
The Modulation of Phospho-Extracellular Signal-Regulated Kinase and Phospho-Protein Kinase B Signaling Pathways plus Activity of Macrophage-Stimulating Protein Contribute to the Protective Effect of Stachydrine on Acetaminophen-Induced Liver Injury

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of ST on HepG2 Cell Viability
2.2. Effects of ST on the Expression of ERK and AKT in HepG2 Cells
2.3. ST Ameliorated APAP-Induced Hepatic Injury
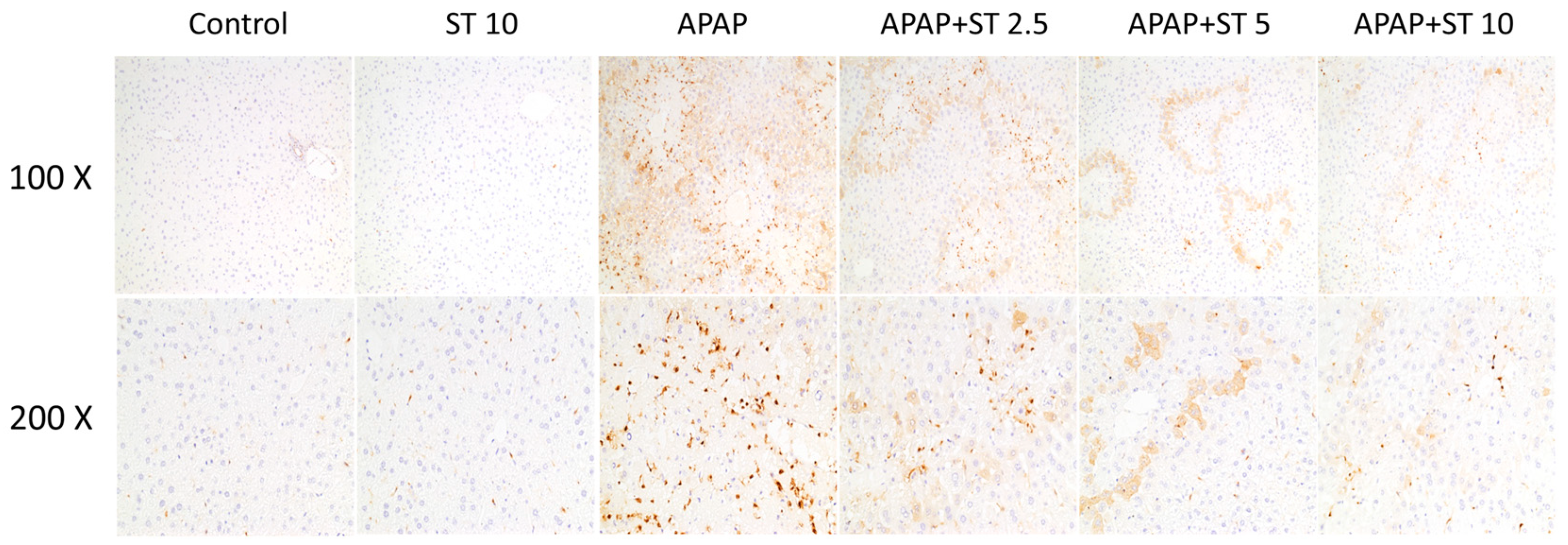
2.4. Effect of ST on Macrophage Accumulation in APAP-Induced Hepatic Injury
2.5. Influence of ST on the Levels of Inflammatory Cytokine in Liver Tissues
2.6. ST Decreased APAP-Induced Oxidative Stress Injury
2.7. Effect of ST on ERK and AKT Expression in Liver Tissues
2.8. Effect of ST on Nrf2 Expression in Liver Tissues
2.9. Effect of ST on MSP Expression in Liver Tissues
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Experimental Design
4.5. Serum Enzyme Determination
4.6. Histology Analysis of Liver Tissues
4.7. Immunohistochemical Analysis on Liver Tissues
4.8. Measurement of Cytokine Levels in Liver Tissues
4.9. Measurement of Liver Oxidative Stress Markers
4.10. Western Blot Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jaeschke, H. Acetaminophen: Dose-Dependent Drug Hepatotoxicity and Acute Liver Failure in Patients. Dig. Dis. 2015, 33, 464–471. [Google Scholar] [CrossRef]
- James, L.P.; Mayeux, P.R.; Hinson, J.A. Acetaminophen-induced hepatotoxicity. Drug Metab. Dispos. 2003, 31, 1499–1506. [Google Scholar] [CrossRef]
- Bunchorntavakul, C.; Reddy, K.R. Acetaminophen-related hepatotoxicity. Clin. Liver Dis. 2013, 17, 587–607. [Google Scholar] [CrossRef]
- Hinson, J.A.; Reid, A.B.; McCullough, S.S.; James, L.P. Acetaminophen-induced hepatotoxicity: Role of metabolic activation, reactive oxygen/nitrogen species, and mitochondrial permeability transition. Drug Metab. Rev. 2004, 36, 805–822. [Google Scholar] [CrossRef] [PubMed]
- Martin-Murphy, B.V.; Holt, M.P.; Ju, C. The role of damage associated molecular pattern molecules in acetaminophen-induced liver injury in mice. Toxicol. Lett. 2010, 192, 387–394. [Google Scholar] [CrossRef]
- Liu, Z.X.; Govindarajan, S.; Kaplowitz, N. Innate immune system plays a critical role in determining the progression and severity of acetaminophen hepatotoxicity. Gastroenterology 2004, 127, 1760–1774. [Google Scholar] [CrossRef] [PubMed]
- Leonard, E.J.; Danilkovitch, A. Macrophage stimulating protein. Adv. Cancer Res. 2000, 77, 139–167. [Google Scholar] [PubMed]
- Li, J.; Chanda, D.; Shiri-Sverdlov, R.; Neumann, D. MSP: An emerging player in metabolic syndrome. Cytokine Growth Factor. Rev. 2015, 26, 75–82. [Google Scholar] [CrossRef]
- Brunelleschi, S.; Penengo, L.; Lavagno, L.; Santoro, C.; Colangelo, D.; Viano, I.; Gaudino, G. Macrophage stimulating protein (MSP) evokes superoxide anion production by human macrophages of different origin. Br. J. Pharmacol. 2001, 134, 1285–1295. [Google Scholar] [CrossRef]
- Wang, T.; Chen, X.; Zhang, W.; Xiang, X.; Leng, C.; Jia, Q. Roles of macrophage stimulating protein and tyrosine kinase receptor RON in smoke-induced airway inflammation of rats. Int. J. Clin. Exp. Pathol. 2015, 8, 8797–8808. [Google Scholar]
- Yu, S.M.; Kim, S.J. The thymoquinone-induced production of reactive oxygen species promotes dedifferentiation through the ERK pathway and inflammation through the p38 and PI3K pathways in rabbit articular chondrocytes. Int. J. Mol. Med. 2015, 35, 325–332. [Google Scholar] [CrossRef]
- Wang, A.Y.; Lian, L.H.; Jiang, Y.Z.; Wu, Y.L.; Nan, J.X. Gentiana manshurica Kitagawa prevents acetaminophen-induced acute hepatic injury in mice via inhibiting JNK/ERK MAPK pathway. World J. Gastroenterol. 2010, 16, 384–391. [Google Scholar] [CrossRef]
- Leng, J.; Wang, Z.; Fu, C.L.; Zhang, J.; Ren, S.; Hu, J.N.; Jiang, S.; Wang, Y.P.; Chen, C.; Li, W. NF-kappaB and AMPK/PI3K/Akt signaling pathways are involved in the protective effects of Platycodon grandiflorum saponins against acetaminophen-induced acute hepatotoxicity in mice. Phytother. Res. 2018, 32, 2235–2246. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, S.; Zhou, H.; Shen, G.; Gan, X.; Zhou, S.; Qiu, J.; Shi, C.; Lu, L. Hyperglycemia exacerbates acetaminophen-induced acute liver injury by promoting liver-resident macrophage proinflammatory response via AMPK/PI3K/AKT-mediated oxidative stress. Cell Death Discov. 2019, 5, 119. [Google Scholar] [CrossRef]
- Wojtyniak, K.; Szymanski, M.; Matlawska, I. Leonurus cardiaca L. (motherwort): A review of its phytochemistry and pharmacology. Phytother. Res. 2013, 27, 1115–1120. [Google Scholar] [CrossRef]
- Zhao, L.; Wu, D.; Sang, M.; Xu, Y.; Liu, Z.; Wu, Q. Stachydrine ameliorates isoproterenol-induced cardiac hypertrophy and fibrosis by suppressing inflammation and oxidative stress through inhibiting NF-kappaB and JAK/STAT signaling pathways in rats. Int. Immunopharmacol. 2017, 48, 102–109. [Google Scholar] [CrossRef]
- Miao, M.; Wang, T.; Lou, X.; MingBai; Xi, P.; Liu, B.; Chang, B. The influence of stachydrine hydrochloride on the reperfusion model of mice with repetitive cerebral ischemia. Saudi J. Biol. Sci. 2017, 24, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.T.; Chen, H.H.; Dong, Z.; Xu, Y.W.; Zhao, P.; Guo, W.; Wei, H.C.; Zhang, C.; Lu, R. Stachydrine Protects Against Pressure Overload-Induced Cardiac Hypertrophy by Suppressing Autophagy. Cell Physiol. Biochem. 2017, 42, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, A.; Wu, Y.; Guan, W.; Xiong, B.; Peng, X.; Wei, X.; Chen, C.; Liu, Z. Stachydrine ameliorates carbon tetrachloride-induced hepatic fibrosis by inhibiting inflammation, oxidative stress and regulating MMPs/TIMPs system in rats. Biomed. Pharmacother. 2018, 97, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.; Babar, A.; Choudhary, M.; Kutner, M.; Pyrsopoulos, N. Acetaminophen-Induced Hepatotoxicity: A Comprehensive Update. J. Clin. Transl. Hepatol. 2016, 4, 131–142. [Google Scholar]
- Fisher, J.E.; McKenzie, T.J.; Lillegard, J.B.; Yu, Y.; Juskewitch, J.E.; Nedredal, G.I.; Brunn, G.J.; Yi, E.S.; Malhi, H.; Smyrk, T.C.; et al. Role of Kupffer cells and toll-like receptor 4 in acetaminophen-induced acute liver failure. J. Surg. Res. 2013, 180, 147–155. [Google Scholar] [CrossRef]
- Holt, M.P.; Cheng, L.; Ju, C. Identification and characterization of infiltrating macrophages in acetaminophen-induced liver injury. J. Leukoc. Biol. 2008, 84, 1410–1421. [Google Scholar] [CrossRef]
- Zimmermann, H.W.; Trautwein, C.; Tacke, F. Functional role of monocytes and macrophages for the inflammatory response in acute liver injury. Front. Physiol. 2012, 3, 56. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A. Regulation of Macrophage Polarization by RON Receptor Tyrosine Kinase Signaling. Front. Immunol. 2014, 5, 546. [Google Scholar] [CrossRef]
- Li, J.; Chanda, D.; van Gorp, P.J.; Jeurissen, M.L.; Houben, T.; Walenbergh, S.M.; Debets, J.; Oligschlaeger, Y.; Gijbels, M.J.; Neumann, D.; et al. Macrophage Stimulating Protein Enhances Hepatic Inflammation in a NASH Model. PLoS ONE 2016, 11, e0163843. [Google Scholar] [CrossRef]
- Leonis, M.A.; Toney-Earley, K.; Degen, S.J.; Waltz, S.E. Deletion of the Ron receptor tyrosine kinase domain in mice provides protection from endotoxin-induced acute liver failure. Hepatology 2002, 36, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Chanda, D.; Li, J.; Oligschlaeger, Y.; Jeurissen, M.L.J.; Houben, T.; Walenbergh, S.M.A.; Shiri-Sverdlov, R.; Neumann, D. MSP is a negative regulator of inflammation and lipogenesis in ex vivo models of non-alcoholic steatohepatitis. Exp. Mol. Med. 2016, 48, e258. [Google Scholar] [CrossRef]
- Stuart, W.D.; Kulkarni, R.M.; Gray, J.K.; Vasiliauskas, J.; Leonis, M.A.; Waltz, S.E. Ron receptor regulates Kupffer cell-dependent cytokine production and hepatocyte survival following endotoxin exposure in mice. Hepatology 2011, 53, 1618–1628. [Google Scholar] [CrossRef]
- Gunella, G.; Bardelli, C.; Amoruso, A.; Viano, I.; Balbo, P.; Brunelleschi, S. Macrophage-stimulating protein differently affects human alveolar macrophages from smoker and non-smoker patients: Evaluation of respiratory burst, cytokine release and NF-kappaB pathway. Br. J. Pharmacol. 2006, 148, 478–489. [Google Scholar] [CrossRef]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [PubMed]
- Wancket, L.M.; Meng, X.; Rogers, L.K.; Liu, Y. Mitogen-activated protein kinase phosphatase (Mkp)-1 protects mice against acetaminophen-induced hepatic injury. Toxicol. Pathol. 2012, 40, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.R.; Kim, Y.H.; Hwang, J.H.; Gang, G.T.; Kim, K.S.; Lee, I.K.; Yun, B.S.; Lee, C.H. Davallialactone protects against acetaminophen overdose-induced liver injuries in mice. Food Chem. Toxicol. 2013, 58, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.U.; Stamper, B.D. Polyphenols reported to shift APAP-induced changes in MAPK signaling and toxicity outcomes. Chem. Biol. Interact. 2017, 277, 129–136. [Google Scholar] [CrossRef]
- Wang, M.; Shu, Z.J.; Wang, Y.; Peng, W. Stachydrine hydrochloride inhibits proliferation and induces apoptosis of breast cancer cells via inhibition of Akt and ERK pathways. Am. J. Transl. Res. 2017, 9, 1834–1844. [Google Scholar]
- Cianciulli, A.; Calvello, R.; Porro, C.; Trotta, T.; Salvatore, R.; Panaro, M.A. PI3k/Akt signalling pathway plays a crucial role in the anti-inflammatory effects of curcumin in LPS-activated microglia. Int. Immunopharmacol. 2016, 36, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L.; Ozment-Skelton, T.; Li, C. Modulation of the phosphoinositide 3-kinase signaling pathway alters host response to sepsis, inflammation, and ischemia/reperfusion injury. Shock 2006, 25, 432–439. [Google Scholar] [CrossRef]
- Zhao, M.; Li, C.; Shen, F.; Wang, M.; Jia, N.; Wang, C. Naringenin ameliorates LPS-induced acute lung injury through its anti-oxidative and anti-inflammatory activity and by inhibition of the PI3K/AKT pathway. Exp. Ther. Med. 2017, 14, 2228–2234. [Google Scholar] [CrossRef]
- Bai, D.; Zhao, Y.; Zhu, Q.; Zhou, Y.; Zhao, Y.; Zhang, T.; Guo, Q.; Lu, N. LZ205, a newly synthesized flavonoid compound, exerts anti-inflammatory effect by inhibiting M1 macrophage polarization through regulating PI3K/AKT/mTOR signaling pathway. Exp. Cell Res. 2018, 364, 84–94. [Google Scholar] [CrossRef]
- Yu, N.; Hu, S.; Hao, Z. Benificial Effect of Stachydrine on the Traumatic Brain Injury Induced Neurodegeneration by Attenuating the Expressions of Akt/mTOR/PI3K and TLR4/NFkappa-B Pathway. Transl. Neurosci. 2018, 9, 175–182. [Google Scholar] [CrossRef]
- Gum, S.I.; Cho, M.K. Recent updates on acetaminophen hepatotoxicity: The role of nrf2 in hepatoprotection. Toxicol. Res. 2013, 29, 165–172. [Google Scholar] [CrossRef]
- Yan, H.; Huang, Z.; Bai, Q.; Sheng, Y.; Hao, Z.; Wang, Z.; Ji, L. Natural product andrographolide alleviated APAP-induced liver fibrosis by activating Nrf2 antioxidant pathway. Toxicology 2018, 396–397, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wu, Y.; Zhang, X.; Gu, W.; Ning, Z. Stachydrine ameliorates hypoxia reoxygenation injury of cardiomyocyte via enhancing SIRT1-Nrf2 pathway. J. Cardiothorac. Surg. 2023, 18, 265. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.-C.; Yu, H.-P.; Lee, H.-C.; Chen, C.-Y.; Liao, C.-C. The Modulation of Phospho-Extracellular Signal-Regulated Kinase and Phospho-Protein Kinase B Signaling Pathways plus Activity of Macrophage-Stimulating Protein Contribute to the Protective Effect of Stachydrine on Acetaminophen-Induced Liver Injury. Int. J. Mol. Sci. 2024, 25, 1484. https://doi.org/10.3390/ijms25031484
Liu F-C, Yu H-P, Lee H-C, Chen C-Y, Liao C-C. The Modulation of Phospho-Extracellular Signal-Regulated Kinase and Phospho-Protein Kinase B Signaling Pathways plus Activity of Macrophage-Stimulating Protein Contribute to the Protective Effect of Stachydrine on Acetaminophen-Induced Liver Injury. International Journal of Molecular Sciences. 2024; 25(3):1484. https://doi.org/10.3390/ijms25031484
Chicago/Turabian StyleLiu, Fu-Chao, Huang-Ping Yu, Hung-Chen Lee, Chun-Yu Chen, and Chia-Chih Liao. 2024. "The Modulation of Phospho-Extracellular Signal-Regulated Kinase and Phospho-Protein Kinase B Signaling Pathways plus Activity of Macrophage-Stimulating Protein Contribute to the Protective Effect of Stachydrine on Acetaminophen-Induced Liver Injury" International Journal of Molecular Sciences 25, no. 3: 1484. https://doi.org/10.3390/ijms25031484
APA StyleLiu, F.-C., Yu, H.-P., Lee, H.-C., Chen, C.-Y., & Liao, C.-C. (2024). The Modulation of Phospho-Extracellular Signal-Regulated Kinase and Phospho-Protein Kinase B Signaling Pathways plus Activity of Macrophage-Stimulating Protein Contribute to the Protective Effect of Stachydrine on Acetaminophen-Induced Liver Injury. International Journal of Molecular Sciences, 25(3), 1484. https://doi.org/10.3390/ijms25031484