Polygenetic-Risk Scores Related to Crystallin Metabolism Are Associated with Age-Related Cataract Formation and Interact with Hyperglycemia, Hypertension, Western-Style Diet, and Na Intake



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Criteria of ARC
2.3. Demographic, Anthropometric, and Biochemical Information
2.4. Food and Nutrient Intake Assessments
2.5. Genotyping Using a Korean Chip and Quality Control
2.6. Genetic Variants Influencing ARC Risk and the Best Model for Detecting Gene–Gene Interactions as Determined by Generalized Multifactor Dimensionality Reduction (GMDR)
2.7. Statistical Analyses
3. Results
3.1. Demographic, Anthropometric, and Biochemical Parameters According to Gender and Cataract Incidence
3.2. The Best SNP Model Selected from Genetic Variants Related to ARC by Logistic Regression Analysis
3.3. Association of the PRS of the Two Models with ARC Risk
3.4. Interaction between PRS and Metabolic Parameters and Lifestyles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shiels, A.; Hejtmancik, J.F. Mutations, and mechanisms in congenital and age-related cataracts. Exp. Eye Res. 2017, 156, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truscott, R.J.W.; Friedrich, M.G. Molecular Processes Implicated in Human Age-Related Nuclear Cataract. Investig. Ophthalmol. Vis. Sci. 2019, 60, 5007–5021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.H.; Wu, J.; Cho, E.; Ogata, S.; Jacques, P.; Taylor, A.; Chiu, C.-J.; Wiggs, J.L.; Seddon, J.M.; Hankinson, S.E.; et al. Contribution of the Nurses’ Health Study to the Epidemiology of Cataract, Age-Related Macular Degeneration, and Glaucoma. Am. J. Public Health 2016, 106, 1684–1689. [Google Scholar] [CrossRef] [PubMed]
- Seddon, J.M. Genetic and environmental underpinnings to age-related ocular diseases. Investig. Ophthalmol. Vis. Sci. 2013, 54, ORSF28–ORSF30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turati, F.; Filomeno, M.; Galeone, C.; Serraino, D.; Bidoli, E.; La Vecchia, C. Dietary glycemic index, glycemic load and risk of age-related cataract extraction: A case-control study in Italy. Eur. J. Nutr. 2015, 54, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, D.Y.; Shen, J.; Wu, Y.B.; Sun, Q.Z.; Dong, J.M.; Yu, J.-C. Prevalence and risk factors on age-related cataract and surgery in adults over 50 years old in Binhu District, Wuxi, China. Int. J. Ophthalmol. 2020, 13, 445–451. [Google Scholar] [CrossRef]
- Shiels, A.; Hejtmancik, J.F. Molecular Genetics of Cataract. Prog. Mol. Biol. Transl. Sci. 2015, 134, 203–218. [Google Scholar]
- Ma, X.; Jiao, X.; Ma, Z.; Hejtmancik, J.F. Polymorphism rs7278468 is associated with Age-related cataract through decreasing transcriptional activity of the CRYAA promoter. Sci Rep. 2016, 6, 23206. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.B.; Qiao, C.; Wei, L.; Han, Y.D.; Cui, N.H.; Huang, Z.L.; Li, Z.-H.; Zheng, F.; Yan, M. Associations of Polymorphisms in MTHFR Gene with the Risk of Age-Related Cataract in Chinese Han Population: A Genotype-Phenotype Analysis. PLoS ONE 2015, 10, e145581. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Daily, J.W.; Song, M.Y.; Kwon, H.K. Gene-gene and gene-lifestyle interactions of AKAP11, KCNMA1, PUM1, SPTBN1, and EPDR1 on osteoporosis risk in middle-aged adults. Nutrition 2020, 79–80, 110859. [Google Scholar] [CrossRef]
- Park, S.; Ahn, J.; Lee, B.K. Self-rated Subjective Health Status Is Strongly Associated with Sociodemographic Factors, Lifestyle, Nutrient Intakes, and Biochemical Indices, but Not Smoking Status: KNHANES 2007–2012. J. Korean Med. Sci. 2015, 30, 1279–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Han, B.G. Cohort Profile: The Korean Genome and Epidemiology Study (KoGES) Consortium. Int. J. Epidemiol. 2017, 46, 1350. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.; Kwon, E.; Shim, J.E.; Park, M.K.; Joo, Y.; Kimm, K.; Park, C.; Kim, D.H. Validation and reproducibility of food frequency questionnaire for Korean genome epidemiologic study. Eur. J. Clin. Nutr. 2007, 61, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kang, S. A minor allele of the haplotype located in the 19q13 loci is associated with a decreased risk of hyper-LDL-cholesterolemia, and a balanced diet and high protein intake can reduce the risk. Lipids Health Dis. 2020, 19, 178. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, M.K. Relationship of sodium intake with obesity among Korean children and adolescents: Korea National Health and Nutrition Examination Survey. Br. J. Nutr. 2016, 115, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Rabbee, N.; Speed, T.P. A genotype calling algorithm for Affymetrix SNP arrays. Bioinformatics 2006, 22, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Zeng, X.; Fan, Y.; Su, Y.; Ma, Q.; Zhu, J.; Yao, H. Gene Polymorphism Association with Type 2 Diabetes and Related Gene-Gene and Gene-Environment Interactions in an Uyghur Population. Med. Sci Monit. 2016, 22, 474–487. [Google Scholar]
- Xu, H.M.; Sun, X.W.; Qi, T.; Lin, W.Y.; Liu, N.; Lou, X.Y. Multivariate dimensionality reduction approaches to identify gene-gene and gene-environment interactions underlying multiple complex traits. PLoS ONE 2014, 9, e108103. [Google Scholar] [CrossRef]
- Uma Jyothi, K.; Reddy, B.M. Gene-gene and gene-environment interactions in the etiology of type 2 diabetes mellitus in the population of Hyderabad, India. Meta Gene 2015, 5, 9–20. [Google Scholar] [CrossRef]
- Tu, Y.; Li, L.; Qin, B.; Wu, J.; Cheng, T.; Kang, L.; Guan, H. Long noncoding RNA glutathione peroxidase 3-antisense inhibits lens epithelial cell apoptosis by upregulating glutathione peroxidase 3 expression in age-related cataract. Mol. Vis. 2019, 25, 734–744. [Google Scholar]
- Berry, V.; Francis, P.; Reddy, M.A.; Collyer, D.; Vithana, E.; MacKay, I.; Dawson, G.; Carey, A.H.; Moore, A.; Bhattacharya, S.S.; et al. Alpha-B crystallin gene (CRYAB) mutation causes dominant congenital posterior polar cataract in humans. Am. J. Hum. Genet. 2001, 69, 1141–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, I.M.; Tran, M.; Musarella, M.A. Ocular genetics: Current understanding. Surv. Ophthalmol. 2004, 49, 159–196. [Google Scholar] [CrossRef] [PubMed]
- Asbell, P.A.; Dualan, I.; Mindel, J.; Brocks, D.; Ahmad, M.; Epstein, S. Age-related cataract. Lancet 2005, 365, 599–609. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, X.; Wang, J.; Jin, L.; Huang, W.; Luo, Y.; Lu, Y. Risk factors of age-related cataract in a Chinese adult population: The Taizhou Eye Study. Clin. Exp. Ophthalmol. 2018, 46, 371–379. [Google Scholar] [CrossRef]
- Rim, T.H.; Kim, M.H.; Kim, W.C.; Kim, T.I.; Kim, E.K. Cataract subtype risk factors identified from the Korea National Health and Nutrition Examination survey 2008–2010. BMC Ophthalmol. 2014, 14, 4. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Kim, S.A.; Kim, N.R.; Rhee, J.-A.; Yun, Y.-W.; Shin, M.-H. Risk Factors for Falls in Older Korean Adults: The 2011 Community Health Survey. J. Korean Med. Sci. 2014, 29, 1482–1487. [Google Scholar] [CrossRef]
- Yu, X.; Lyu, D.; Dong, X.; He, J.; Yao, K. Hypertension and risk of cataract: A meta-analysis. PLoS ONE 2014, 9, e114012. [Google Scholar] [CrossRef]
- Kanthan, G.L.; Mitchell, P.; Burlutsky, G.; Wang, J.J. Fasting blood glucose levels and the long-term incidence and progression of cataract—The Blue Mountains Eye Study. Acta Ophthalmol. 2011, 89, e434–e438. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, E.M.; Marklund, S.L.; Behndig, A. Glucose-induced cataract in CuZn-SOD null lenses: An effect of nitric oxide? Free Radic. Biol. Med. 2007, 42, 1098–1105. [Google Scholar] [CrossRef]
- Balog, Z.; Sikic, J.; Vojnikovic, B.; Balog, S. Senile cataract and the absorption activity of cytochrome C oxidase. Coll. Antropol. 2001, 25, 33–36. [Google Scholar]
- Bae, J.H.; Shin, D.S.; Lee, S.C.; Hwang, I.C. Sodium Intake and Socioeconomic Status as Risk Factors for Development of Age-Related Cataracts: The Korea National Health and Nutrition Examination Survey. PLoS ONE 2015, 10, e136218. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, Y.; Ishimori, N.; Oguchi, J.; Nagai, N.; Kimura, M.; Funakoshi-Tago, M.; Tamura, H. Coffee brew intake can prevent the reduction of lens glutathione and ascorbic acid levels in HFD-fed animals. Exp. Ther Med. 2019, 17, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Varma, S.D. Effect of coffee (caffeine) against human cataract blindness. Clin. Ophthalmol. 2016, 10, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Lou, L.X.; He, J.J.; Xu, Y.F. Body mass index and risk of age-related cataract: A meta-analysis of prospective cohort studies. PLoS ONE 2014, 9, e89923. [Google Scholar] [CrossRef]
- Tan, A.G.; Kifley, A.; Flood, V.M.; Holliday, E.G.; Scott, R.J.; Cumming, R.G.; Mitchell, P.; Wang, J.J. Evaluating the associations between obesity and age-related cataract: A Mendelian randomization study. Am. J. Clin. Nutr. 2019, 110, 969–976. [Google Scholar] [CrossRef]
- Kim, D.J. The epidemiology of diabetes in Korea. Diabetes Metab. J. 2011, 35, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Choi, H.N.; Hwang, J.Y.; Chang, N.; Kim, W.Y.; Chung, H.W.; Yang, Y.J. Development and evaluation of a food frequency questionnaire for Vietnamese female immigrants in Korea: The Korean Genome and Epidemiology Study (KoGES). Nutr. Res. Pract. 2011, 5, 260–265. [Google Scholar] [CrossRef]
- Kim, D.; Kim, J. Dairy consumption is associated with a lower incidence of the metabolic syndrome in middle-aged and older Korean adults: The Korean Genome and Epidemiology Study (KoGES). Br. J. Nutr. 2017, 117, 148–160. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Parameters | Men | Women | ||
|---|---|---|---|---|
| Non-ARC (n = 14,806) | ARC (n = 805) | Non-ARC (n = 24,289) | ARC (n = 1167) | |
| Age (years) | 59.5 ± 5.7 c | 63.0 ± 5.6 a | 56.8 ± 5.3 d | 61.0 ± 5.2 b***+++ |
| BMI (kg/m2) | 24.4 ± 2.6 a | 24.5 ± 2.7 a | 23.9 ± 2.9 b | 23.9 ± 3.0 b+++ |
| Waist circumferences (cm) | 85.2 ± 7.3 a | 85.2 ± 7.3 a | 79.7 ± 7.9 b | 80.0 ± 8.0 b+++ |
| Serum glucose (mg/dL) | 99.6 ± 23 b | 103 ± 30 a | 94.8 ± 18.3 c | 98.6 ± 23 b***+++ |
| HbA1c (%) | 5.81 ± 0.83 b | 5.90 ± 0.96 a | 5.78 ± 0.68 b | 5.95 ± 0.94 a*** |
| Serum total cholesterol (mg/dL) | 191 ± 35 b | 185 ± 37 c | 207 ± 36 a | 206 ± 38 a**+++ |
| Serum LDL (mg/dL) | 112 ± 33 b | 108 ± 33 b | 125 ± 33 a | 125 ± 35 a+++ |
| Serum HDL (mg/dL) | 50.0 ± 12.1 b | 49.9 ± 12.2 b | 56.9 ± 13.1 a | 56.3 ± 12.5 a+++ |
| Serum triglyceride (mg/dL) | 142 ± 95 a | 134 ± 109 a | 122 ± 74 b | 127 ± 74 b+++ |
| Hypertension (%) | 4935 (35.3) | 351 (43.6) +++ | 6504 (26.8) | 452 (38.7) *+++ |
| Metabolic syndrome (%) | 2711 (19.4) | 196 (24.4) | 3762 (15.5) | 302 (25.9) ***+ |
| Education (Number, %) | ||||
| <High school High school College or more | 1440 (15.8) 2104 (23.0) 5589 (61.2) | 103 (20.1) 121 (23.6) 288 (56.3) | 5395 (26.4) 5536 (27.1) 9487 (46.5) | 377 (38.7) +++ 274 (28.1) 324 (33.2) |
| Income (Number, %) <USD 1000/month | 1405 (10.6) | 111 (14.6) | 3457 (15.2) | 311 (28.7) +++ |
| USD 1000–2000 | 2962 (22.4) | 197 (25.8) | 5829 (25.6) | 333 (30.7) |
| USD 2000–4000 | 5577 (42.1) | 300 (39.3) | 9272 (40.8) | 341 (31.5) |
| USD 4000 | 3308 (25.0) | 155 (20.3) | 4172 (18.4) | 99 (9.1) |
| Exercise (Number, %) No Yes | 5472 (39.2) 8487 (60.8) | 272 (34.1) 526 (65.9) | 11,121 (45.9) 13,102 (54.1) | 536 (46.1) +++ 627 (53.9) |
| Smoking (Number, %) No Former smoking | 4689 (33.5) 6005 (42.9) | 264 (32.9) 348 (43.3) | 23719 (97.7) 228 (0.94) | 1144 (98.0) ***++ 14 (0.26) |
| Smoking | 2250 (23.7) | 192 (23.9) | 332 (1.37) | 9 (0.78) |
| Alcohol intake (Number, %) | ||||
| No (0 g/day) Mild (0–20 g/day) | 4568 (32.6) 180 (1.29) | 302 (37.5) + 9 (1.12) | 18,555 (76.4) 643 (2.65) | 988 (84.7) +++ 26 (2.23) |
| Moderate (≥20 g/day) | 9253 (66.1) | 494 (61.4) | 5091 (21.0) | 153 (13.1) |
| Coffee intake (Number %) | ||||
| Low (<3 g/day) | 4660 (33.3) | 308 (38.3) ++ | 10,883 (44.8) | 645 (55.3) *+++ |
| Medium (3–16 g/day) | 9161 (65.4) | 493 (61.2) | 13,250 (54.6) | 514 (44.0) |
| High (≥16 g/day) | 180 (1.29) | 4 (0.50) | 156 (0.64) | 8 (0.69) |
| Balanced diet pattern (Number, %) | ||||
| Low (<70th percentile) | 9883 (70.6) | 582 (72.3) | 15,810 (65.1) | 823 (70.5) *** |
| High (≥70th percentile) | 4118 (29.4) | 233 (27.7) | 8479 (34.9) | 344 (29.5) |
| Western-style diet pattern (Number, %) | ||||
| Low (<70th percentile) | 10,012 (71.5) | 623 (77.4) *** | 19,869 (81.8) | 989 (84.8) * |
| High (≥70th percentile) | 3989 (28.5) | 182 (22.6) | 4420 (18.2) | 178 (15.3) |
| Rice-based diet pattern (Number, %) | ||||
| Low (<70th percentile) | 10,009 (71.5) | 592 (73.5) | 19,456 (80.1) | 993 (85.1) *** |
| High (≥70th percentile) | 3992 (28.5) | 213 (26.5) | 4833 (19.9) | 174 (14.9) |
| Chr a | SNP b | Position | Mi c | Ma d | OR e | p-Value Adjusted f | MAF g | p-Value for HWE h | Gene | Functional Location |
|---|---|---|---|---|---|---|---|---|---|---|
| 3 | rs1417380362 | 41898108 | C | T | 0.770 | 1.13 × 10−5 | 0.1177 | 0.5644 | ULK4 | intron |
| 5 | rs117418426 | 150398496 | G | A | 1.648 | 5.74 × 10−5 | 0.01404 | 0.8807 | GPX3 | intron |
| 7 | rs200053781 | 8250586 | T | G | 0.859 | 5.55 × 10−5 | 0.3477 | 0.9565 | ICA1 | intron |
| 7 | rs147082589 | 97954290 | C | T | 1.684 | 1.26 × 10−5 | 0.01497 | 1.0 | BAIAP2L1 | intron |
| 7 | rs322348 | 136992106 | C | A | 0.700 | 4.77 × 10−5 | 0.05569 | 0.9063 | PTN | intron |
| 9 | rs553983141 | 131368777 | G | T | 1.493 | 9.28 × 10−5 | 0.02212 | 0.6329 | SPTAN1 | intron |
| 10 | rs117583209 | 105320759 | G | A | 1.658 | 3.76 × 10−5 | 0.01418 | 0.3722 | NEURL1 | intron |
| 11 | rs2070894 | 111780837 | G | A | 0.837 | 8.61 × 10−5 | 0.2054 | 0.3761 | CRYAB | intron |
| 17 | rs55785344 | 31914770 | T | C | 1.211 | 9.44 × 10−5 | 0.1311 | 0.9711 | ACCN1 | upstream transcript |
| 17 | rs879419608 | 71159820 | C | T | 0.804 | 5.78 × 10−5 | 0.1367 | 0.1619 | SSTR2 | upstream transcript |
| Genetic Model | Adjusted for Sex, Age | Adjusted for Sex, Age, Residence Area, BMI, Survey Year | ||||||
|---|---|---|---|---|---|---|---|---|
| TRBA | TEBA | p-Value | CVC | TRBA | TEBA | p-Value | CVC | |
| CRYAB_rs2070894 | 0.5236 | 0.5142 | 8 (0.055) | 7/10 | 0.5237 | 0.5143 | 8 (0.055) | 7/10 |
| ULK4_rs1417380362 SSTR2_rs879419608 | 0.5345 | 0.5206 | 9 (0.011) | 7/10 | 0.5345 | 0.5206 | 9 (0.011) | 7/10 |
| ULK4_rs1417380362 ACCN1_rs55785344 SSTR2_rs879419608 | 0.5393 | 0.5175 | 8 (0.055) | 3/10 | 0.5393 | 0.5175 | 8 (0.055) | 3/10 |
| ULK4_rs1417380362 CRYAB_rs2070894 ACCN1_rs55785344 SSTR2_rs879419608 | 0.5479 | 0.5228 | 8 (0.055) | 6/10 | 0.5479 | 0.5228 | 8 (0.0547) | 6/10 |
| PTN_rs322348 plus model 4 | 0.5581 | 0.5247 | 10 (0.001) | 10/10 | 0.5581 | 0.5248 | 10 (0.001) | 10/10 |
| ICA1_rs200053781 plus model 5 | 0.5673 | 0.5292 | 10 (0.001) | 10/10 | 0.5673 | 0.5283 | 10 (0.001) | 10/10 |
| BAIAP2L1_rs147082 plus model 6 | 0.5743 | 0.5257 | 10 (0.001) | 7/10 | 0.5743 | 0.5263 | 10 (0.001) | 7/10 |
| SPTAN1_rs553983141 plus model 7 | 0.5808 | 0.5215 | 9 (0.011) | 5/10 | 0.5807 | 0.5243 | 9 (0.011) | 5/10 |
| NEURL1_rs11758320 plus model 8 | 0.5873 | 0.5274 | 10 (0.001) | 10/10 | 0.5873 | 0.5272 | 10 (0.001) | 10/10 |
| GPX3_rs117418426 plus model 9 | 0.5922 | 0.5277 | 9 (0.011) | 10/10 | 0.5922 | 0.5281 | 9 (0.011) | 10/10 |
| Groups | Low-PRS (n = 2295) | Medium-PRS (n = 29,067) | High-PRS (n = 8949) | Genetic Variant–MetS Interaction p-Value |
|---|---|---|---|---|
| Middle-aged Elderly a | 1 | 2.07 (1.27–3.38) 1.43 (0.87–2.35) | 2.92 (1.76–4.84) 2.03 (1.21–3.42) | < 0.0001 |
| Men Women | 1 | 1.98 (1.09–3.59) 1.63 (1.06–2.50) | 2.48 (1.51–5.11) 2.35 (1.50–3.67) | 0.813 |
| Without MetS With MetS | 1 | 1.86 (1.22–2.84) 1.47 (0.80–2.72) | 2.81 (1.82–4.34) 1.74 (0.91–3.32) | 0.715 |
| Normal waist High waist b | 1 | 1.75 (1.23–2.47) 1.59 (1.07–2.38) | 2.48 (1.73–3.55) 2.53 (1.67–3.83) | 0.279 |
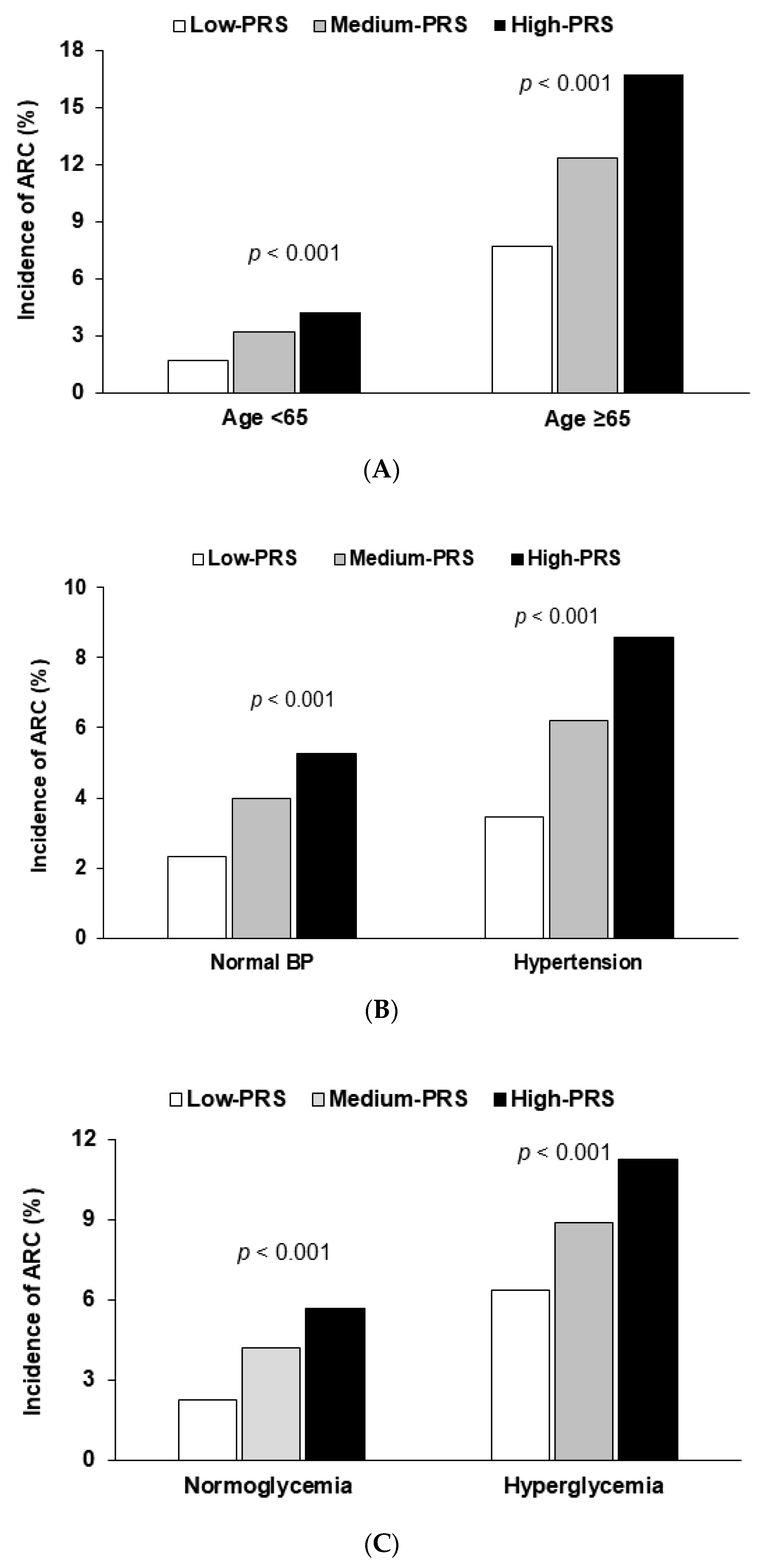
| Normotension Hypertension c | 1 | 1.73 (1.10–2.71) 1.76 (1.03–3.04) | 2.38 (1.49–3.80) 2.61 (1.03–3.04) | 0.030 |
| Low serum glucose High serum glucose d | 1 | 1.91 (1.28–2.87) 1.35 (0.70–2.58) | 2.78 (1.83–4.21) 1.86 (0.92–3.79) | 0.042 |
| Groups | Low-PRS (n = 2295) | Medium-PRS (n = 29,067) | High-PRS (n = 8949) | Genetic Variant–MetS Interaction p-Value |
|---|---|---|---|---|
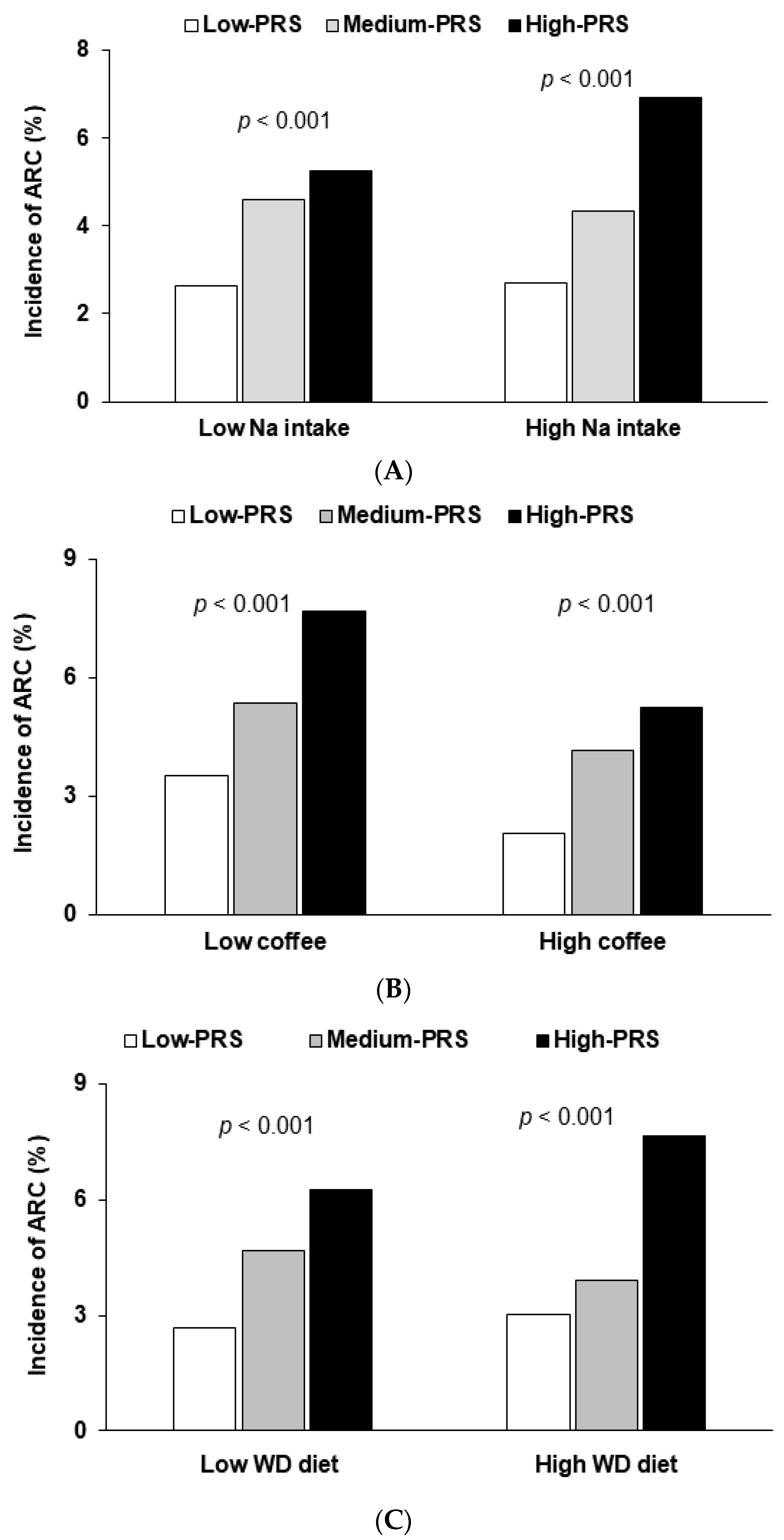
| Low Na intake High Na intake a | 1 | 1.85 (1.54–3.70) 1.52 (0.83–5.01) | 1.58 (1.14–3.50) 2.68 (1.43–5.01) | 0.016 |
| Low coffee intake High coffee intake b | 1 | 1.77 (1.05–3.00) 1.71 (1.08–2.72) | 2.93 (1.71–5.04) 2.15 (1.33–3.45) | 0.049 |
| Low BD intake High BD intake c | 1 | 1.75 (1.24~2.48) 1.67 (1.13–2.47) | 2.48 (1.73~3.55) 2.16 (1.44–3.24) | 0.648 |
| Low WD intake High WD intake d | 1 | 1.75 (1.23–2.47) 1.20(0.27–5.27) | 2.27 (0.50–10.3) 2.48 (1.73–3.55) | 0.049 |
| Low RD intake High RD intake e | 1 | 1.75 (1.24–2.48) 2.04 (1.34–3.12) | 2.48 (1.73–3.55) 2.94 (1.90–4.54) | 0.146 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jee, D.; Kang, S.; Huang, S.; Park, S. Polygenetic-Risk Scores Related to Crystallin Metabolism Are Associated with Age-Related Cataract Formation and Interact with Hyperglycemia, Hypertension, Western-Style Diet, and Na Intake. Nutrients 2020, 12, 3534. https://doi.org/10.3390/nu12113534
Jee D, Kang S, Huang S, Park S. Polygenetic-Risk Scores Related to Crystallin Metabolism Are Associated with Age-Related Cataract Formation and Interact with Hyperglycemia, Hypertension, Western-Style Diet, and Na Intake. Nutrients. 2020; 12(11):3534. https://doi.org/10.3390/nu12113534
Chicago/Turabian StyleJee, Donghyun, Suna Kang, ShaoKai Huang, and Sunmin Park. 2020. "Polygenetic-Risk Scores Related to Crystallin Metabolism Are Associated with Age-Related Cataract Formation and Interact with Hyperglycemia, Hypertension, Western-Style Diet, and Na Intake" Nutrients 12, no. 11: 3534. https://doi.org/10.3390/nu12113534
APA StyleJee, D., Kang, S., Huang, S., & Park, S. (2020). Polygenetic-Risk Scores Related to Crystallin Metabolism Are Associated with Age-Related Cataract Formation and Interact with Hyperglycemia, Hypertension, Western-Style Diet, and Na Intake. Nutrients, 12(11), 3534. https://doi.org/10.3390/nu12113534