Distinctive Nuclear Features of Dinoflagellates with A Particular Focus on Histone and Histone-Replacement Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. An Overview of Dinoflagellates
2. Distinctive Features of Dinokaryons
2.1. Genome Size and DNA Structure
2.2. Nucleofilaments
2.3. Transcription
2.4. Chromatin Condensation and Stabilization
2.5. Cell Division
3. Nuclear Proteins of Dinoflagellates
3.1. Non-Histone Proteins of Dinokaryon
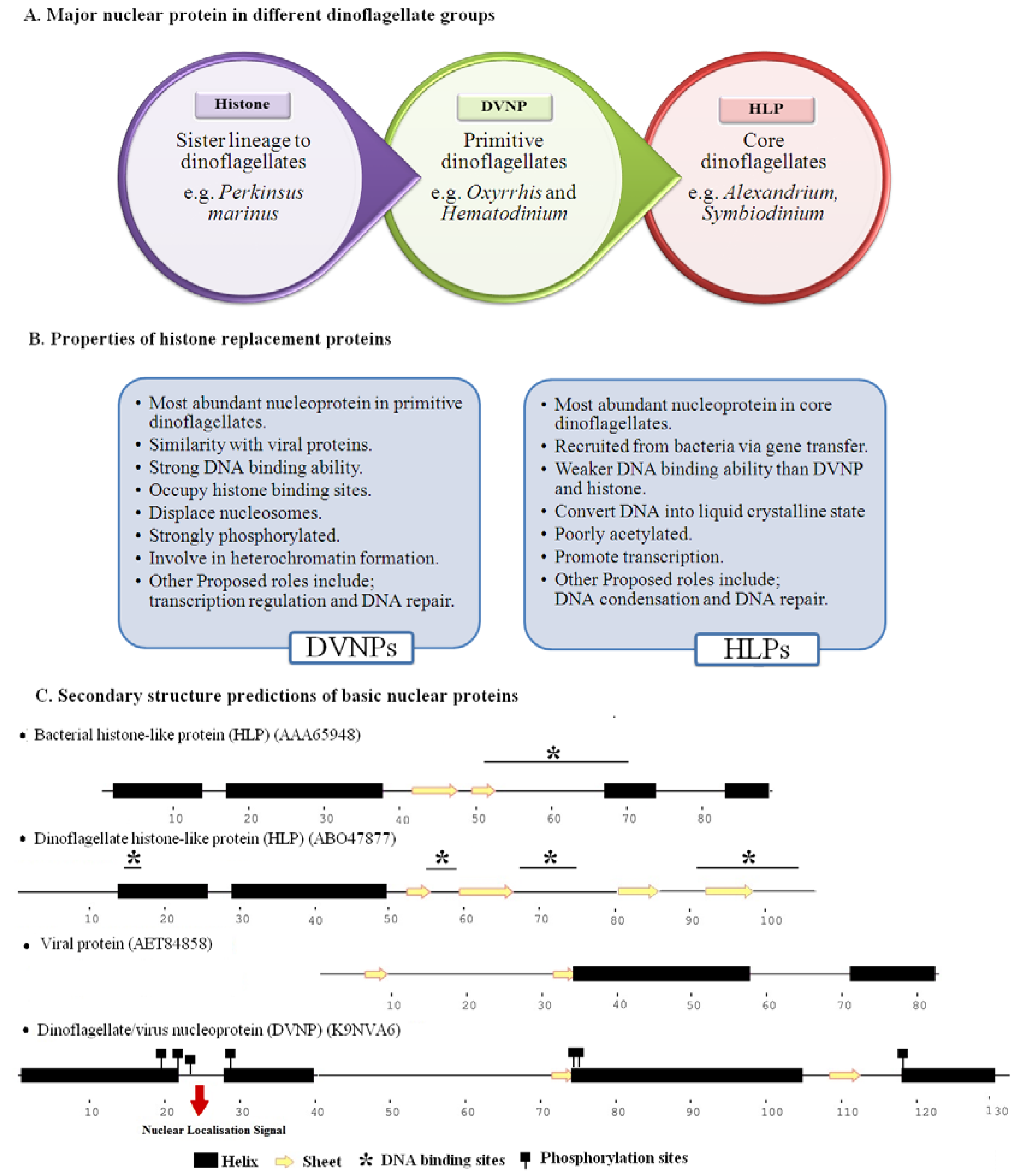
3.2. Histone and Histone-Replacement Proteins of Dinokaryons
3.2.1. Histone Like Proteins (HLPs) of C. cohnii Known as HCc
3.2.2 Histone-Like Protein-HLPs
3.2.3 Dinoflagellate/Viral Nucleoproteins-DVNPs
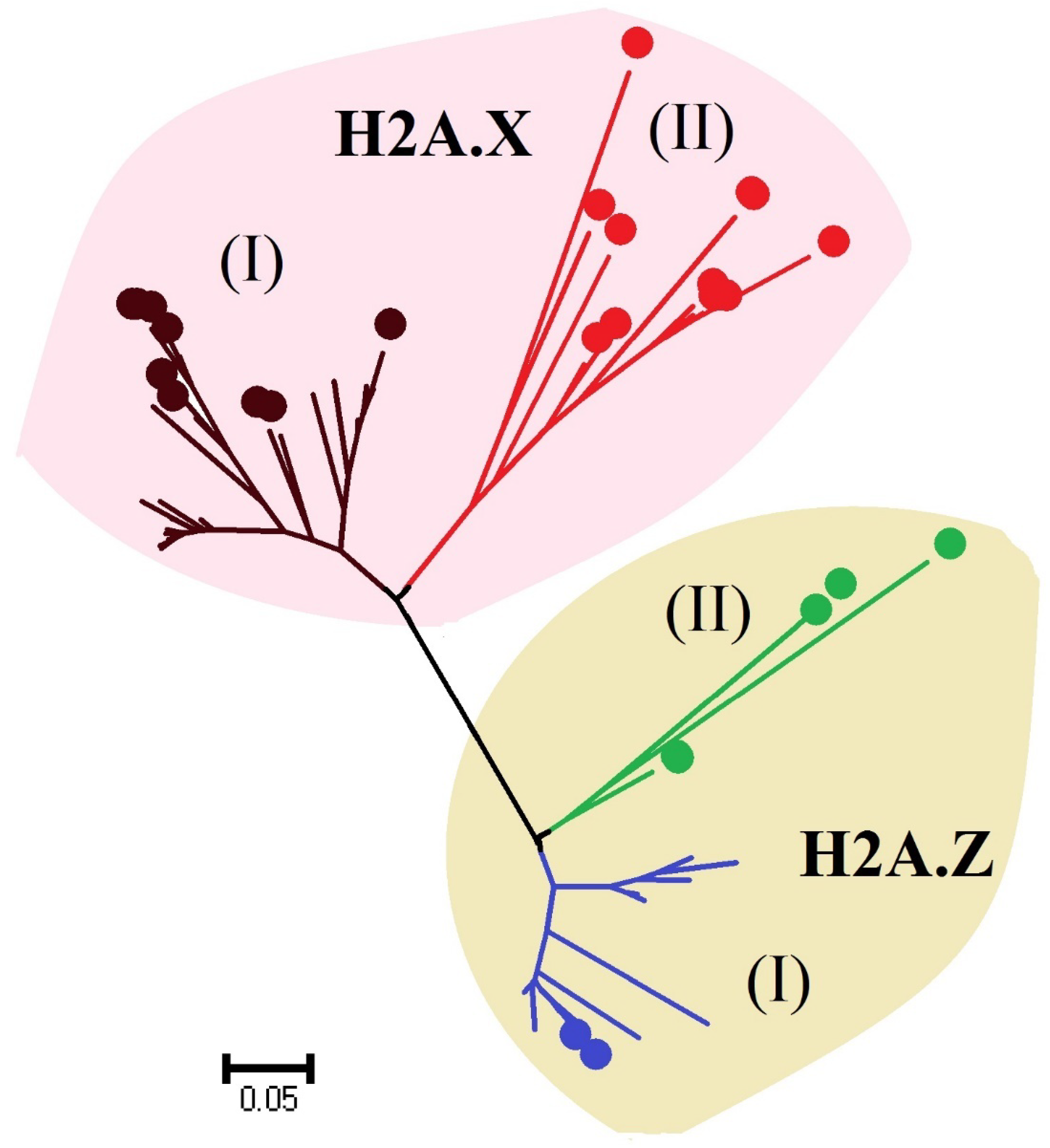
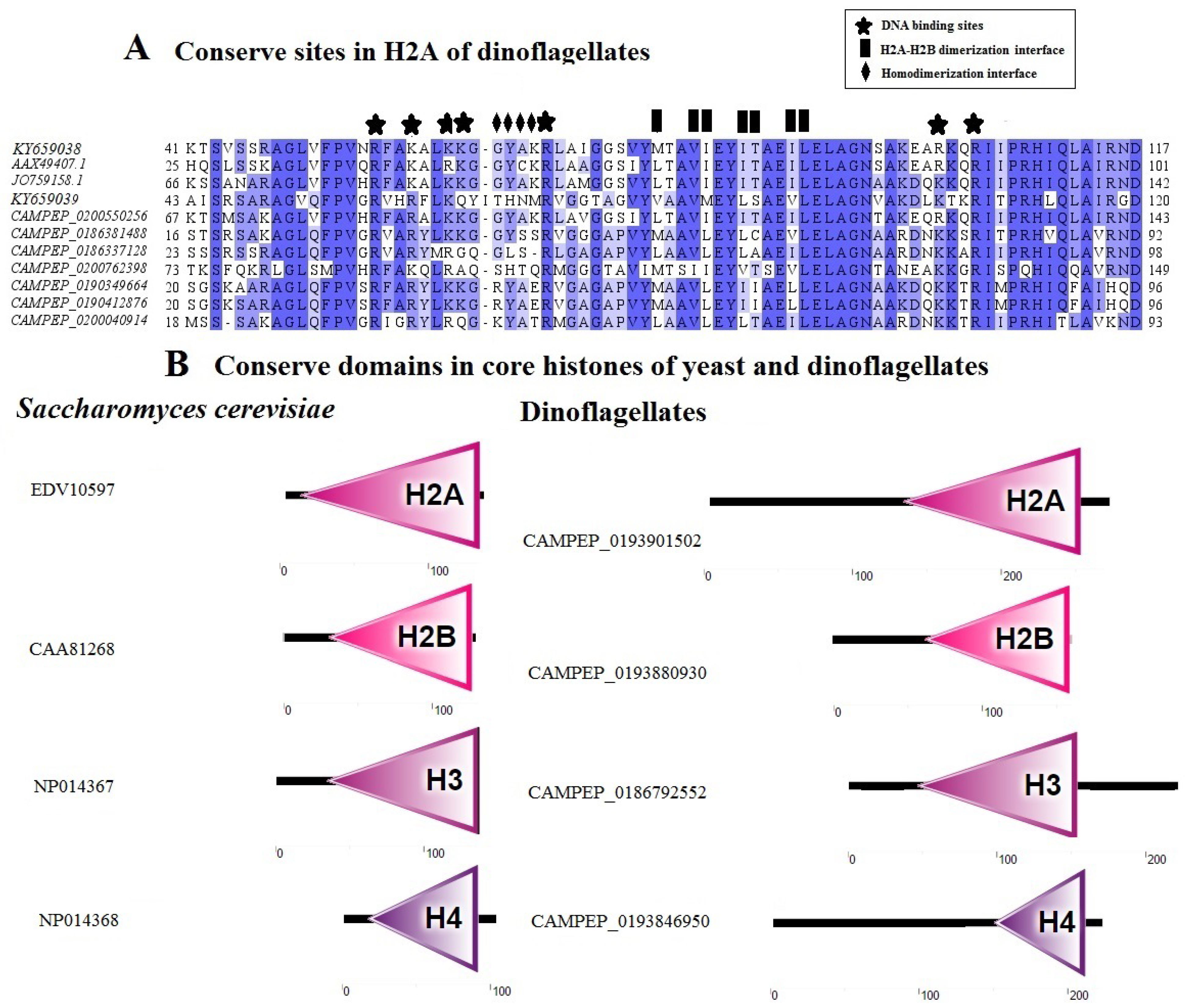
3.2.4 Histone Proteins
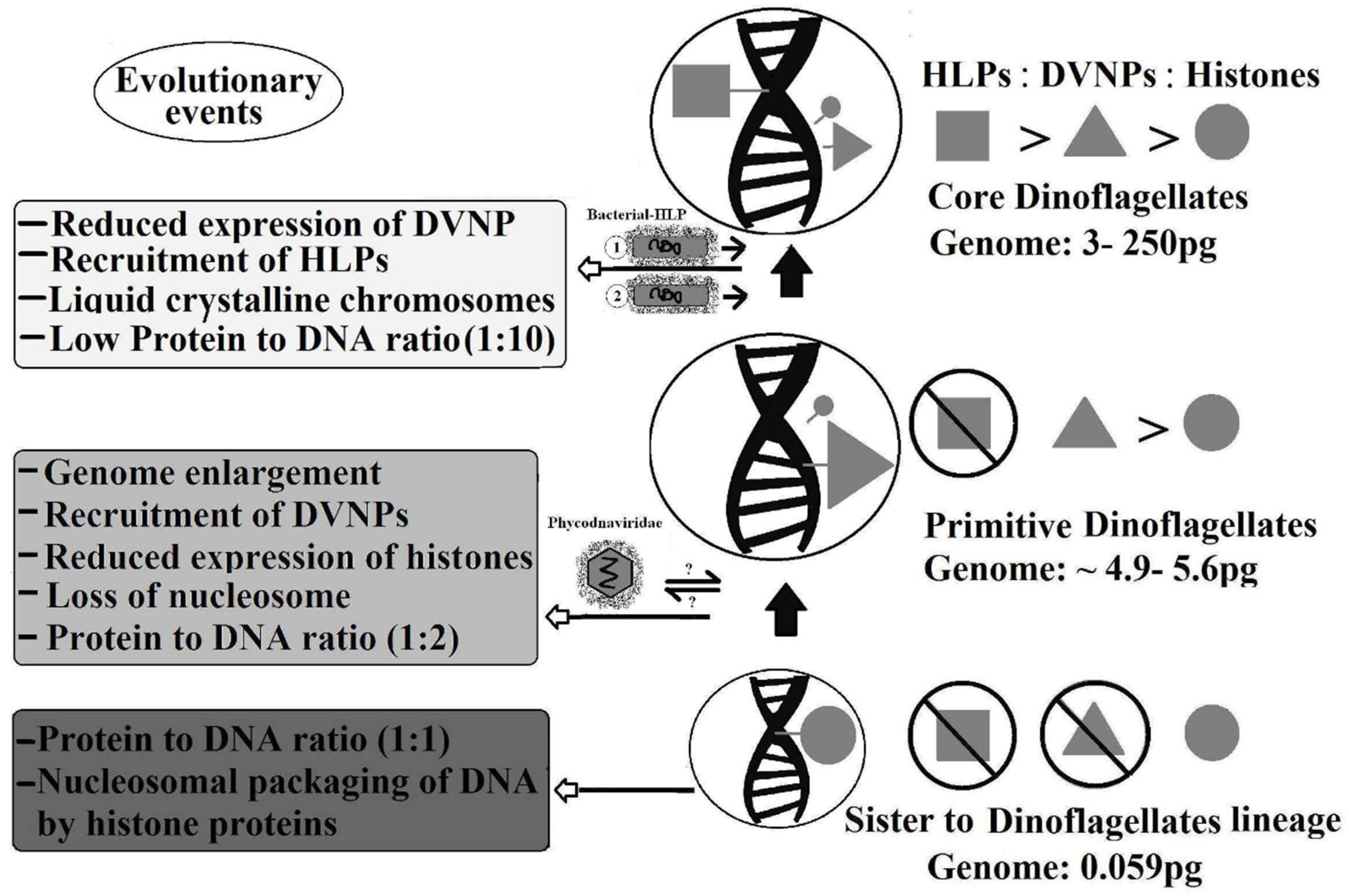
4. Evolution of Dinokaryon, with Reference to Histones and Histone-Replacement Proteins
5. Concluding Remarks
Funding
Conflicts of Interest
References
- Rizzo, P.J. The enigma of the dinoflagellate chromosome. J. Eukaryot. Microbiol. 1991, 38, 246–252. [Google Scholar] [CrossRef]
- Costas, E.; Goyanes, V. Architecture and evolution of dinoflagellate chromosomes: An enigmatic origin. Cytogenet. Genome Res. 2005, 109, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, Y.; Suzaki, T. Unusual features of dinokaryon, the enigmatic nucleus of dinoflagellates. In Marine Protists; Springer: Tokyo, Japan, 2015; pp. 23–45. [Google Scholar]
- Velo-Suárez, L.; Brosnahan, M.L.; Anderson, D.M.; McGillicuddy, J.D. A quantitative assessment of the role of the parasite Amoebophrya in the termination of Alexandrium fundyense blooms within a small coastal embayment. PLoS ONE 2013, 8, e81150. [Google Scholar]
- Moldowan, J.M.; Talyzina, N.M. Biogeochemical evidence for dinoflagellate ancestors in the Early Cambrian. Science 1998, 281, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.T.; Kwok, A. Proliferation of dinoflagellates: Blooming or bleaching. Bioessays 2005, 27, 730–740. [Google Scholar] [CrossRef]
- Morse, D.; Salois, P.; Markovic, P.; Hastings, J.W. A nuclear-encoded form II RuBisCO in dinoflagellates. Science 1995, 268, 1622. [Google Scholar] [CrossRef]
- Tang, E.P. Why do dinoflagellates have lower growth rates? J. Phycol. 1996, 32, 80–84. [Google Scholar] [CrossRef]
- Gong, W.; Browne, J.; Hall, N.; Schruth, D.; Paerl, H.; Marchetti, A. Molecular insights into a dinoflagellate bloom. ISME J. 2016, 11, 439–452. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Geng, H.; Sui, Z.; Zhang, S.; Du, Q.; Ren, Y.; Liu, Y.; Kong, F.; Zhong, J.; Ma, Q. Identification of microRNAs in the Toxigenic Dinoflagellate Alexandrium catenella by High-Throughput Illumina Sequencing and Bioinformatic Analysis. PLoS ONE 2015, 10, e0138709. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E. Bivalve Molluscs as Vectors of Marine Biotoxins Involved in Seafood Poisoning. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 53–82. [Google Scholar]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J.R. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J. Diversity and evolutionary history of plastids and their hosts. Am. J. Bot. 2004, 91, 1481–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.J.; Norman, J.E.; Schnare, M.N.; Gray, M.W.; Keeling, P.J.; Waller, R.F. Broad genomic and transcriptional analysis reveals a highly derived genome in dinoflagellate mitochondria. BMC Biol. 2007, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.F.; Jackson, C.J. Dinoflagellate mitochondrial genomes: Stretching the rules of molecular biology. Bioessays 2009, 31, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Spector, D.L. Dinoflagellate nuclei. Dinoflagellates 1984, 1, 107–147. [Google Scholar]
- De la Espina, S.M.D.; Alverca, E.; Cuadrado, A.; Franca, S. Organization of the genome and gene expression in a nuclear environment lacking histones and nucleosomes: The amazing dinoflagellates. Eur. J. Cell Biol. 2005, 84, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, P.J. Those amazing dinoflagellate chromosomes. Cell Res. 2003, 13, 215–217. [Google Scholar] [CrossRef] [Green Version]
- Holm-Hansen, O. Algae: Amounts of DNA and organic carbon in single cells. Science 1969, 163, 87–88. [Google Scholar] [CrossRef]
- Blank, R.J.; Huss, V.A.; Kersten, W. Base composition of DNA from symbiotic dinoflagellates: A tool for phylogenetic classification. Arch. Microbiol. 1988, 149, 515–520. [Google Scholar] [CrossRef]
- Rae, P.M. 5-Hydroxymethyluracil in the DNA of a dinoflagellate. Proc. Natl. Acad. Sci. USA 1973, 70, 1141–1145. [Google Scholar] [CrossRef]
- Rae, P. Hydroxymethyluracil in eukaryote DNA: A natural feature of the pyrrophyta (dinoflagellates). Science 1976, 194, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Rae, P.M.; Steele, R.E. Modified bases in the DNAs of unicellular eukaryotes: An examination of distributions and possible roles, with emphasis on hydroxymethyluracil in dinoflagellates. Biosystems 1978, 10, 37–53. [Google Scholar] [CrossRef]
- Michael, R.; Miller, D.J. Light-Regulated Transcription of Genes Encoding Peridinin Chlorophyll a Proteins and the Major Intrinsic Light-Harvesting Complex Proteins in the DinoflagellateAmphidinium carterae Hulburt (Dinophycae) Changes in Cytosine Methylation Accompany Photoadaptation. Plant Physiol. 1998, 117, 189–196. [Google Scholar]
- McEwan, M.; Humayun, R.; Slamovits, C.H.; Keeling, P.J. Nuclear genome sequence survey of the dinoflagellate Heterocapsa triquetra. J. Euk. Microbiol. 2008, 55, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Shoguchi, E.; Shinzato, C.; Kawashima, T.; Gyoja, F.; Mungpakdee, S.; Koyanagi, R.; Takeuchi, T.; Hisata, K.; Tanaka, M.; Fujiwara, M.; et al. Draft assembly of the Symbiodinium minutum nuclear genome reveals dinoflagellate gene structure. Curr. Biol. 2013, 23, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lin, S. Complex gene structure of the form II RUBISCO in the dinoflagellate Prorocentrum minimum (Dinophyceae). J. Phycol. 2003, 39, 1160–1171. [Google Scholar] [CrossRef]
- Rowan, R.; Whitney, S.M.; Fowler, A.; Yellowlees, D. Rubisco in marine symbiotic dinoflagellates: Form II enzymes in eukaryotic oxygenic phototrophs encoded by a nuclear multigene family. Plant Cell. 1996, 8, 539–553. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Leander, B.S. Dinoflagellate phylogeny as inferred from heat shock protein 90 and ribosomal gene sequences. PLoS ONE 2010, 5, e13220. [Google Scholar] [CrossRef]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative genomic and transcriptomic characterization of the toxigenic marine dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef]
- Lin, S.; Cheng, S.; Song, B.; Zhong, X.; Lin, X.; Li, W.; Li, L.; Zhang, Y.; Zhang, H.; Ji, Z. The symbiodinium kawagutii genome illuminates dinoflagellate gene expression and coral symbiosis. Science 2015, 350, 691–694. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Sebbo, J.; Feist-Burkhardt, S.; Irwin, A.; Katz, M.; Schofield, O.; Young, J.R.; Falkowski, P.G. A universal driver of macroevolutionary change in the size of marine phytoplankton over the Cenozoic. Proc. Natl. Acad. Sci. USA 2007, 104, 20416–20420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, J.A.; Oliver, M.J.; Beaulieu, J.M.; Knight, C.A.; Tomanek, L.; Moline, M.A. Correlated evolution of genome size and cell volume in diatoms (Bacillariophyceae). J. Phycol. 2008, 44, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Irwin, A.J.; Finkel, Z.V.; Schofield, O.M.E.; Falkowski, P.G. Scaling-up from nutrient physiology to the size-structure of phytoplankton communities. J. Plankton Res. 2006, 28, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Talbert, P.B.; Henikoff, S. Chromatin: Packaging without nucleosomes. Curr. Biol. 2012, 22, R1040–R1043. [Google Scholar] [CrossRef] [PubMed]
- Gornik, S.G.; Ford, K.L.; Mulhern, T.D.; Bacic, A.; McFadden, G.I.; Waller, R.F. Loss of nucleosomal DNA condensation coincides with appearance of a novel nuclear protein in dinoflagellates. Curr. Biol. 2012, 22, 2303–2312. [Google Scholar] [CrossRef] [PubMed]
- Herzog, M.; Soyer, M. Distinctive features of dinoflagellate chromatin. Absence of nucleosomes in a primitive species Prorocentrum micans E. Eur. J Cell Biol. 1981, 23, 295–302. [Google Scholar] [PubMed]
- Bodansky, S.; Mintz, L.B.; Holmes, D.S. The mesokaryote Gyrodiniumcohnii lacks nucleosomes. Biochem. Biophys. Res. Commun. 1979, 88, 1329–1336. [Google Scholar] [CrossRef]
- Rill, R.L.; Livolant, F.; Aldrich, H.C.; Davidson, M.W. Electron microscopy of liquid crystalline DNA: Direct evidence for cholesteric-like organization of DNA in dinoflagellate chromosomes. Chromosoma 1989, 98, 280–286. [Google Scholar] [CrossRef]
- Sigee, D.C. Structural DNA and genetically active DNA in dinoflagellate chromosomes. Biosystems. 1983, 16, 203–210. [Google Scholar] [CrossRef]
- Soyer-Gobillard, M.O.; Géraud, M.L.; Coulaud, D.; Barray, M.; Théveny, B.; Révet, B.; Delain, E. Location of B-and Z-DNA in the chromosomes of a primitive eukaryote dinoflagellate. J. Cell Biol. 1990, 111, 293–304. [Google Scholar] [CrossRef]
- Zhang, H.; Hou, Y.; Miranda, L.; Campbell, D.A.; Sturm, N.R.; Gaasterland, T.; Lin, S. Spliced leader RNA trans-splicing in dinoflagellates. Proc. Natl. Acad. Sci. USA 2007, 104, 4618–4623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slamovits, C.H.; Keeling, P.J. Widespread recycling of processed cDNAs in dinoflagellates. Curr. Biol. 2008, 18, R550–R552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slamovits, C.H.; Keeling, P.J. Contributions of Oxyrrhis marina to molecular biology, genomics and organelle evolution of dinoflagellates. J. Plankton Res. 2010, 33, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lin, S. Retrieval of missing spliced leader in dinoflagellates. PLoS ONE 2009, 4, e4129. [Google Scholar] [CrossRef]
- Hastings, K.E. SL trans-splicing: Easy come or easy go? Trends Genet. 2005, 21, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, H.; Zhuang, Y.; Tran, B.; Gill, J. Spliced leader–based metatranscriptomic analyses lead to recognition of hidden genomic features in dinoflagellates. Proc. Natl. Acad. Sci. USA 2010, 107, 20033–20038. [Google Scholar] [CrossRef] [PubMed]
- Lin, S. Genomic understanding of dinoflagellates. Res. Microbiol. 2011, 162, 551–569. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhang, H.; Lin, S. Polyadenylation of 18S rRNA in algae. J. Phycol. 2013, 49, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Guillebault, D.; Sasorith, S.; Derelle, E.; Wurtz, J.M.; Lozano, J.C.; Bingham, S.; Tora, L.; Moreau, H. A new class of transcription initiation factors, intermediate between TATA box-binding proteins (TBPs) and TBP-like factors (TLFs), is present in the marine unicellular organism, the dinoflagellate Crypthecodinium cohnii. J. Biol. Chem. 2002, 277, 40881–40886. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.K. Dinoflagellate viral nucleoproteins. Master’s Thesis, The Hong Kong university of science and technology, Hong Kong, China, 2014. [Google Scholar]
- Erdner, D.L.; Anderson, D.M. Global transcriptional profiling of the toxic dinoflagellate Alexandrium fundyense using massively parallel signature sequencing. BMC Genomics. 2006, 7, 88. [Google Scholar] [CrossRef]
- Moustafa, A.; Evans, A.N.; Kulis, D.M.; Hackett, J.D.; Erdner, D.L.; Anderson, D.M.; Bhattacharya, D. Transcriptome profiling of a toxic dinoflagellate reveals a gene-rich protist and a potential impact on gene expression due to bacterial presence. PLoS ONE 2010, 5, e9688. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, M.; Morse, D. A proteomic portrait of dinoflagellate chromatin reveals abundant RNA-binding proteins. Chromosoma 2017, 127, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, O.K.; Hastings, J.W. Genome-wide analysis of redox-regulated genes in a dinoflagellate. Gene 2003, 321, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, O.K.; Hastings, J. Novel dinoflagellate clock-related genes identified through microarray analysis. J. Phycol. 2003, 39, 519–526. [Google Scholar] [CrossRef]
- Van Dolah, F.M.; Lidie, K.B.; Morey, J.S.; Brunelle, S.A.; Ryan, J.C.; Monroe, E.A.; Haynes, B. Microarray analysis of diurnal-and circadian-regulated genes in the florida red-tide dinoflagellate karenia brevis (dinophyceae). J. Phycol. 2007, 43, 741–752. [Google Scholar] [CrossRef]
- Monroe, E.A.; Johnson, J.G.; Wang, Z.; Pierce, R.K.; Van Dolah, F.M. Characterization and expression of nuclear-encoded polyketide synthases in the brevetoxin-producing dinoflagellate karenia brevis. J. Phycol. 2010, 46, 541–552. [Google Scholar] [CrossRef]
- Yang, I.; John, U.; Beszteri, S.; Glöckner, G.; Krock, B.; Goesmann, A.; Cembella, A.D. Comparative gene expression in toxic versus non-toxic strains of the marine dinoflagellate Alexandrium minutum. BMC Genomics 2010, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Aranda, M.; Li, Y.; Liew, Y.J.; Baumgarten, S.; Simakov, O.; Wilson, M.C.; Piel, J.; Ashoor, H.; Bougouffa, V.B.; Bajic, T.; et al. Genomes of coral dinoflagellate symbionts highlight evolutionary adaptations conducive to a symbiotic lifestyle. Sci. Rep. 2016, 6, 639734. [Google Scholar] [CrossRef] [PubMed]
- Wisecaver, J.H.; Hackett, J.D. Dinoflagellate genome evolution. Annu. Rev. Microbiol. 2011, 65, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Strzelecka, T.E.; Davidson, M.W.; Rill, R.L. Multiple liquid crystal phases of DNA at high concentrations. Nature 1988, 331, 457–460. [Google Scholar] [CrossRef] [Green Version]
- Bendet, I.J.; Goldstein, D.A.; Lauffer, M.A. Evidence for internal organization of nucleic acid in T2 bacteriophage. Nature 1960, 187, 781–782. [Google Scholar] [CrossRef] [PubMed]
- Reich, Z.; Wachtel, E.J.; Minsky, A. Liquid-crystalline mesophases of plasmid DNA in bacteria. Science 1994, 264, 1460–1462. [Google Scholar] [CrossRef] [PubMed]
- Kierszenbaum, A.; Tres, L.L. Structural and transcriptional features of the mouse spermatid genome. J. Cell Biol. 1975, 65, 258–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouligand, Y.; Soyer, M.; Puiseux-Dao, S. The fibrillary structure and orientation of chromosomes in dinoflagellata. Chromosoma 1968, 24, 251. [Google Scholar] [CrossRef] [PubMed]
- Kearns, L.; Sigee, D. High levels of transition metals in dinoflagellate chromosomes. Cell. Mol. Life Sci. 1979, 35, 1332–1334. [Google Scholar] [CrossRef]
- Soyer, M.-O.; Haapala, O. Structural changes of dinoflagellate chromosomes by pronase and ribonuclease. Chromosoma 1974, 47, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Soyer-Gobillard, M.-O.; Ausseil, J.; Géraud, M.-L. Nuclear and cytoplasmic actin in dinoflagellates. Biol. Cell 1996, 87, 17–35. [Google Scholar] [CrossRef]
- Mak, C.K.; Hung, V.K.; Wong, J.T. Type ii topoisomerase activities in both the g1 and g2/m phases of the dinoflagellate cell cycle. Chromosoma 2005, 114, 420–431, Erratum in 2006, 115, 341. [Google Scholar] [CrossRef]
- Zhang, J. Dinoflagellate histone-like proteins. Ph.D. Thesis, The Hong Kong university of science and technology, Hong Kong, China, 2014. [Google Scholar]
- Shi, X.; Ma, M.; Lin, S. Cell cycle-dependent expression dynamics of g1/s specific cyclin, cellulose synthase and cellulase in the dinoflagellate prorocentrum donghaiense. Front. Microbiol. 2017, 8, 1118. [Google Scholar] [CrossRef]
- Hastings, J. Chemistry, clones, and circadian control of the dinoflagellate bioluminescent system. The marlene deluca memorial lecture. Luminescence 1989, 4, 12–19. [Google Scholar] [CrossRef]
- Yeung, P.; Wong, J. Inhibition of cell proliferation by mechanical agitation involves transient cell cycle arrest at g 1 phase in dinoflagellates. Protoplasma 2003, 220, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Bhaud, Y.; Guillebault, D.; Lennon, J.; Defacque, H.; Soyer-Gobillard, M.-O.; Moreau, H. Morphology and behaviour of dinoflagellate chromosomes during the cell cycle and mitosis. J. Cell Sci. 2000, 113, 1231–1239. [Google Scholar] [PubMed]
- Soyer-Gobillard, M.-O.; Gillet, B.; Géraud, M.-L.; Bhaud, Y. Dinoflagellate chromosome behaviour during stages of replication. Int. Microbiol. 1999, 2, 93–102. [Google Scholar] [PubMed]
- Holck, A.; Lossius, I.; Aasland, R.; Haarr, L.; Kleppe, K. DNA-and rna-binding proteins of chromatin from escherichia coli. Biochim. Biophys. 1987, 908, 188–199. [Google Scholar] [CrossRef]
- Franker, C.K. DNA-binding protein and the cell cycle in cryptothecodinium cohnii. I. On the resolution of metabolically stable components. J. Phycol. 1972, 8, 264–268. [Google Scholar] [CrossRef]
- Franker, C.; Prichard, C.D.; Lamden, C.A. DNA-binding protein and the cell cycle in cryptothecodinium cohnii: 2. Short-lived protein with affinity for double-stranded bulk DNA. Differentiation 1973, 1, 383–391. [Google Scholar] [CrossRef]
- Bhaud, Y.; Géraud, M.l.; Ausseil, J.; Soyer-Gobillard, M.O.; Moreau, H. Cyclic expression of a nuclear protein in a dinoflagellate. J. Eukaryot. Microbiol. 1999, 46, 259–267. [Google Scholar] [CrossRef]
- Mínguez, A.; Franca, S.; De La Espina, S.M.D. Dinoflagellates have a eukaryotic nuclear matrix with lamin-like proteins and topoisomerase ii. J. Cell Sci. 1994, 107, 2861–2873. [Google Scholar]
- Zhang, H.; Hou, Y.; Lin, S. Isolation and characterization of proliferating cell nuclear antigen from the dinoflagellate pfiesteria piscicida. J. Eukaryot. Microbiol. 2006, 53, 142–150. [Google Scholar] [CrossRef]
- Riaz, S.; Sui, Z.; Niaz, Z; Yuan, L.; Khan, S. Detection, characterization and expression dynamics of histone proteins in the dinoflagellate Alexandrium pacificum during growth regulation. Microorganisms 2018. (Under review). [Google Scholar]
- Chan, Y.H.; Wong, J.T. Concentration-dependent organization of DNA by the dinoflagellate histone-like protein HCc3. Nucleic Acids Res. 2007, 35, 2573–2583. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.; Kwok, A.; Tsang, J.S.; Wong, J.T. Alveolata histone-like proteins have different evolutionary origins. J. Evol. Biol. 2006, 19, 1717–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chudnovsky, Y.; Li, J.F.; Rizzo, P.J.; Hastings, J.; Fagan, T.F. Cloning, expression, and characterization of a histone-like protein from the marine dinoflagellate lingulodinium polyedrum (dinophyceae) 1. J. Phycol. 2002, 38, 543–550. [Google Scholar] [CrossRef]
- Wargo, M.J.; Rizzo, P.J. Characterization of Gymnodinium Mikimotoi (dinophyceae) nuclei and identification of the major histone-like protein, hgm. J. Phycol. 2000, 36, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Marinov, G.K.; Lynch, M. Diversity and divergence of dinoflagellate histone proteins. G3 (Bethesda) 2016, 6, 397–422. [Google Scholar] [CrossRef] [PubMed]
- Ris, H. Interpretation of ultrastructure in the cell nucleus. In The interpretation of ultrastructure; Elsevier: Amsterdam, The Netherlands, 1962; pp. 69–88. [Google Scholar]
- Dodge, J.D. Chromosome structure in the dinophyceae. Arch. Microbiol 1964, 48, 66–80. [Google Scholar] [CrossRef]
- Stewart, J.M.; BECK, J.S. Distribution of the DNA and the DNA-histone antigens in the nuclei of free-living and parasitic sarcomastigophora. J. Eukaryot. Microbiol. 1967, 14, 225–231. [Google Scholar] [CrossRef]
- Rizzo, P.J.; Noodén, L.D. Chromosomal proteins in the dinoflagellate alga gyrodinium cohnii. Science 1972, 176, 796–797. [Google Scholar] [CrossRef]
- Rizzo, P.; Morris, R. Some properties of the histone-like protein from crypthecodinium cohnii (hcc). Biosystems 1983, 16, 211–216. [Google Scholar] [CrossRef]
- Rizzo, P.; Nooden, L. Isolation and partial characterization of dinoflagellate chromatin. Biochim. Biophys. 1974, 349, 402–414. [Google Scholar] [CrossRef]
- Rizzo, P.; Nooden, L. Partial characterization of dinoflagellate chromosomal proteins. Biochim. Biophys. 1974, 349, 415–427. [Google Scholar] [CrossRef]
- Bonner, J.; Dahmus, M.E.; Fambrough, D.; Ru-chih, C.H.; Marushige, K.; Tuan, D.Y. The biology of isolated chromatin. Science 1968, 159, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Vernet, G.; Sala-Rovira, M.; Maeder, M.; Jacques, F.; Herzog, M. Basic nuclear proteins of the histone-less eukaryote crypthecodinium cohnii (pyrrhophyta): Two-dimensional electrophoresis and DNA-binding properties. Biochim. Biophys. 1990, 1048, 281–289. [Google Scholar] [CrossRef]
- Sala-Rovira, M.; Geraud, M.; Caput, D.; Jacques, F.; Soyer-Gobillard, M.; Vernet, G.; Herzog, M. Molecular cloning and immunolocalization of two variants of the major basic nuclear protein (hcc) from the histone-less eukaryote crypthecodinium cohnii (pyrrhophyta). Chromosoma 1991, 100, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Kasinsky, H.E.; Lewis, J.D.; Dacks, J.B.; Ausio, J. Origin of H1 linker histones. FASEB J. 2001, 15, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T.; Luijsterburg, M.S.; Krin, E.; Bertin, P.N.; Wagner, R.; Wuite, G.J. DNA bridging: A property shared among H-NS-like proteins. J. Bacteriol. 2005, 187, 1845–1848. [Google Scholar] [CrossRef] [PubMed]
- Luijsterburg, M.S.; Noom, M.C.; Wuite, G.J.; Dame, R.T. The architectural role of nucleoid-associated proteins in the organization of bacterial chromatin: a molecular perspective. J. Struct. Biol. 2006, 156, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Spurio, R.; Falconi, M.; Brandi, A.; Pon, C.L.; Gualerzi, C.O. The oligomeric structure of nucleoid protein H-NS is necessary for recognition of intrinsically curved DNA and for DNA bending. EMBO J. 1997, 16, 1795–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, R.; Lurz, R.; Lüder, G.; Tolksdorf, C.; Travers, A.; Muskhelishvili, G. An architectural role of the Escherichia coli chromatin protein FIS in organising DNA. Nucleic Acids Res. 2001, 29, 5107–5114. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.T.; New, D.; Wong, J.; Hung, V. Histone-like proteins of the dinoflagellate Crypthecodinium cohnii have homologies to bacterial DNA-binding proteins. Eukaryot. Cell. 2003, 2, 646–650. [Google Scholar] [CrossRef]
- Sun, S.; Liu, M.; Dong, F.; Fan, S.; Yao, Y. A histone-like protein induces plasmid DNA to form liquid crystals in vitro and gene compaction in vivo. Int. J. Mol. Sci. 2013, 14, 23842–23857. [Google Scholar] [CrossRef]
- Javaherian, K.; Liu, L.F. Association of eukaryotic DNA topoisomerase i with nucleosomes and chromosomal proteins. Nucleic Acids Res. 1983, 11, 461–472. [Google Scholar] [CrossRef] [PubMed]
- West, A.G.; Fraser, P. Remote control of gene transcription. Hum. Mol. Genet. 2005, 14, R101–R111. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, P.J. Comparative aspects of basic chromatin proteins in dinoflagellates. Biosystems 1981, 14, 433–443. [Google Scholar] [CrossRef]
- Janouškovec, J.; Gavelis, G.S.; Burki, F.; Dinh, D.; Bachvaroff, T.R.; Gornik, S.G.; Bright, K.J.; Imanian, B.; Strom, S.L.; Delwiche, C.F.; et al. Major transitions in dinoflagellate evolution unveiled by phylotranscriptomics. Proc. Natl. Acad. Sci. USA 2017, 114, E171–E180. [Google Scholar] [CrossRef] [PubMed]
- Fagan, F.; Li, J.; Chudnovsky, J.; Rizzo, P.; Hastings, J. Cloning, sequencing and expression of a histone-like protein from the photosynthetic dinoflagellate gonyaulax polyedra. J. Phycol. 2000, 36, 21–22. [Google Scholar] [CrossRef]
- Wang, D.; Lin, L.; Wang, M.; Li, C.; Hong, H. Proteomic analysis of a toxic dinoflagellate alexandrium catenella under different growth phases and conditions. Chi. Sci. Bull. 2012, 1–14. [Google Scholar] [CrossRef]
- Jean, N.; Dumont, E.; Herzi, F.; Balliau, T.; Laabir, M.; Masseret, E.; Mounier, S. Modifications of the soluble proteome of a mediterranean strain of the invasive neurotoxic dinoflagellate alexandrium catenella under metal stress conditions. Aquat. Toxicol. 2017, 188, 80–91. [Google Scholar] [CrossRef]
- Taroncher-Oldenburg, G.; Anderson, D.M. Identification and characterization of three differentially expressed genes, encoding s-adenosylhomocysteine hydrolase, methionine aminopeptidase, and a histone-like protein, in the toxic dinoflagellate alexandrium fundyense. Appl. Environ. Microbiol. 2000, 66, 2105–2112. [Google Scholar] [CrossRef]
- Siu, G.K.; Young, M.L.; Chan, D. Environmental and nutritional factors which regulate population dynamics and toxin production in the dinoflagellate Alexandrium catenella. In Presented at the Asia-Pacific Conference on Science and Management of Coastal Environment; Springer: Dodrecht, The Netherlands, 1997; pp. 117–140. [Google Scholar]
- Taroncher-Oldenburg, G.; Kulis, D.M.; Anderson, D.M. Toxin variability during the cell cycle of the dinoflagellate alexandrium fundyense. Limnol. Oceanogr. 1997, 42, 1178–1188. [Google Scholar] [CrossRef]
- John, U.; Quilliam, M.A.; Medlin, L.; Cembella, A. Spirolide production and photoperiod-dependent growth of the marine dinoflagellate Alexandrium ostenfeldii. In Proceedings of the Harmful Algal Blooms 2000, Hobart, Australia, 7–11 February 2000. [Google Scholar]
- Zhang, S.; Sui, Z.; Chang, L.; Kang, K.; Ma, J.; Kong, F.; Zhou, W.; Wang, J.; Guo, L.; Geng, H.; et al. Transcriptome de novo assembly sequencing and analysis of the toxic dinoflagellate Alexandrium catenella using the Illumina platform. Gene 2014, 537, 285–293. [Google Scholar] [CrossRef]
- Kato, K.H.; Moriyama, A.; Huitorel, P.; Cosson, J.; Cachon, M.; Sato, H. Isolation of the major basic nuclear protein and its localization on chromosomes of the dinoflagellate, oxyrrhis marina. Biol. Cell 1997, 89, 43–52. [Google Scholar] [CrossRef]
- Lee, R.; Lai, H.; Malik, S.B.; Saldarriaga, J.F.; Keeling, P.J.; Slamovits, C.H. Analysis of est data of the marine protist oxyrrhis marina, an emerging model for alveolate biology and evolution. BMC genomics 2014, 15, 122. [Google Scholar] [CrossRef]
- Riaz, S.; Sui, Z. Molecular cloning, transcriptome profiling, and characterization of histone genes in the dinoflagellate Alexandrium pacificum. J. Microbiol. Biotechnol. 2018, 7, 1185–1198. [Google Scholar]
- Black, D.L. Mechanisms of alternative pre-messenger rna splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed]
- Jones, DT. Protein secondary structure prediction based on position-specific scoring matrices1. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Irwin, N.A.; Martin, B.J.; Young, B.P.; Browne, M.J.; Flaus, A.; Loewen, C.J.; Keeling, P.J.; Howe, L.J. Viral proteins as a potential driver of histone depletion in dinoflagellates. Nat. Commu. 2018, 9, 1535. [Google Scholar] [CrossRef]
- Kohli, G.S.; John, U.; Figueroa, R.I.; Rhodes, L.L.; Harwood, D.T.; Groth, M.; Bolch, J.S.; Murray, S.A. Polyketide synthesis genes associated with toxin production in two species of Gambierdiscus (Dinophyceae). BMC Genomics 2015, 16, 410. [Google Scholar] [CrossRef]
- Roy, S.; Morse, D. A full suite of histone and histone modifying genes are transcribed in the dinoflagellate lingulodinium. PLoS ONE 2012, 7, e34340. [Google Scholar] [CrossRef]
- Bayer, T.; Aranda, M.; Sunagawa, S.; Yum, L.K.; DeSalvo, M.K.; Lindquist, E.; Coffroth, M.A.; Voolstra, C.R.; Medina, M. Symbiodinium transcriptomes: Genome insights into the dinoflagellate symbionts of reef-building corals. PLoS ONE 2012, 7, e35269. [Google Scholar] [CrossRef]
- Ismail, I.H.; Hendzel, M.J. The γ-h2a. X: Is it just a surrogate marker of double-strand breaks or much more? Environ. Mol. Mutagen. 2008, 49, 73–82. [Google Scholar] [CrossRef]
- Hake, S.B.; Allis, C.D. Histone h3 variants and their potential role in indexing mammalian genomes: The “h3 barcode hypothesis”. Proc. Natl. Acad. Sci. USA 2006, 103, 6428–6435. [Google Scholar] [CrossRef] [PubMed]
- Otero, S.; Desvoyes, B.; Gutierrez, C. Histone h3 dynamics in plant cell cycle and development. Cytogenet. Genome Res. 2014, 143, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Burki, F.; Wilcox, H.M.; Allam, B.; Allen, E.E.; Amaral-Zettler, L.A.; Armbrust, E.V.; Archibald, J.M; Bharti, A.K.; Bell, C.J.; et al. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): Illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Biol. 2014, 12, e1001889. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2014, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glozak, M.A.; Sengupta, N.; Zhang, X.; Seto, E. Acetylation and deacetylation of non-histone proteins. Gene 2005, 363, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Frye, R.A. Phylogenetic classification of prokaryotic and eukaryotic Sir2-like proteins. Biochem. Biophys. Res. Commun. 2000, 273, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Trievel, R.C.; Beach, B.M.; Dirk, L.M.; Houtz, R.L.; Hurley, J.H. Structure and catalytic mechanism of a SET domain protein methyltransferase. Cell 2002, 111, 91–103. [Google Scholar] [CrossRef]
- Pennini, M.E.; Perrinet, S.; Dautry-Varsat, A.; Subtil, A. Histone methylation by NUE, a novel nuclear effector of the intracellular pathogen Chlamydia trachomatis. PLoS Pathog. 2010, 6, e1000995. [Google Scholar] [CrossRef]
- Meyer, J.M.; Rödelsperger, C.; Eichholz, K.; Tillmann, U.; Cembella, A.; McGaughran, A.; Uwe, J. Transcriptomic characterisation and genomic glimps into the toxigenic dinoflagellate Azadinium spinosum, with emphasis on polykeitde synthase genes. BMC Genomics 2015, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.; Daoust, P.; Benribague, S. A transcriptome-based perspective of cell cycle regulation in dinoflagellates. Protist 2016, 167, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Lin, S. Distinct gene number-genome size relationships for eukaryotes and non-eukaryotes: Gene content estimation for dinoflagellate genomes. PLoS ONE 2009, 4, e6978. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Lawrence, J.G.; Groisman, E.A. Lateral gene transfer and the nature of bacterial innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Herzog, M.; Von Boletzky, S.; Soyer, M.-O. Ultrastructural and biochemical nuclear aspects of eukaryote classification: Independent evolution of the dinoflagellates as a sister group of the actual eukaryotes? Orig. Life 1984, 13, 205–215. [Google Scholar] [CrossRef]
- Broyles, S.S.; Pettijohn, D.E. Interaction of the escherichia coli hu protein with DNA: Evidence for formation of nucleosome-like structures with altered DNA helical pitch. J. Mol. Biol. 1986, 187, 47–60. [Google Scholar] [CrossRef]
- Carone, B.R.; Hung, J.-H.; Hainer, S.J.; Chou, M.-T.; Carone, D.M.; Weng, Z.; Fazzio, T.G.; Rando, O.J. High-resolution mapping of chromatin packaging in mouse embryonic stem cells and sperm. Dev. Cell 2014, 30, 11–22. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riaz, S.; Sui, Z.; Niaz, Z.; Khan, S.; Liu, Y.; Liu, H. Distinctive Nuclear Features of Dinoflagellates with A Particular Focus on Histone and Histone-Replacement Proteins. Microorganisms 2018, 6, 128. https://doi.org/10.3390/microorganisms6040128
Riaz S, Sui Z, Niaz Z, Khan S, Liu Y, Liu H. Distinctive Nuclear Features of Dinoflagellates with A Particular Focus on Histone and Histone-Replacement Proteins. Microorganisms. 2018; 6(4):128. https://doi.org/10.3390/microorganisms6040128
Chicago/Turabian StyleRiaz, Sadaf, Zhenghong Sui, Zeeshan Niaz, Sohrab Khan, Yuan Liu, and Haoxin Liu. 2018. "Distinctive Nuclear Features of Dinoflagellates with A Particular Focus on Histone and Histone-Replacement Proteins" Microorganisms 6, no. 4: 128. https://doi.org/10.3390/microorganisms6040128