
Commercial Vaccines Do Not Confer Protection against Two Genogroups of Piscirickettsia salmonis, LF-89 and EM-90, in Atlantic Salmon

 , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Commercial Vaccines
2.3. Challenge with LF-89 Genogroup (Trial 1)
2.4. Challenge with EM-90-LIKE Genogroup and Coinfection with sea Lice (Trial 2)
2.5. Necropsy Analysis
2.6. ELISA
2.7. Statistical Analysis
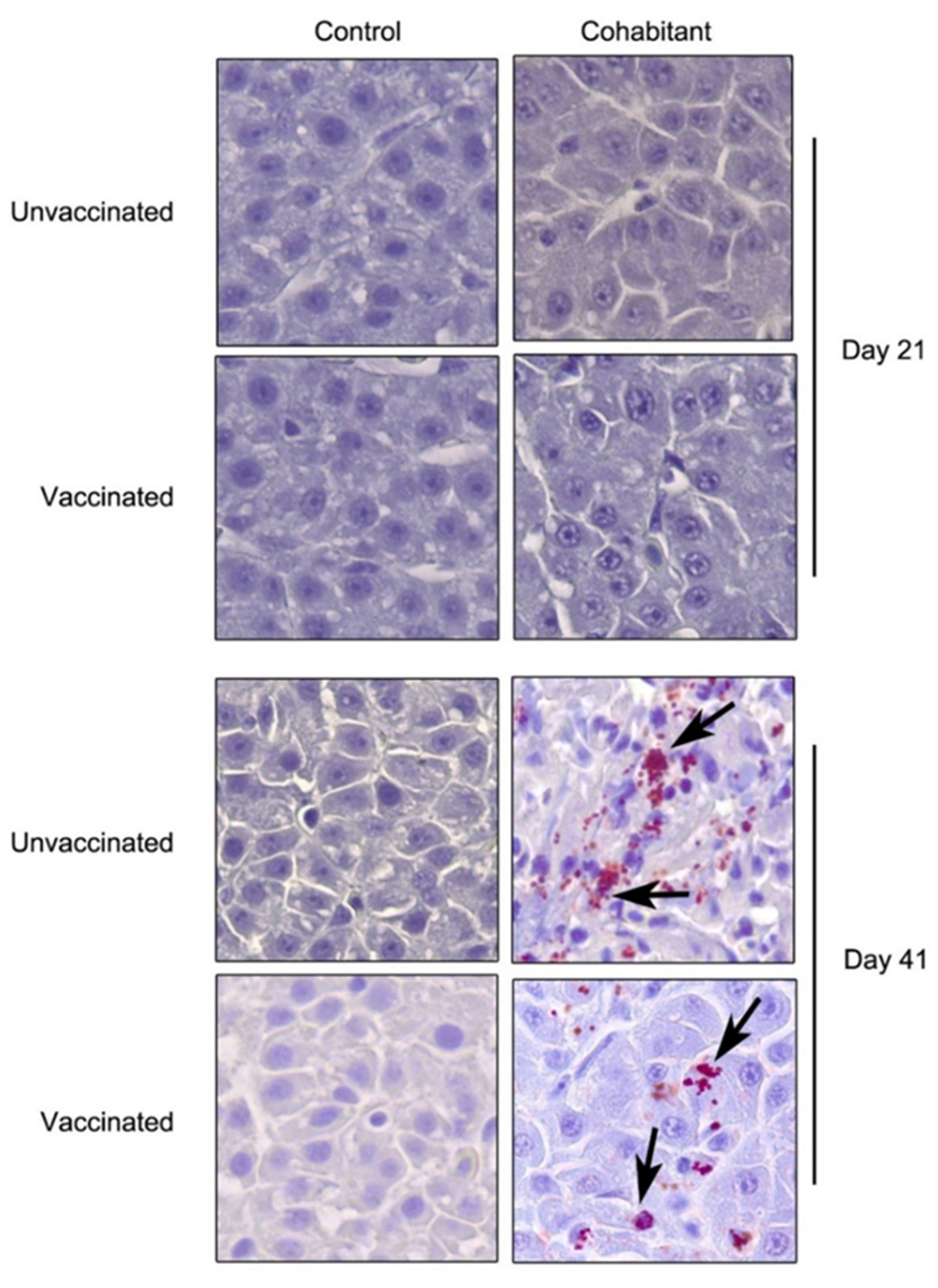
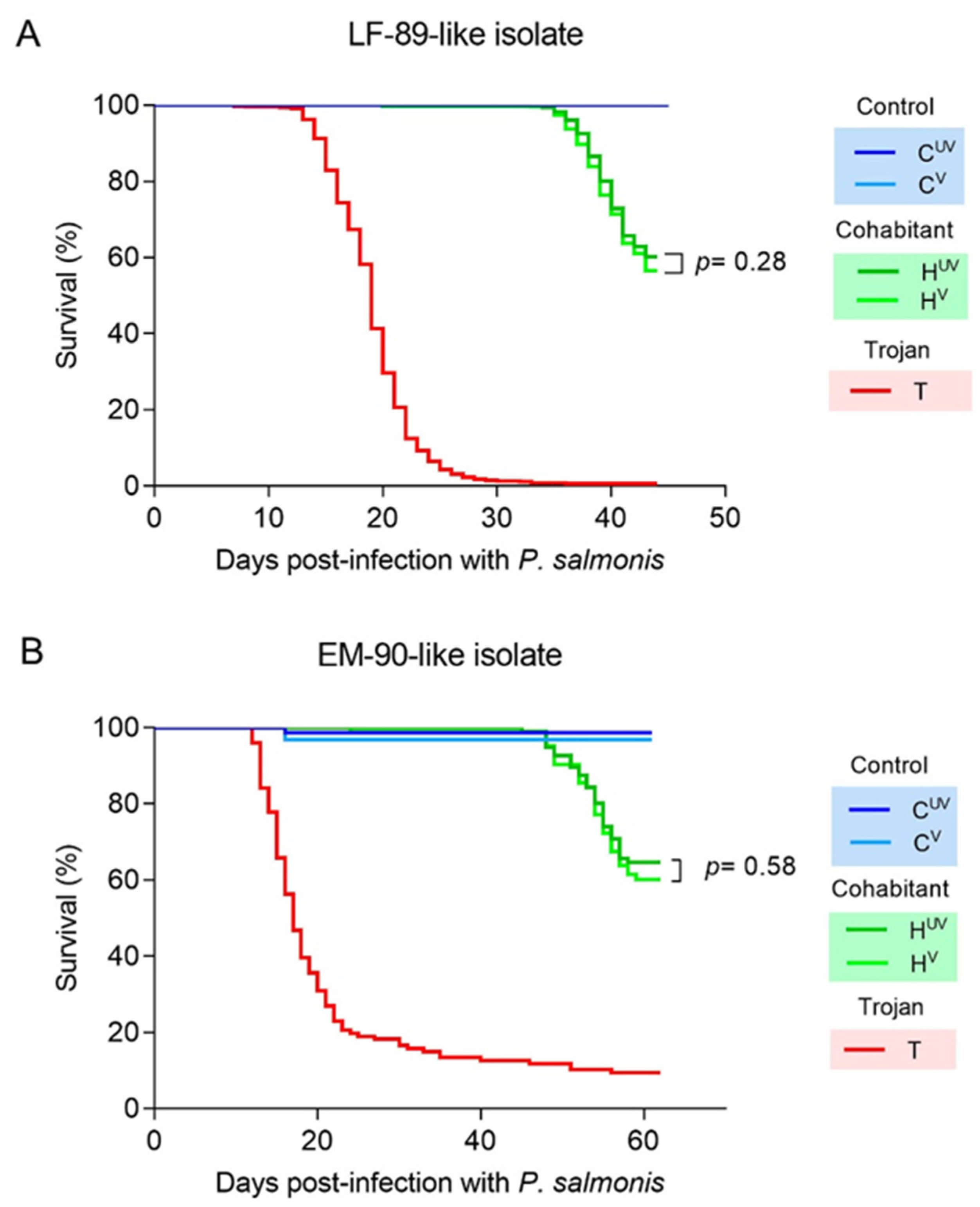
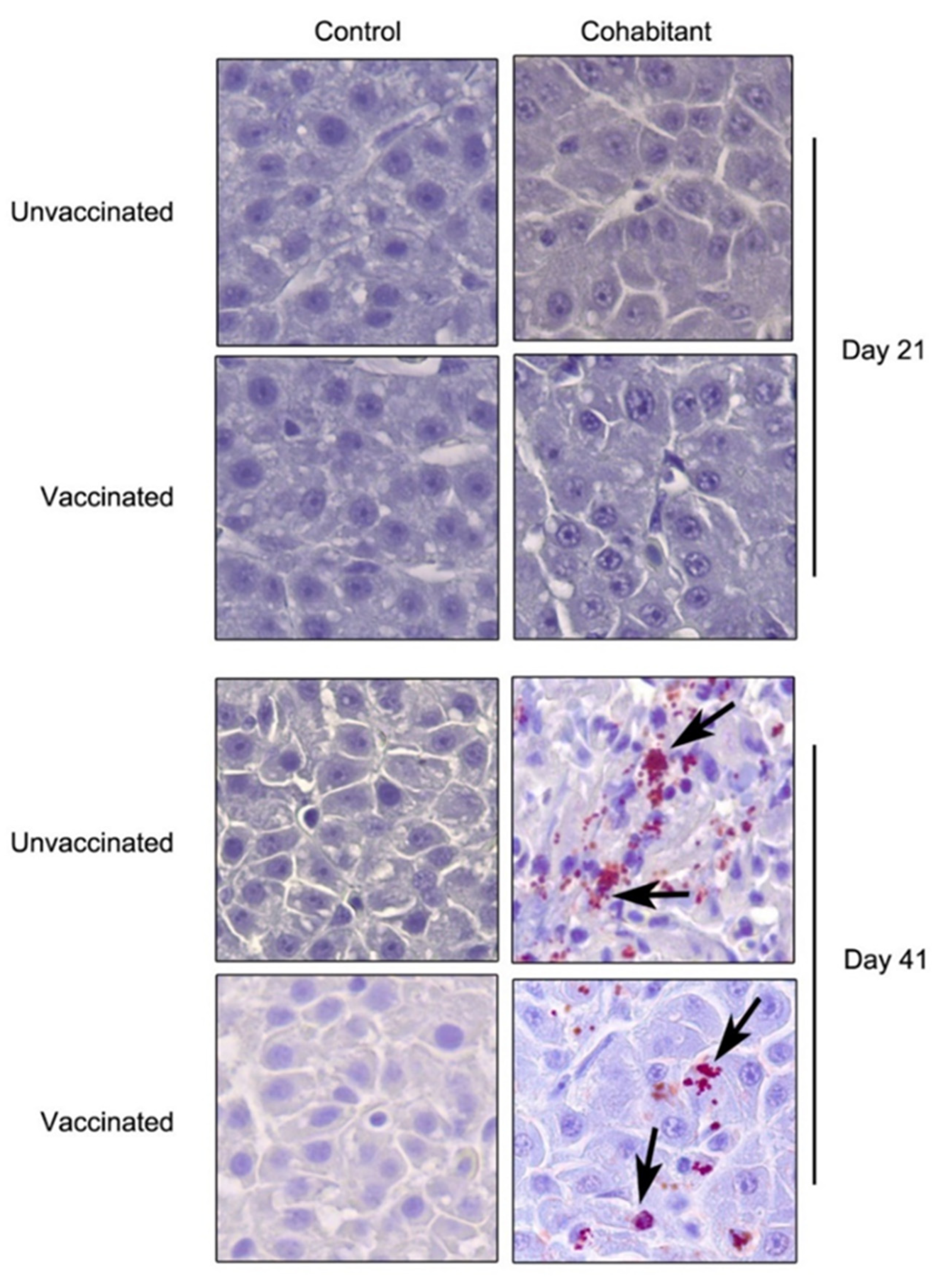
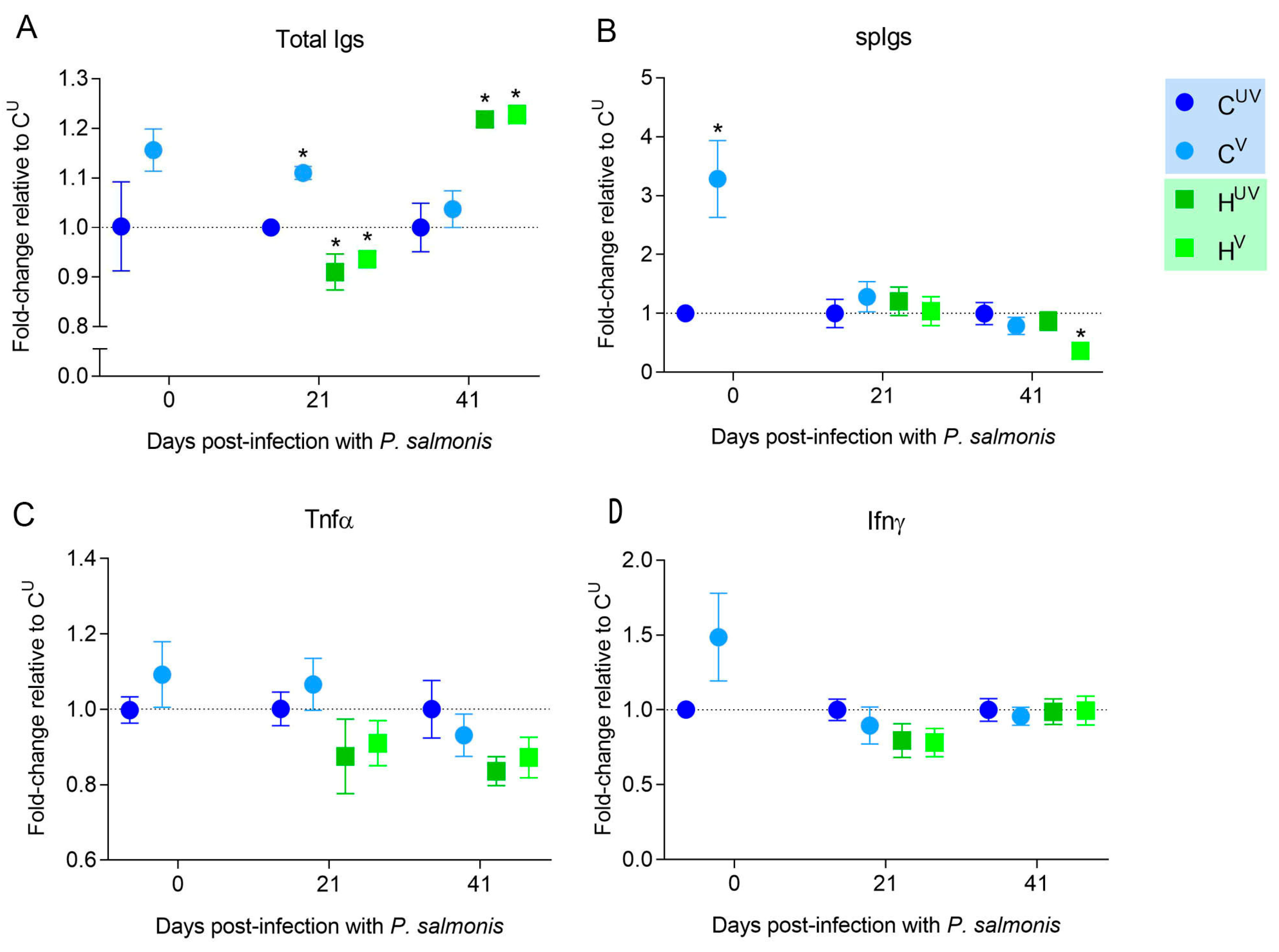
3. Results
3.1. Vaccine Efficacy against the LF-89 Genogroup
3.2. Vaccine Efficacy against the EM-90 Genogroup with Low Sea Lice Coinfection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maisey, K.; Montero, R.; Christodoulides, M. Vaccines for piscirickettsiosis (salmonid rickettsial septicaemia, SRS): The Chile perspective. Expert Rev. Vaccines 2017, 16, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Rozas, M.; Enriquez, R. Piscirickettsiosis and Piscirickettsia salmonis in fish: A review. J. Fish Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef] [PubMed]
- SERNAPESCA. Informe Sanitario Salmonicultura en Centros Marinos 2019; Servicio Nacional de Pesca y Acuicultura: Santiago, Chile, 2020; pp. 1–38. [Google Scholar]
- Brocklebank, J.; Ebelyn, T.; Speare, D.J.; Armstrong, R. Rickettsial septicemia in farmed Atlantic and chinook salmon in British Columbia: Clinical presentation and experimental transmission. Can. Vet. J. 1993, 34, 745–748. [Google Scholar]
- Grant, A.N.; Brown, A.G.; Cox, D.I.; Birkbeck, T.H.; Griffen, A.A. Rickettsia-like organism in farmed salmon. Vet. Rec. 1996, 138, 423. [Google Scholar]
- Olsen, A.J.; Melby, H.; Speilberg, L.; Evensen, O.; Hastein, T. Piscirickettsia salmonis infection in Atlantic salmon Salmo salar in Norway-epidemiological, pathological and microbiological findings. Dis. Aquat. Org. 1997, 31, 35–48. [Google Scholar] [CrossRef] [Green Version]
- SERNAPESCA. Informe Sanitario de Salmonicultura en Centros Marinos 2016; Servicio Nacional de Pesca y Acuicultura: Santiago, Chile, 2017; pp. 1–37. [Google Scholar]
- Happold, J.; Sadler, R.; Meyer, A.; Hillman, A.; Cowled, B.; Mackenzie, C.; Gallardo Lagno, A.L.; Cameron, A. Effectiveness of vaccination for the control of salmonid rickettsial septicaemia in commercial salmon and trout farms in Chile. Aquaculture 2020, 520, 734968. [Google Scholar] [CrossRef]
- Figueroa, C.; Bustos, P.; Torrealba, D.; Dixon, B.; Soto, C.; Conejeros, P.; Gallardo, J.A. Coinfection takes its toll: Sea lice override the protective effects of vaccination against a bacterial pathogen in Atlantic salmon. Sci. Rep. 2017, 7, 17817. [Google Scholar] [CrossRef]
- Figueroa, C.; Veloso, P.; Espin, L.; Dixon, B.; Torrealba, D.; Elalfy, I.S.; Afonso, J.M.; Soto, C.; Conejeros, P.; Gallardo, J.A. Host genetic variation explains reduced protection of commercial vaccines against Piscirickettsia salmonis in Atlantic salmon. Sc. Rep. 2020, 10, 18252. [Google Scholar] [CrossRef]
- Nourdin-Galindo, G.; Sanchez, P.; Molina, C.F.; Espinoza-Rojas, D.A.; Oliver, C.; Ruiz, P.; Vargas-Chacoff, L.; Carcamo, J.G.; Figueroa, J.E.; Mancilla, M.; et al. Comparative Pan-Genome Analysis of Piscirickettsia salmonis Reveals Genomic Divergences within Genogroups. Front. Cell. Infect. Microbiol. 2017, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Otterlei, A.; Brevik, O.J.; Jensen, D.; Duesund, H.; Sommerset, I.; Frost, P.; Mendoza, J.; McKenzie, P.; Nylund, A.; Apablaza, P. Phenotypic and genetic characterization of Piscirickettsia salmonis from Chilean and Canadian salmonids. BMC Vet. Res. 2016, 12, 55. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, J.; Hernandez, N.; Osses, A.; Castillo, A.; Cancino, A.; Grothusen, H.; Navas, E.; Henriquez, P.; Bohle, H.; Bustamante, F.; et al. Prevalence, geographic distribution and phenotypic differences of Piscirickettsia salmonis EM-90-like isolates. J. Fish Dis. 2017, 40, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, A.; Tapia, P.; Gomez, F.; Marshall, S.; Fuentes, D.; BValdes, J. Draft genomes and reference transcriptomes extend the coding potential of the fish pathogen Piscirickettsia salmonis. Electron. J. Biotechnol. 2018, 33, 36–38. [Google Scholar] [CrossRef]
- Bohle, H.; Hemríquez, P.; Grothusen, H.; Navas, E.; Sandoval, A.; Bustamante, F.; Bstos, P.; Mancilla, M. Comparative Genome Analysis of Two Isolates of the Fish Pathogen Piscirickettsia salmonis from Different Hosts Reveals Major Differences in Virulence-Associated Secretion Systems. Genomea 2014, 2, e01219-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas-Serri, M.; Ildefonso, R.; Pena, A.; Enriquez, R.; Barrientos, S.; Maldonado, L. Comparative pathogenesis of piscirickettsiosis in Atlantic salmon (Salmo salar L.) post-smolt experimentally challenged with LF-89-like and EM-90-like Piscirickettsia salmonis isolates. J. Fish Dis. 2017, 40, 1451–1472. [Google Scholar] [CrossRef]
- Rozas-Serri, M.; Pena, A.; Arriagada, G.; Enriquez, R.; Maldonado, L. Comparison of gene expression in post-smolt Atlantic salmon challenged by LF-89-like and EM-90-like Piscirickettsia salmonis isolates reveals differences in the immune response associated with pathogenicity. J. Fish Dis. 2018, 41, 539–552. [Google Scholar] [CrossRef]
- Cardella, M.A.; Eimers, M.E. Safety and Potency Testing of Federally Licensed Fish Bacterins. J. Aquat. Anim. Health 1990, 2, 49–55. [Google Scholar] [CrossRef]
- Meza, K.; Inami, M.; Dalum, A.S.; Lund, H.; Bjelland, A.M.; Sorum, H.; Lovoll, M. Comparative evaluation of experimental challenge by intraperitoneal injection and cohabitation of Atlantic salmon (Salmo salar L) after vaccination against Piscirickettsia salmonis (EM90-like). J. Fish Dis. 2019, 42, 1713–1730. [Google Scholar] [CrossRef] [Green Version]
- Araya, A.; Mancilla, M.; Lhorente, J.P.; Neira, R.; Gallardo, J.A. Experimental challenges of Atlantic salmon Salmo salar with incremental levels of copepodids of sea louse Caligus rogercresseyi: Effects on infestation and early development. Aquacult. Res. 2012, 43, 1904–1908. [Google Scholar] [CrossRef]
- Bravo, S.; Treasurer, J.; Sepulveda, M.; Lagos, C. Effectiveness of hydrogen peroxide in the control of Caligus rogercresseyi in Chile and implications for sea louse management. Aquaculture 2010, 303, 22–27. [Google Scholar] [CrossRef]
- Morales-Lange, B.; González-Aravena, M.; Font, A.; Guzmán, F.; Mercado, L. Detection of peroxiredoxin-like protein in Antarctic sea urchin (Sterechinus neumayeri) under heat stress and induced with pathogen-associated molecular pattern from Vibrio anguillarum. Polar Biol. 2018, 41, 2065–2073. [Google Scholar] [CrossRef]
- Fishman, J.B.; Berg, E.A. Preparation of Antibody Using Caprylic Acid. Cold Spring Harb. Protoc. 2018, 2018, 475–476. [Google Scholar] [CrossRef] [PubMed]
- Carril, G.; Gómez, F.; Marshall, S. Expression of flagellin and key regulatory flagellar genes in the non-motile bacterium Piscirickettsia salmonis. Dis. Aquat. Org. 2017, 123, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Veter. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef]
- Adams, A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol. 2019, 90, 210–214. [Google Scholar] [CrossRef]
- Alvarez, C.A.; Gomez, F.A.; Mercado, L.; Ramirez, R.; Marshall, S.H. Piscirickettsia salmonis Imbalances the Innate Immune Response to Succeed in a Productive Infection in a Salmonid Cell Line Model. PloS OnE 2016, 11, e0163943. [Google Scholar] [CrossRef] [Green Version]
- Cabello, F.C.; Godfrey, H.P. Salmon aquaculture, Piscirickettsia salmonis virulence, and One Health: Dealing with harmful synergies between heavy antimicrobial use and piscine and human health. Aquaculture 2019, 507, 451–456. [Google Scholar] [CrossRef]
- Nordmo, R. Strengths and weaknesses of different challenge methods. Dev. Biol. Standard. 1997, 90, 303–309. [Google Scholar]
- Kuzyk, M.; Burian, J.; Machander, D.; Dolhaine, D.; Cameron, S.; Thornton, J.; Kay, W. An efficacious recombinant subunit vaccine against the salmonid rickettsial pathogen Piscirickettsia salmonis. Vaccine 2001, 19, 2337–2344. [Google Scholar] [CrossRef]
- Salonius, K.; Siderakis, C.; Mackinnon, A.; Griffiths, S. Use of Arthrobacter davidanieli as a live vaccine against Renibacterium salmoninarum and Piscireckettsia salmonis in salmonis. Prog. Fish Vaccinol. 2005, 121, 189–197. [Google Scholar]
- Tobar, J.A.; Jerez, S.; Caruffo, M.; Bravo, C.; Contreras, F.; Bucarey, S.A.; Harel, M. Oral vaccination of Atlantic salmon (Salmo salar) against salmonid rickettsial septicaemia. Vaccine 2011, 29, 2336–2340. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, V.; Miquel, A.; Burzio, L.O.; Rosemblatt, M.; Engel, E.; Valenzuela, S.; Parada, G.; Valenzuela, P.D. A vaccine against the salmonid pathogen Piscirickettsia salmonis based on recombinant proteins. Vaccine 2006, 24, 5083–5091. [Google Scholar] [CrossRef] [PubMed]
- Midtlyng, P. Critical assessment of regulatory standards and tests for fi sh vaccines. Dev. Biol. 2005, 121, 219–226. [Google Scholar]
- Rozas-Serri, M.; Pena, A.; Maldonado, L. Gene expression associated with immune response in Atlantic salmon head-kidney vaccinated with inactivated whole-cell bacterin of Piscirickettsia salmonis and pathogenic isolates. Fish Shellfish Immunol. 2019, 93, 789–795. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Group | Treatments | First Trial | Second Trial |
|---|---|---|---|
| Cohabitant | Vaccinated (HV) | 496 | 83 |
| Unvaccinated (HUV) | 335 | 96 | |
| Total cohabitant (H) | 831 | 179 | |
| HUV/H | 42% | 53% | |
| Trojan | Total Trojans (T) | 2903 | 183 |
| T/(H + T) | 77% | 41% | |
| Control | Vaccinated (CV) | 506 | 38 |
| Unvaccinated (CUV) | 727 | 42 | |
| Total control (C) | 1233 | 80 | |
| Total fish (H + T + C) | 4987 | 442 |
| Group | Pathological Signs | Presence of Pathological Signs | Treatment | Proportion | Chi-Square Test | |||
|---|---|---|---|---|---|---|---|---|
| UV | V | UV | V | X2 | p-Value | |||
| Cohabitant | Vacuolar | No | 2 | 4 | 0.2 | 0.4 | 0.24 | 0.63 |
| degeneration | Yes | 8 | 6 | 0.8 | 0.6 | |||
| Total | 10 | 10 | ||||||
| Hepatitis | No | 9 | 9 | 0.9 | 0.9 | 0 | 1 | |
| Yes | 1 | 1 | 0.1 | 0.1 | ||||
| Total | 10 | 10 | ||||||
| Hepatocyte | No | 8 | 4 | 0.8 | 0.4 | 1.88 | 0.17 | |
| atrophy | Yes | 2 | 6 | 0.2 | 0.6 | |||
| Total | 10 | 10 | ||||||
| Control | Vacuolar | No | 1 | 3 | 0.1 | 0.3 | 0.31 | 0.58 |
| degeneration | Yes | 9 | 7 | 0.9 | 0.7 | |||
| Total | 10 | 10 | ||||||
| Hepatitis | No | 8 | 7 | 0.8 | 0.7 | 0 | 1 | |
| Yes | 2 | 3 | 0.2 | 0.3 | ||||
| Total | 10 | 10 | ||||||
| Hepatocyte | No | 10 | 5 | 1 | 0.5 | 4.27 | <0.05 * | |
| atrophy | Yes | 0 | 5 | 0 | 0.5 | |||
| Total | 10 | 10 | ||||||
| Group | Pathological Signs | Presence of Pathological Signs | Treatment | Proportion | Chi-Square Test | |||
|---|---|---|---|---|---|---|---|---|
| UV | V | UV | V | X2 | p-Value | |||
| Cohabitant | Nodules in | No | 0 | 0 | 0 | 0 | 0 | 1 |
| liver | Yes | 10 | 10 | 1 | 1 | |||
| Total | 10 | 10 | ||||||
| Congestive | No | 1 | 0 | 0.1 | 0 | 0.02 | 0.96 | |
| liver | Yes | 9 | 10 | 0.9 | 1 | |||
| Total | 10 | 10 | ||||||
| Hepatomegaly | No | 0 | 0 | 0 | 0 | 0 | 1 | |
| Yes | 10 | 10 | 1 | 1 | ||||
| Total | 10 | 10 | ||||||
| Control | Nodules in | No | 10 | 10 | 1 | 1 | 0 | 1 |
| liver | Yes | 0 | 0 | 0 | 0 | |||
| Total | 10 | 10 | ||||||
| Congestive | No | 10 | 9 | 1 | 0.9 | 0 | 1 | |
| liver | Yes | 0 | 1 | 0 | 0.1 | |||
| Total | 10 | 10 | ||||||
| Hepatomegaly | No | 10 | 9 | 1 | 0.9 | 0 | 1 | |
| Yes | 0 | 1 | 0 | 0.1 | ||||
| Total | 10 | 10 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa, C.; Torrealba, D.; Morales-Lange, B.; Mercado, L.; Dixon, B.; Conejeros, P.; Silva, G.; Soto, C.; Gallardo, J.A. Commercial Vaccines Do Not Confer Protection against Two Genogroups of Piscirickettsia salmonis, LF-89 and EM-90, in Atlantic Salmon. Biology 2022, 11, 993. https://doi.org/10.3390/biology11070993
Figueroa C, Torrealba D, Morales-Lange B, Mercado L, Dixon B, Conejeros P, Silva G, Soto C, Gallardo JA. Commercial Vaccines Do Not Confer Protection against Two Genogroups of Piscirickettsia salmonis, LF-89 and EM-90, in Atlantic Salmon. Biology. 2022; 11(7):993. https://doi.org/10.3390/biology11070993
Chicago/Turabian StyleFigueroa, Carolina, Débora Torrealba, Byron Morales-Lange, Luis Mercado, Brian Dixon, Pablo Conejeros, Gabriela Silva, Carlos Soto, and José A. Gallardo. 2022. "Commercial Vaccines Do Not Confer Protection against Two Genogroups of Piscirickettsia salmonis, LF-89 and EM-90, in Atlantic Salmon" Biology 11, no. 7: 993. https://doi.org/10.3390/biology11070993
APA StyleFigueroa, C., Torrealba, D., Morales-Lange, B., Mercado, L., Dixon, B., Conejeros, P., Silva, G., Soto, C., & Gallardo, J. A. (2022). Commercial Vaccines Do Not Confer Protection against Two Genogroups of Piscirickettsia salmonis, LF-89 and EM-90, in Atlantic Salmon. Biology, 11(7), 993. https://doi.org/10.3390/biology11070993