.; Rao, T.K. The effect of Mandookaparni (Centella asiatica) on the general mental ability (Medhya) of mentally retarded children. JRIM 1973, 8, 9–16. [Google Scholar]
- Boely, C. Indication of titrated extract of Centella asiatica in phlebology. Gaz. Med. France 1975, 82, 741–744. [Google Scholar]
- Anand, T.; Mahadeva, N.; Phani, K.G.; Farhath, K. Antioxidant and DNA Damage Preventive Properties of Centella asiatica (L) Urb. Pharmacog. J. 2010, 2, 53–58. [Google Scholar] [CrossRef]
- März, A.M.; Fabian, A.K.; Kozany, C.; Bracher, A.; Hausch, F. Large FK506-binding proteins shape the pharmacology of rapamycin. Mol. Cell. Biol. 2013, 33, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- Discovery Studio Visualizer; v16.1.0.15350; Dassault Systèmes BIOVIA: San Diego, CA, USA, 2016.
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein–ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.; Nguyen, T.H.; Pham, T.N.H.; Huy, N.T.; Bay, M.V.; Pham, M.Q.; Nam, P.C.; Vu, V.V.; Ngo, S.T. Autodock vina adopts more accurate binding poses but autodock4 forms better binding affinity. J. Chem. Inf. Model. 2020, 60, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Vieira, T.F.; Sousa, S.F. Comparing AutoDock and Vina in Ligand/Decoy Discrimination for Virtual Screening. Appl. Sci. 2019, 9, 4538. [Google Scholar] [CrossRef] [Green Version]
- Ala, A.A.; Olotu, B.B.; Ohia, C.M.D. Assessment of cytotoxicity of leaf extracts of Andrographis paniculata and Aspilia africana on murine cells in vitro. Arch. Basic Appl. Med. 2018, 6, 61–65. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds (asiaticoside and asiatic acid) are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligands | Docking Score (kcal/mol) | Hydrogen Bond (s) | Hydrophobic Interactions |
|---|---|---|---|
| Everolimus | −11.86 | Ser2035 | Tyr57, Phe67, Phe77, Trp90, Phe130, Trp2101, Tyr2038, Phe2039, Tyr2105 |
| Asiaticoside | −11.98 | Asp68, Val86, Phe2039 | Phe77, Tyr113, Phe2039, Trp2101, Tyr2105, Phe2108 |
| Asiatic acid | −10.37 | Asp68 | Val86, Ile87, Phe77, Phe2039, Trp2101, Tyr2105, Phe2108 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulkipli, N.N.; Zakaria, R.; Long, I.; Abdullah, S.F.; Muhammad, E.F.; Wahab, H.A.; Sasongko, T.H. In Silico Analyses and Cytotoxicity Study of Asiaticoside and Asiatic Acid from Malaysian Plant as Potential mTOR Inhibitors. Molecules 2020, 25, 3991. https://doi.org/10.3390/molecules25173991
Zulkipli NN, Zakaria R, Long I, Abdullah SF, Muhammad EF, Wahab HA, Sasongko TH. In Silico Analyses and Cytotoxicity Study of Asiaticoside and Asiatic Acid from Malaysian Plant as Potential mTOR Inhibitors. Molecules. 2020; 25(17):3991. https://doi.org/10.3390/molecules25173991
Chicago/Turabian StyleZulkipli, Ninie Nadia, Rahimah Zakaria, Idris Long, Siti Fadilah Abdullah, Erma Fatiha Muhammad, Habibah A. Wahab, and Teguh Haryo Sasongko. 2020. "In Silico Analyses and Cytotoxicity Study of Asiaticoside and Asiatic Acid from Malaysian Plant as Potential mTOR Inhibitors" Molecules 25, no. 17: 3991. https://doi.org/10.3390/molecules25173991
APA StyleZulkipli, N. N., Zakaria, R., Long, I., Abdullah, S. F., Muhammad, E. F., Wahab, H. A., & Sasongko, T. H. (2020). In Silico Analyses and Cytotoxicity Study of Asiaticoside and Asiatic Acid from Malaysian Plant as Potential mTOR Inhibitors. Molecules, 25(17), 3991. https://doi.org/10.3390/molecules25173991