Biofilms in Diabetic Foot Ulcers: Significance and Clinical Relevance

 , and
, and 
Abstract
:1. Introduction
2. Pathophysiology of Diabetic Foot Ulcers
2.1. Main Host-Related Factors
- ▪
- Diabetic immunopathy: Diabetic patients have an altered function of polymorphonuclear cells and impaired phagocytosis, chemotaxis, and bactericidal activity (related to both non oxidative and oxidative mechanisms), which are more evident in the presence of high hyperglycemia [8]. A study on diabetic mice showed that persistent hyperglycemia had a deleterious effect on the innate immunity and could lead to skin and soft tissue infections by Staphylococcus aureus [9].
- ▪
- Diabetic neuropathy: Neuropathy by C-fiber and autonomic nerve fiber dysfunction is a common and frequent complication of diabetes mellitus. An evolution of the deregulation of glycemic balance is the inhibition of nociception and the perception of pain, a process called loss of protective sensation [10]. Thus, patients may not initially notice small wounds in the legs and feet, and may fail to prevent infection. Studies have observed a reduction in foot skin innervation and the expression of neurogenic factors in DFU, correlated with low inflammatory cell accumulation and therefore in the chronicity of DFU. This contributes to enhancing susceptibility to infection of diabetic neuropathic foot ulcers [11].
- ▪
- Diabetic angiopathy: Peripheral arterial disease (PAD) and microangiopathy are the main risk factors for DFU. The decrease in the oxygenation of tissues by thickening the capillary basement membrane is a hallmark of diabetic angiopathy [12]. Disease of arteries in the lower limb is a well-known risk factor for DFU. Indeed, studies have shown that PAD presents a 5.5-fold increased risk for DFU [13]. The ischemia caused by the angiopathy also enhances the severity of the infection as a result of a poor delivery of oxygen and nutrients in the infected wound and because of poor antibiotic tissue penetration [14].
2.2. DFU Microbiota
2.3. Disturbances in the Host–Microorganism Interplay
- ▪
- Bacterial virulence: The virulence of pathogens is a key element in the pathophysiology of DFU. The ability of a bacterium to be virulent is key to the precarious balance between colonization and infection [26]. Bacterial virulence has been characterized using DNA microarray-based genotyping, multiplex polymerase chain reaction (PCR), and in vivo assays [26,27]. Among the large panel of virulence factors, bacterial proteases (serine-, cysteine-, and metallo-proteases), produced by a wide range of pathogenic bacteria, could play a major role in the pathogenesis of wound healing [28]. However, these wounds, and especially DFU, are highly polymicrobial, and bacterial interactions should also be studied in order to better understand the mechanisms of infection and the role of each of the pathogens involved in DFU.
- ▪
- Biofilm organization: In a 2008 study assessing wound tissue biopsies using electron microscopy, James et al. suggested that 60% of chronic wounds present biofilms versus 6% for acute wounds [29]. In the following sections of this review, we focus on the formation of biofilms, evidence of biofilms in DFU, influence of the diabetic environment, and finally the clinical implications of biofilms in DFU.
3. Overview of Biofilms in DFU
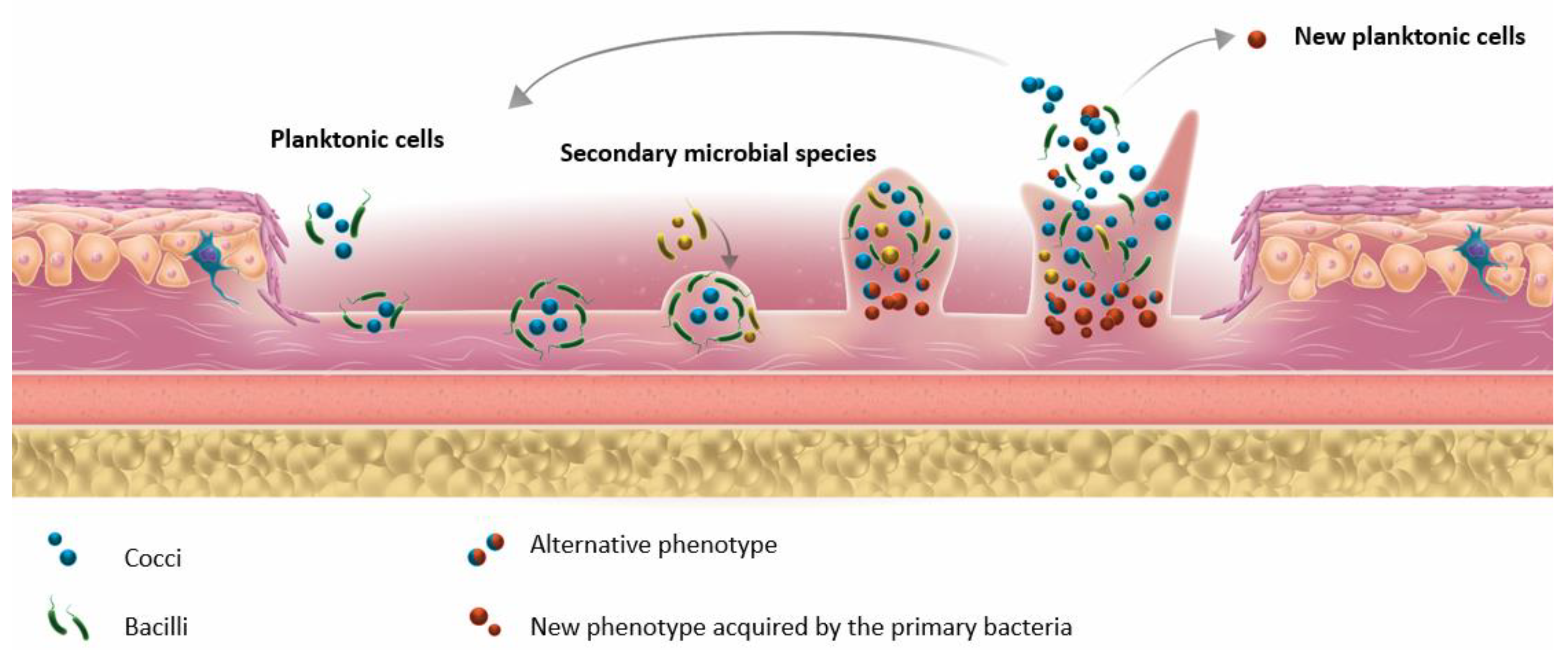
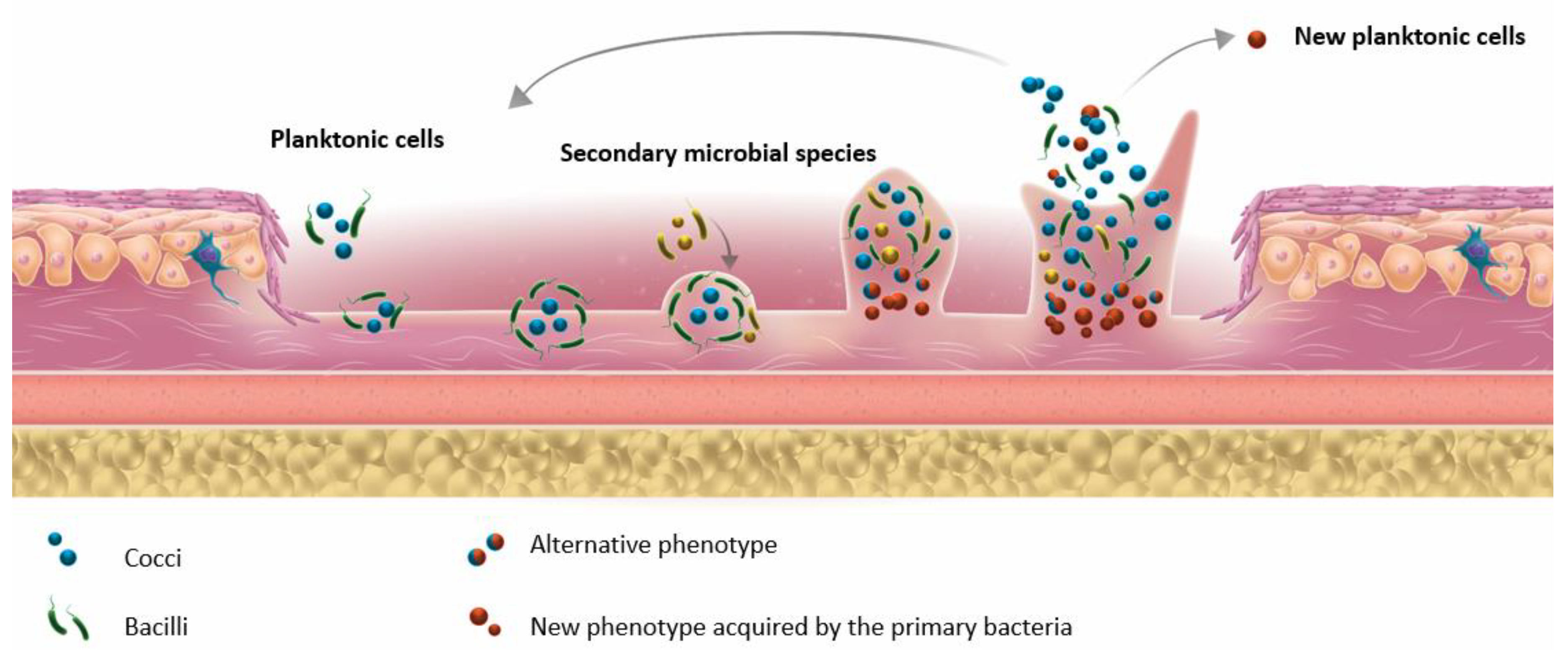
3.1. Biofilm Formation in DFU and Tools for Detection
- ▪
- ▪
- ▪
- The concept of FEP was proposed by Dowd et al. after observing that different bacterial species can collaborate and interact with each other. FEPs are responsible for the chronicity of infection and for the maintenance of the pathogenic biofilm [7].
3.2. Biofilm Studies in Animal Models of DFU
3.3. Biofilm Studies in Human Clinical DFU
3.4. Factors Influencing Biofilm Formation in DFU
- -
- -
- Increased S. aureus population [15], particularly in neuropathic DFUs [61]. However, their microbiota present a similar level of richness (number of different species in the wound community), abundance, and diversity compared to other chronic wounds [63], suggesting that the microbiota is not influenced by the wound type.
- -
- The wound depth with a more diverse and complex microbiota in the deep part of the wound [64] where pathogenic, particularly anaerobic, bacteria are sheltered.
- -
- Environmental factors (e.g., demographic characteristics, personal hygiene, geographical location of the patient, high glycemic level, and previous exposure to antimicrobial therapy) [65].
- -
- Patient immune status that modifies the role of low-virulence bacteria (e.g., Staphylococcus sp. and corynebacteria) towards a higher pathogenicity [66], and where excessive secretion of pro-inflammatory cytokines, pH, temperature, or antimicrobial treatment (topic or systemic administration) [67] can increase tissue destruction [68].
- -
- DFU duration is positively correlated with the ecological diversity of the bacteria present in the wounds, species richness, and relative abundance of Proteobacteria. It is also negatively correlated with the relative abundance of staphylococci [69].
- -
- -
- The development of a “unique microbiota” in each DFU (new or recurrent) [17].
3.5. Bacterial Organization Inside DFU
4. Clinical Impact of Biofilms in DFU
4.1. Antibiotics Resistance
4.2. Host Immune Response
5. Therapeutic Perspectives
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic foot ulcers and their recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef]
- Prompers, L.; Schaper, N.; Apelqvist, J.; Edmonds, M.; Jude, E.; Mauricio, D.; Uccioli, L.; Urbancic, V.; Bakker, K.; Holstein, P.; et al. Prediction of outcome in individuals with diabetic foot ulcers: Focus on the differences between individuals with and without peripheral arterial disease. The EURODIALE Study. Diabetologia 2008, 51, 747–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, K.; Apelqvist, J.; Lipsky, B.A.; Van Netten, J.J.; International Working Group on the Diabetic Foot. The 2015 IWGDF guidance documents on prevention and management of foot problems in diabetes: Development of an evidence-based global consensus. Diabetes Metab. Res. Rev. 2016, 32, 2–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, P.J.; Melton, L.J.I. Diabetes in America: Diabetes Data Compiled 1984; Peripheral vascular disease and diabetes; Government Printing Office: Washington, DC, USA, 1985. [Google Scholar]
- Adler, A.I.; Boyko, E.J.; Ahroni, J.H.; Smith, D.G. Lower-extremity amputation in diabetes. The independent effects of peripheral vascular disease, sensory neuropathy, and foot ulcers. Diabetes Care 1999, 22, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Ndosi, M.; Wright-Hughes, A.; Brown, S.; Backhouse, M.; Lipsky, B.A.; Bhogal, M.; Reynolds, C.; Vowden, P.; Jude, E.B.; Nixon, J.; et al. Prognosis of the infected diabetic foot ulcer: A 12-month prospective observational study. Diabet. Med. 2018, 35, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Dowd, S.E.; Wolcott, R.D.; Sun, Y.; McKeehan, T.; Smith, E.; Rhoads, D. Polymicrobial nature of chronic diabetic foot ulcer biofilm infections determined using bacterial tag encoded FLX amplicon pyrosequencing (bTEFAP). PLoS ONE 2008, 3, e3326. [Google Scholar] [CrossRef] [PubMed]
- Lecube, A.; Pachón, G.; Petriz, J.; Hernández, C.; Simó, R. Phagocytic activity is impaired in type 2 diabetes mellitus and increases after metabolic improvement. PLoS ONE 2011, 6, e23366. [Google Scholar] [CrossRef]
- Park, S.; Rich, J.; Hanses, F.; Lee, J.C. Defects in innate immunity predispose C57BL/6J-Leprdb/Leprdb mice to infection by Staphylococcus aureus. Infect. Immun. 2009, 77, 1008–1014. [Google Scholar] [CrossRef] [Green Version]
- Yagihashi, S.; Mizukami, H.; Sugimoto, K. Mechanism of diabetic neuropathy: Where are we now and where to go? J. Diabetes Investig. 2011, 2, 18–32. [Google Scholar] [CrossRef] [Green Version]
- Galkowska, H.; Olszewski, W.L.; Wojewodzka, U.; Rosinski, G.; Karnafel, W. Neurogenic factors in the impaired healing of diabetic foot ulcers. J. Surg. Res. 2006, 134, 252–258. [Google Scholar] [CrossRef]
- Chawla, A.; Chawla, R.; Jaggi, S. Microvasular and macrovascular complications in diabetes mellitus: Distinct or continuum? Indian J. Endocrinol. Metab. 2016, 20, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.J.; Lipsky, B.A.; Berendt, A.R.; Embil, J.M.; Lavery, L.A.; Senneville, E.; Urbančič-Rovan, V.; Bakker, K.; Jeffcoate, W.J. A systematic review of the effectiveness of interventions in the management of infection in the diabetic foot. Diabetes Metab. Res. Rev. 2012, 28, 142–162. [Google Scholar] [CrossRef] [PubMed]
- Raymakers, J.T.; Houben, A.J.; van der Heyden, J.J.; Tordoir, J.H.; Kitslaar, P.J.; Schaper, N.C. The effect of diabetes and severe ischaemia on the penetration of ceftazidime into tissues of the limb. Diabet. Med. 2001, 18, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Redel, H.; Gao, Z.; Li, H.; Alekseyenko, A.V.; Zhou, Y.; Perez-Perez, G.I.; Weinstock, G.; Sodergren, E.; Blaser, M.J. Quantitation and composition of cutaneous microbiota in diabetic and nondiabetic men. J. Infect. Dis. 2013, 207, 1105–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontcharova, V.; Youn, E.; Sun, Y.; Wolcott, R.D.; Dowd, S.E. A comparison of bacterial composition in diabetic ulcers and contralateral intact skin. Open Microbiol. J. 2010, 4, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Jneid, J.; Lavigne, J.P.; La Scola, B.; Cassir, N. The diabetic foot microbiota: A review. Hum. Microbiome J. 2017, 5–6, 1–6. [Google Scholar] [CrossRef]
- Pereira, S.G.; Moura, J.; Carvalho, E.; Empadinhas, N. Microbiota of Chronic Diabetic Wounds: Ecology, Impact, and Potential for Innovative Treatment Strategies. Front. Microbiol. 2017, 8, 1791. [Google Scholar] [CrossRef]
- Gardiner, M.; Vicaretti, M.; Sparks, J.; Bansal, S.; Bush, S.; Liu, M.; Darling, A.; Harry, E.; Burke, C.M. A longitudinal study of the diabetic skin and wound microbiome. PeerJ 2017, 5, e3543. [Google Scholar] [CrossRef]
- Malone, M.; Johani, K.; Jensen, S.O.; Gosbell, I.B.; Dickson, H.G.; Hu, H.; Vickery, K. Next Generation DNA Sequencing of Tissues from Infected Diabetic Foot Ulcers. EBioMedicine 2017, 21, 142–149. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, A.; Brodell, J.D.; Daiss, J.L.; Schwarz, E.M.; Oh, I. Evidence of differential microbiomes in healing versus non-healing diabetic foot ulcers prior to and following foot salvage therapy. J. Orthop. Res. 2019, 37, 1596–1603. [Google Scholar] [CrossRef]
- Verbanic, S.; Shen, Y.; Lee, J.; Deacon, J.M.; Chen, I.A. Microbial predictors of healing and short- term effect of debridement on the microbiome of chronic wounds. NPJ Biofilms Microbiomes 2020, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Sloan, T.J.; Turton, J.C.; Tyson, J.; Musgrove, A.; Fleming, V.M.; Lister, M.M.; Loose, M.W.; Sockett, R.E.; Diggle, M.; Game, F.L.; et al. Examining diabetic heel ulcers through an ecological lens: Microbial community dynamics associated with healing and infection. J. Med. Microbiol. 2019, 68, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Loesche, M.; Gardner, S.E.; Kalan, L.; Horwinski, J.; Zheng, Q.; Hodkinson, B.P.; Tyldsley, A.S.; Franciscus, C.L.; Hillis, S.I.; Mehta, S.; et al. Temporal Stability in Chronic Wound Microbiota is Associated with Poor Healing. J. Investig. Dermatol. 2017, 137, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalan, L.R.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Uberoi, A.; Gardner, S.E.; Grice, E.A. Strain- and Species- Level Variation in the Microbiome of Diabetic Wounds Is Associated with Clinical Outcomes and Therapeutic Efficacy. Cell Host Microbe 2019, 25, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Spichler, A.; Hurwitz, B.L.; Armstrong, D.G.; Lipsky, B.A. Microbiology of diabetic foot infections: From Louis Pasteur to ‘crime scene investigation’. BMC Med. 2015, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavigne, J.P.; Sotto, A.; Dunyach-Remy, C.; Lipsky, B.A. New molecular techniques to study the skin microbiota of diabetic foot ulcers. Adv. Wound Care 2015, 4, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, S.M.; Cochrane, C.A.; Clegg, P.D.; Percival, S.L. The Role of Endogenous and Exogenous Enzymes in Chronic Wounds: A Focus on the Implications of Aberrant Levels of Both Host and Bacterial Proteases in Wound Healing. Wound Repair Regen. 2012, 20, 125–136. [Google Scholar] [CrossRef]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Percival, S.L.; McCarty, S.M.; Lipsky, B. Biofilms and Wounds: An Overview of the Evidence. Adv. Wound Care 2015, 4, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. APMIS 2013, 121, 1–58. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.L.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Holá, V.; Imbert, C.; Kirketerp-Møller, K.; et al. ESCMID guideline for the diagnosis and treatment of biofilm infections 2014. Clin. Microbiol. Infect. 2015, 21, S1–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asfour, H. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Guilhen, C.; Forestier, C.; Balestrino, D. Biofilm dispersal: Multiple elaborate strategies for dissemination of bacteria with unique properties. Mol. Microbiol. 2017, 105, 188–210. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Usui, M.L.; Underwood, R.A.; Singh, P.K.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Time course study of delayed wound healing in a biofilm challenged diabetic mouse model. Wound Repair Regen. 2012, 20, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Oglesby-Sherrouse, A.G. Interactions between Pseudomonas aeruginosa and Staphylococcus aureus during co-cultivations and polymicrobial infections. Appl. Microbiol. Biotechnol. 2016, 100, 6141–6148. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Shu, J.C.; Lin, M.H.; Chong, K.Y.; Chen, C.C.; Wen, S.M.; Hsieh, Y.T.; Lia, W.T. High Glucose concentration promotes vancomycin-enhanced biofilm formation of vancomycin-non-susceptible Staphylococcus aureus in diabetic mice. PLoS ONE 2015, 10, e0134852. [Google Scholar] [CrossRef]
- Nouvong, A.; Ambrus, A.M.; Zhang, E.R.; Hultman, L.; Coller, H.A. Reactive oxygen species and bacterial biofilms in diabetic wound healing. Physiol. Genom. 2016, 48, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Dhall, S.; Do, D.C.; Garcia, M.; Kim, J.; Mirebrahim, S.H.; Lyubovitsky, J.; Lonardi, S.; Nothnagel, E.A.; Schiller, N.; Martins-Green, M. Generating and reversing chronic wounds in diabetic mice by manipulating wound redox parameters. J. Diabetes Res. 2014, 2014, 1–18. [Google Scholar] [CrossRef] [Green Version]
- James, G.A.; Ge Zhao, A.; Usui, M.; Underwood, R.A.; Nguyen, H.; Beyenal, H.; Pulcini, E.D.; Agostino Hunt, A.; Bernstein, H.C.; Fleckman, P.; et al. Microsensor and transcriptomic signatures of oxygen depletion in biofilms associated with chronic wounds. Wound Repair Regen. 2016, 24, 373–383. [Google Scholar] [CrossRef]
- Agostinho Hunt, A.M.; Gibson, J.A.; Larrivee, C.L.; O’Reilly, S.; Navitskaya, S.; Needle, D.B.; Abramowitch, R.B.; Busik, J.V.; Waters, C.M. A bioluminescent Pseudomonas aeruginosa wound model reveals increased mortality of type 1 diabetic mice to biofilm infection. J. Wound Care 2017, 26, S24–S33. [Google Scholar] [PubMed]
- Goldufsky, J.; Wood, S.J.; Jayaraman, V.; Majdobeh, O.; Chen, L.; Qin, S.; Zhang, C.; DiPietro, L.A.; Shafikhani, S.H. Pseudomonas aeruginosa uses T3SS to inhibit diabetic wound healing. Wound Repair Regen. 2015, 23, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neut, D.; Tijdens-Creusen, E.J.; Bulstra, S.K.; van der Mei, H.C.; Busscher, H.J. Biofilms in chronic diabetic foot ulcers—A study of 2 cases. Acta Orthop. 2011, 82, 383–385. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.; Mohammad, Z.; Ahmad, J. The diabetic foot infections: Biofilms and antimicrobial resistance. Diabetes Metab. Syndr. 2013, 7, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Murali, T.S.; Kavitha, S.; Spoorthi, J.; Bhat, D.V.; Prasat, A.S.B.; Upton, Z.; Ramachandra, L.; Acharya, R.V.; Satyamoorthy, K. Characteristics of microbial drug resistance and its correlates in chronic diabetic foot ulcer infections. J. Med. Microbiol. 2014, 63, 1377–1385. [Google Scholar] [CrossRef]
- Banu, A.; Noorul Hassan, M.M.; Rajkumar, J.; Srinivasa, S. Spectrum of bacteria associated with diabetic foot ulcer and biofilm formation: A prospective study. Australas. Med. J. 2015, 8, 280–285. [Google Scholar] [CrossRef]
- Oates, A.; Bowling, F.L.; Boulton, A.J.; Bowler, P.G.; Metcalf, D.G.; McBain, A.J. The visualization of biofilms in chronic diabetic foot wounds using routine diagnostic microscopy methods. J. Diabetes Res. 2014, 2014, 153586. [Google Scholar] [CrossRef]
- Zhao, G.; Usui, M.L.; Lippman, S.I.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Biofilms and Inflammation in Chronic Wounds. Adv. Wound Care 2013, 2, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial interactions: Impact on pathogenesis and human disease. Clin. Microbiol. Rev. 2012, 25, 193–213. [Google Scholar] [CrossRef] [Green Version]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef] [PubMed]
- Mottola, C.; Semedo-Lemsaddek, T.; Mendes, J.J.; Melo-Cristino, J.; Tavares, L.; Cavaco-Silva, P.; Oliveira, M. Molecular typing, virulence traits and antimicrobial resistance of diabetic foot staphylococci. J. Biomed. Sci 2016, 23, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The microbial “protective clothing’ in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Banerjee, T.; Chakravarty, J.; Singh, S.K.; Dwivedi, A.; Tilak, R. Identification, antifungal resistance profile, in vitro biofilm formation and ultrastructural characteristics of Candida species isolated from diabetic foot patients in Northern India. Indian J. Med. Microbiol. 2016, 34, 308–314. [Google Scholar] [PubMed]
- Sugimoto, S.; Iwamoto, T.; Takada, K.; Okuda, K.I.; Tajima, A.; Iwase, T.; Mizunoe, Y. Staphylococcus epidermidis Esp degrades specific proteins associated with Staphylococcus aureus biofilm formation and host-pathogen interaction. J. Bacteriol. 2013, 195, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Philips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the chronic wound microbiota of 2,963 patients by 16S rDNA pyrosequencing. Wound Repair Regen. 2016, 24, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Jneid, J.; Cassir, N.; Schuldiner, S.; Jourdan, N.; Sotto, A.; Lavigne, J.P.; La Scola, B. Exploring the microbiota of diabetic foot infections with culturomics. Front. Cell Infect. Microbiol. 2018, 8, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound Microbiology and Associated Approaches to Wound Management. Clin. Microbiol. Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef] [Green Version]
- Oates, A.; Bowling, F.L.; Boulton, A.J.M.; McBain, A.J. Molecular and culture-based assessment of the microbial diversity of diabetic chronic foot wounds and contralateral skin sites. J. Clin. Microbiol. 2012, 50, 2263–2271. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.E.; Hillis, S.L.; Heilmann, K.; Segre, J.A.; Grice, E.A. The neuropathic diabetic foot ulcer microbiome is associated with clinical factors. Diabetes 2013, 62, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Percival, S.L.; Malone, M.; Mayer, D.; Salisbury, A.M.; Schultz, G. Role of anaerobes in polymicrobial communities and biofilms complicating diabetic foot ulcers. Int. Wound J. 2018, 15, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.R.; Gómez, B.I.; McIntyre, M.K.; Dubick, M.A.; Christy, R.J.; Nicholson, S.E.; Burmeister, D.M. The Cutaneous Microbiome and Wounds: New Molecular Targets to Promote Wound Healing. Int. J. Mol. Sci. 2018, 19, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipsky, B.A.; Richard, J.L.; Lavigne, J.P. Diabetic foot ulcer microbiome: One small step for molecular microbiology... One giant leap for understanding diabetic foot ulcers? Diabetes 2013, 62, 679–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percival, S.L.; Thomas, J.G.; Williams, D.W. Biofilms and bacterial imbalances in chronic wounds: Anti-Koch. Int. Wound J. 2010, 7, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Collier, A.; Townsend, E.M.; O’Donnell, L.E.; Bal, A.M.; Butcher, J.; MacKay, W.G.; Ramage, G.; Williams, C. One step closer to understanding the role of bacteria in diabetic foot ulcers: Characterising the microbiome of ulcers. BMC Microbiol. 2016, 16, 54. [Google Scholar] [CrossRef] [Green Version]
- Rahim, K.; Saleha, S.; Zhu, X.; Huo, L.; Basit, A.; Franco, O.L. Bacterial contribution in chronicity of wounds. Microb. Ecol. 2017, 73, 710–721. [Google Scholar] [CrossRef]
- Patel, S.; Srivastava, S.; Singh, M.R.; Singh, D. Mechanistic insight into diabetic wounds: Pathogenesis, molecular targets and treatment strategies to pace wound healing. Biomed. Pharm. 2019, 112, 108615. [Google Scholar] [CrossRef]
- Cogen, A.L.; Nizet, V.; Gallo, R.L. Skin microbiota: A source of disease or defence? Br. J. Dermatol. 2008, 158, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Fazli, M.; Bjarnsholt, T.; Kirketerp-Møller, K.; Jørgensen, B.; Andersen, A.S.; Krogfelt, K.A.; Givskov, M.; Tolker-Nielsen, T. Nonrandom distribution of Pseudomonas aeruginosa and Staphylococcus aureus in chronic wounds. J. Clin. Microbiol. 2009, 47, 4084–4089. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, L.R.; Deziel, E.; D’Argenio, D.A.; Lépine, F.; Emerson, J.; McNamara, S.; Gibson, R.L.; Ramsey, B.W.; Miller, S.I. Selection for Staphylococcus aureus small-colony variants due to growth in the presence of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2006, 103, 19890–19895. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.G.; Liu, Y.C.; Chang, C.Y. Inhibiting N-acyl-homoserine lactone synthesis and quenching Pseudomonas quinolone quorum sensing to attenuate virulence. Front. Microbiol. 2015, 6, 1173. [Google Scholar] [CrossRef] [PubMed]
- Schurr, M.J. Pseudomonas aeruginosa alginate benefits Staphylococcus aureus? J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.E.; Brown, D.G.; Limoli, D.H.; Phelan, V.V.; O’Toole, G.A. Exogenous alginate protects Staphylococcus aureus from killing by Pseudomonas aeruginosa. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, K.J.; Burd, T.A.; Anglen, J.O.; Simpson, A.W.; Christensen, G.D.; Gainor, B.J. Synergy between Staphylococcus aureus and Pseudomonas aeruginosa in a rat model of complex orthopaedic wounds. J. Bone Joint Surg. 2001, 83, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Hotterbeekx, A.; Kumar-Singh, S.; Goossens, H.; Malhotra-Kumar, S. In vivo and in vitro interactions between Pseudomonas aeruginosa and Staphylococcus spp. Front. Cell Infect. Microbiol. 2017, 7, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an in vitro wound model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef] [Green Version]
- Mastropaolo, M.D.; Evans, N.P.; Byrnes, M.K.; Stevens, A.M.; Robertson, J.L.; Melville, S.B. Synergy in polymicrobial infections in a mouse model of Type 2 diabetes. Infect. Immun. 2005, 73, 6055–6063. [Google Scholar] [CrossRef] [Green Version]
- Nair, N.; Biswas, R.; Gotz, F.; Biswas, L. Impact of Staphylococcus aureus on pathogenesis in polymicrobial infections. Infect. Immun. 2014, 82, 2162–2169. [Google Scholar] [CrossRef] [Green Version]
- Ngba Essebe, C.; Visvikis, O.; Fines-Guyon, M.; Vergne, A.; Cattoir, V.; Lecoustumier, A.; Lemichez, E.; Sotto, A.; Lavigne, J.P.; Dunyach-Remy, C. Decrease of Staphylococcus aureus virulence by Helcococcus kunzii in a Caenorhabditis elegans model. Front. Cell Infect. Microbiol. 2017, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.W.; Freire, M.O.; Gabrilska, R.A.; Rumbaugh, K.P.; Lemon, K.P. Staphylococcus aureus shifts toward commensalism in response to Corynebacterium Species. Front. Microbiol. 2016, 7, 1230. [Google Scholar] [CrossRef] [Green Version]
- Vandecandelaere, I.; Depuydt, P.; Nelis, H.J.; Coenye, T. Protease production by Staphylococcus epidermidis and its effect on Staphylococcus aureus biofilms. Pathog. Dis. 2014, 70, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, T.; Kokubu, E.; Kawana, T.; Saito, A.; Okuda, K.; Ishihara, K. Synergy in biofilm formation between Fusobacterium nucleatum and Prevotella species. Anaerobe 2012, 18, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottola, C.; Mendes, J.J.; Cristino, J.M.; Cavasco-Silva, P.; Tavares, L.; Oliveira, M. Polymicrobial biofilms by diabetic foot clinical isolates. Folia Microbiol. 2016, 61, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Lynch, A.S.; Robertson, G.T. Bacterial and fungal biofilm infections. Annu. Rev. Med. 2008, 59, 415–428. [Google Scholar] [CrossRef]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; de Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarćon, K.P.; Melo, W.C.M.A.; Marcelino, M.Y.; Braz, J.D.; et al. Fungal Biofilms and Polymicrobial Diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the chronic-wound microbiome: Fungal communities are prevalent, dynamic, and associated with delayed healing. mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Malone, M.; Bjarnsholt, T.; McBain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The prevalence of biofilms in chronic wounds: A systematic review and meta-analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghigo, J.M. Natural conjugative plasmids induce bacterial biofilm development. Nature 2001, 412, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological implications. Microb. Cell Fact. 2016, 15, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebreyohannes, G.; Nyerere, C.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganism. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suller, M.T.E.; Lloyd, D. The antibacterial activity of vancomycin towards Staphylococcus aureus under aerobic and anaerobic conditions. J. Appl. Microbiol. 2002, 92, 866–872. [Google Scholar] [CrossRef] [Green Version]
- González, J.F.; Hahn, M.M.; Gunn, J.S. Chronic biofilm-based infections: Skewing of the immune response. Pathog. Dis. 2018, 76, fty023. [Google Scholar] [CrossRef]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef]
- Castillo-Juárez, I.; Maeda, T.; Mandujano-Tinoco, E.A.; Tomás, M.; Pérez-Eretza, B.; García-Contreras, S.J.; Wood, T.K.; García-Contreras, R. Role of quorum sensing in bacterial infections. World J. Clin. Cases 2015, 3, 575–598. [Google Scholar] [CrossRef]
- Hirschfeld, J. Dynamic interactions of neutrophils and biofilms. J. Oral Microbiol. 2014, 6, 26102. [Google Scholar] [CrossRef]
- Wolska, K.I.; Grudniak, A.M.; Rudnicka, Z.; Markowska, K. Genetic control of bacterial biofilms. J. Appl. Genet. 2016, 57, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Moser, C.; Pedersen, H.T.; Lerche, C.J.; Kolpen, M.; Line, L.; Thomsen, K.; Høiby, N.; Jensen, P.O. Biofilms and host response—Helpful or harmful. APMIS 2017, 125, 320–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinton, A.; Carter, T. Chronic Wound Biofilms: Pathogenesis and Potential Therapies. Lab. Med. 2015, 46, 277–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipsky, B.A.; Senneville, E.; Abbas, Z.G.; Aragón-Sánchez, J.; Diggle, M.; Embil, J.M.; Kono, S.; Lavery, L.A.; Malone, M.; van Asten, S.A.; et al. Guideline on the diagnostic and treatment of foot infection in persons with diabetes (IWGDF 2019 update). Diabetes Metab. Res. Rev. 2020, 36, e3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolcott, R.D.; Kennedy, J.P.; Dowd, S.E. Regular debridement is the main tool for maintaining a healthy wound bed in most chronic wounds. J. Wound Care 2009, 18, 54–56. [Google Scholar] [CrossRef] [Green Version]
- Lázaro-Martinez, J.L.; Alvaro-Afonso, F.J.; Garcia-Alvarez, Y.; Molines-Barroso, R.J.; García-Morales, E.; Sevillano-Fernández, D. Ultrasound-assisted debridement of neuroischaemic diabetic foot ulcers, clinical and microbiological effects: A case series. J. Wound Care 2018, 27, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Raad, I.I.; Fang, X.; Keutgen, X.M.; Jiang, Y.; Sherertz, R.; Hachem, R. The role of chelators in preventing biofilm formation and catheter-related bloodstream infections. Curr. Opin. Infect. Dis. 2008, 21, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Li, W.; Li, C.; Vittayapadung, S.; Lin, L. Liposome containing cinnamon oil with antibacterial activity against methicillin-resistant Staphylococcus aureus biofilm. Biofouling 2016, 32, 215–225. [Google Scholar] [CrossRef]
- Kim, S.G.; Yoon, Y.H.; Choi, J.W.; Rha, K.S.; Park, Y.H. Effect of furanone on experimentally induced Pseudomonas aeruginosa biofilm formation: In vitro study. Int. J. Pediatr. Otorhinolaryngol. 2012, 76, 1575–1578. [Google Scholar] [CrossRef]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef]
- Coraça-Huber, D.C.; Dichtl, S.; Steixner, S.; Nogler, M.; Weiss, G. Iron chelation destabilizes bacterial biofilms and potentiates the antimicrobial activity of antibiotics against coagulase-negative Staphylococci. Pathog. Dis. 2018, 76, fty052. [Google Scholar] [CrossRef]
- Kalpana, B.J.; Aarthy, S.; Pandian, S.K. Antibiofilm activity of α-amylase from Bacillus subtilis S8–18 against biofilm forming human bacterial pathogens. Appl. Biochem. Biotechnol. 2012, 167, 1778–1794. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.A.; Huigens, R.W., III; Cavanagh, J.; Melander, C. Synergistic effects between conventional antibiotics and 2-aminoimidazole-derived antibiofilm agents. Antimicrob. Agents Chemother. 2010, 54, 2112–2118. [Google Scholar] [CrossRef] [Green Version]
- Chung, P.Y.; Toh, Y.S. Anti-biofilm agents: Recent breakthrough against multi-drug resistant Staphylococcus aureus. Pathog. Dis. 2014, 70, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardivo, J.P.; Adami, F.; Correa, J.A.; Pinhal, M.A.S.; Baptista, M.S. A clinical trial testing the efficacy of PDT in preventing amputation in diabetic patients. Photodiagnosis Photodyn. Ther. 2014, 11, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Taha, O.A.; Connerton, P.L.; Connerton, I.F.; El-Shibiny, A. Bacteriophage ZCKP1: A potential treatment for Klebsiella pneumoniae isolated from diabetic foot patients. Front. Microbiol. 2018, 9, 2127. [Google Scholar] [CrossRef] [PubMed]
- Fish, R.; Kutter, E.; Bryan, D.; Wheat, G.; Kuhl, S. Resolving digital staphylococcal osteomyelitis using bacteriophage—A case report. Antibiotics 2018, 7, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, M.; Adibhesami, M. The effect of silver nanoparticles on wounds contaminated with Pseudomonas aeruginosa in mice: An experimental study. Iran. J. Pharm. Res. 2017, 16, 661–669. [Google Scholar]
- Thombare, N.; Jha, U.; Mishra, S.; Siddiqui, M.Z. Guar gum as a promising starting material for diverse applications: A review. Int. J. Biol. Macromol. 2016, 88, 361–372. [Google Scholar] [CrossRef]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Kamysz, W.; Orlando, F.; Mocchegiani, F.; Silvestri, C.; Licci, A.; Chiodi, L.; Lukasiak, J.; et al. Citropin 1.1-treated central venous catheters improve the efficacy of hydrophobic antibiotics in the treatment of experimental staphylococcal catheter-related infection. Peptides 2006, 27, 1210–1216. [Google Scholar] [CrossRef]
- Dutta, P.; Das, S. Mammalian antimicrobial peptides: Promising therapeutic targets against infection and chronic inflammation. Curr. Top. Med. Chem. 2016, 16, 99–129. [Google Scholar] [CrossRef]
- Bilyayeva, O.O.; Neshta, V.V.; Golub, A.A.; Sams-Dodd, F. Comparative clinical study of the wound healing effects of a novel micropore particle technology: Effects on wounds, venous leg ulcers, and diabetic foot ulcers. Wounds 2017, 29, 1–9. [Google Scholar] [PubMed]
- Minden-Bjrkenmajer, B.; Bowlin, G. Honey-based templates in wound healing and tissue engineering. Bioengineering 2018, 5, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |

{kind=link}
| Animal Model | Strain Used | Findings | Reference |
|---|---|---|---|
| db/db mice | P. aeruginosa (PAO1) | Biofilm evidence after a 6-mm punch biopsy wound on the dorsal skin | [36] |
| db/db mice | P. aeruginosa (PAO1) | Biofilm delays wound healing | [37] |
| TallyHo mice (Type 2 diabetes mellitus) | P. aeruginosa | Biofilm decreases TLR 2, TLR 4, IL-1α, and TNF-α expression and neutrophil oxidative burst activity | [39] |
| BALB/c mice with injection of STZ | Vancomycin-resistant S. aureus | Correlation between glucose concentration and biofilm formation | [38] |
| db/db mice | Wound microbiome | Oxidative stress and ROS favor biofilm formation and establish a chronic wound | [40] |
| db/db mice | P. aeruginosa | Bacteria in biofilm induce oxygen stress by producing metabolites and recruiting defense cells that reduce oxygen | [41] |
| Mice with injection of STZ | P. aeruginosa | Biofilm increases wound depth, mortality rate, and pus production | [42] |
| db/db mice | P. aeruginosa | P. aeruginosa infection is independent of its ability to form biofilm and primarily depends on T3SS | [43] |
| Model | N° of Patients | Biofilm Visualization | Findings | Reference |
|---|---|---|---|---|
| DFU | 2 | CLSM | Evidence of biofilms | [44] |
| DFU | 162 | Microtiter plate assay | Biofilms in 67.9% of infected DFUs | [45] |
| DFU | 26 | FISH and ESEM | Observation of the formed biofilms and their bacterial constitution | [48] |
| DFU | 357 | Crystal violet | Observation of the formed biofilms | [46] |
| DFU | 100 | Congo Red dye, tissue culture plates, and crystal violet staining | Biofilm formation in 46.3% of isolates, predominantly by S. aureus (38.8% of isolates) and MDR bacteria (46.3%) | [47] |
| DFU | 49 | Calgary biofilm pin lid device with resazurin and PCR of genes associated with biofilm formation | Biofilms are resistant to antibiotics at concentrations 10–1000 times higher than those required to kill planktonic cells | [53] |
| DFU | 155 | Microtiter plate assay and ELISA, XTT formazan, and SEM | Presence and importance of non-Candida albicans species in biofilms | [55] |
| DFU | 95 | Microtiter plate assay and FISH | Polymicrobial biofilms are thicker | [56] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouget, C.; Dunyach-Remy, C.; Pantel, A.; Schuldiner, S.; Sotto, A.; Lavigne, J.-P. Biofilms in Diabetic Foot Ulcers: Significance and Clinical Relevance. Microorganisms 2020, 8, 1580. https://doi.org/10.3390/microorganisms8101580
Pouget C, Dunyach-Remy C, Pantel A, Schuldiner S, Sotto A, Lavigne J-P. Biofilms in Diabetic Foot Ulcers: Significance and Clinical Relevance. Microorganisms. 2020; 8(10):1580. https://doi.org/10.3390/microorganisms8101580
Chicago/Turabian StylePouget, Cassandra, Catherine Dunyach-Remy, Alix Pantel, Sophie Schuldiner, Albert Sotto, and Jean-Philippe Lavigne. 2020. "Biofilms in Diabetic Foot Ulcers: Significance and Clinical Relevance" Microorganisms 8, no. 10: 1580. https://doi.org/10.3390/microorganisms8101580
APA StylePouget, C., Dunyach-Remy, C., Pantel, A., Schuldiner, S., Sotto, A., & Lavigne, J.-P. (2020). Biofilms in Diabetic Foot Ulcers: Significance and Clinical Relevance. Microorganisms, 8(10), 1580. https://doi.org/10.3390/microorganisms8101580