
Release of Bioactive Peptides from Erythrina edulis (Chachafruto) Proteins under Simulated Gastrointestinal Digestion



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Seed Flour Analysis
2.3. Protein Concentrate from Chachafruto Seeds (CPC)
2.4. Simulated Gastrointestinal Digestion
2.5. Antioxidant Activity by Biochemical Assays
2.6. Protective Effects in Macrophage RAW264.7
2.6.1. Cell Culture
2.6.2. Effects on Cell Viability
2.6.3. Effects on ROS Generation
2.6.4. Effects on NO Levels
2.7. Statistical Analysis
3. Results and Discussion
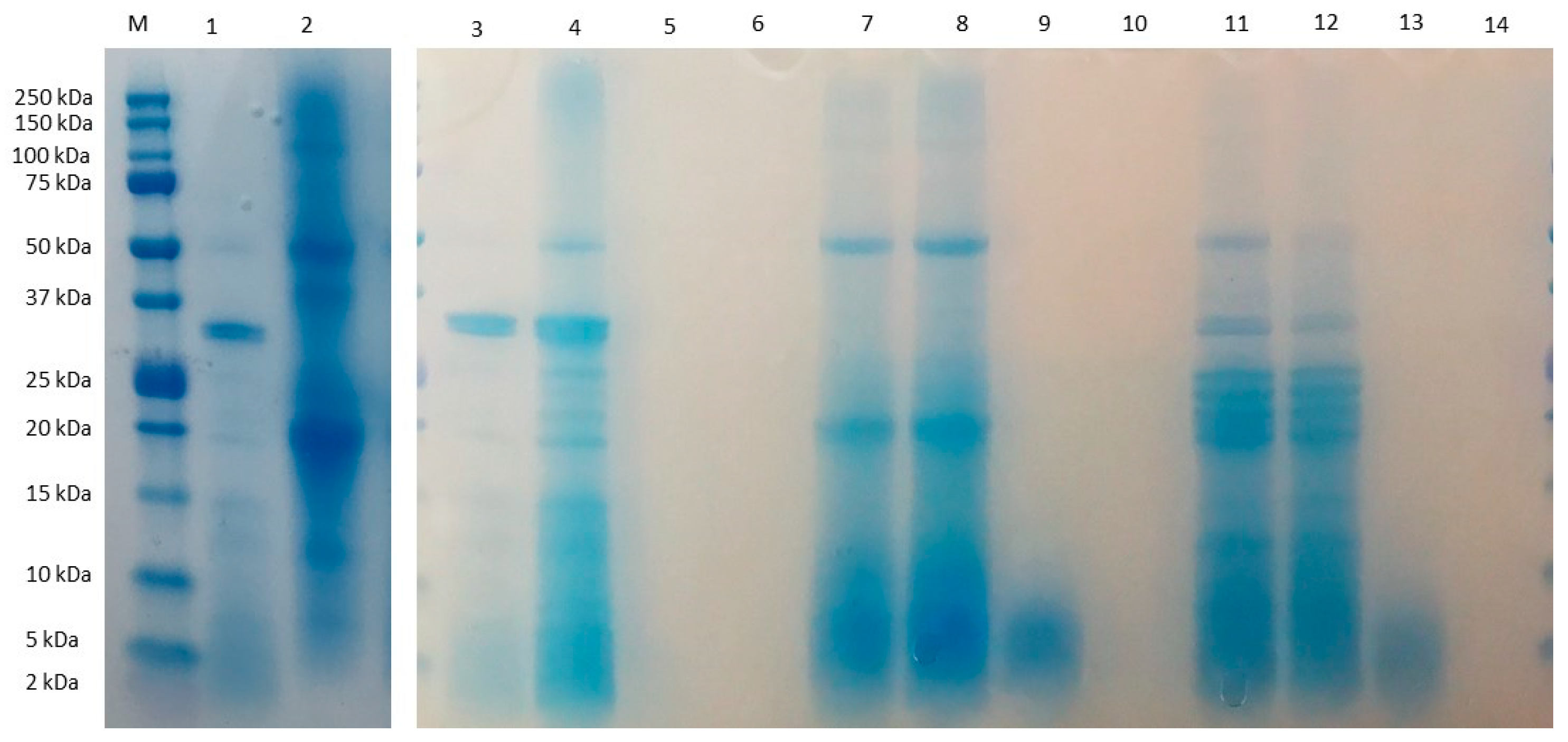
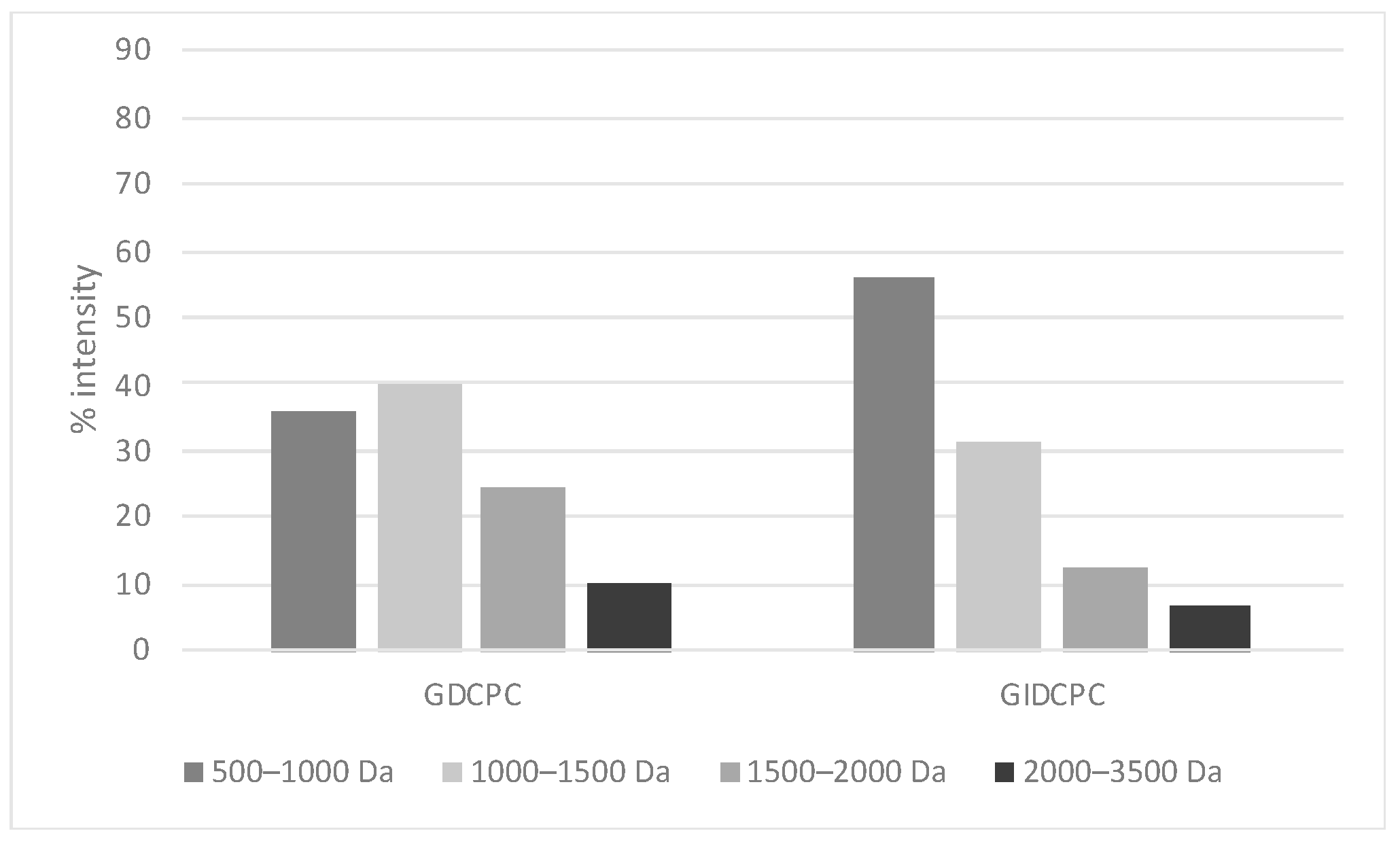
3.1. Characterization of Chachafruto Seed Protein Concentrate (CPC): Behavior under Simulated Gastrointestinal Digestion
3.2. Impact of the Gastrointestinal Digestion on the Antioxidant Activity of Chachafruto Seed Protein Concentrate (CPC)
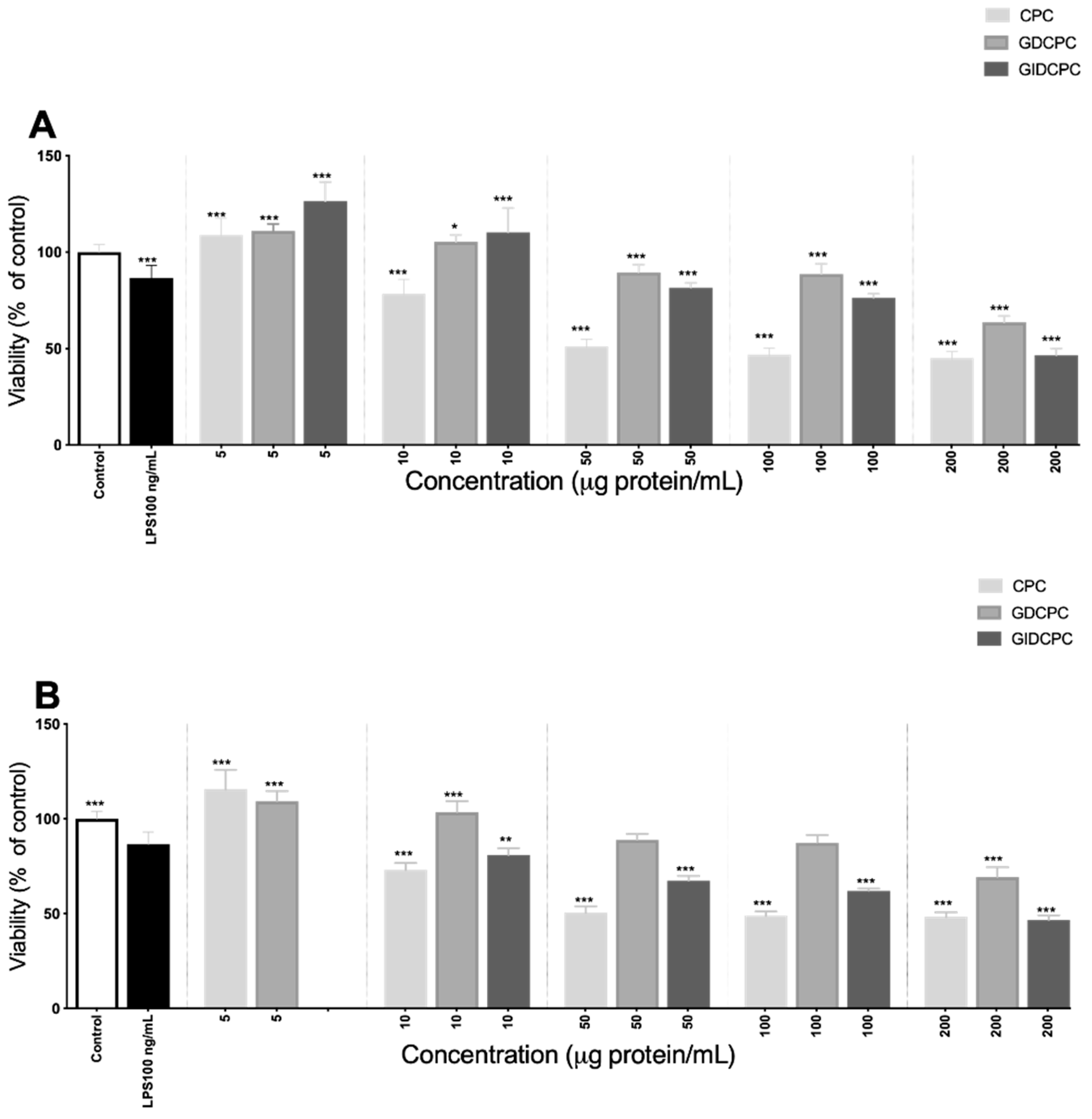
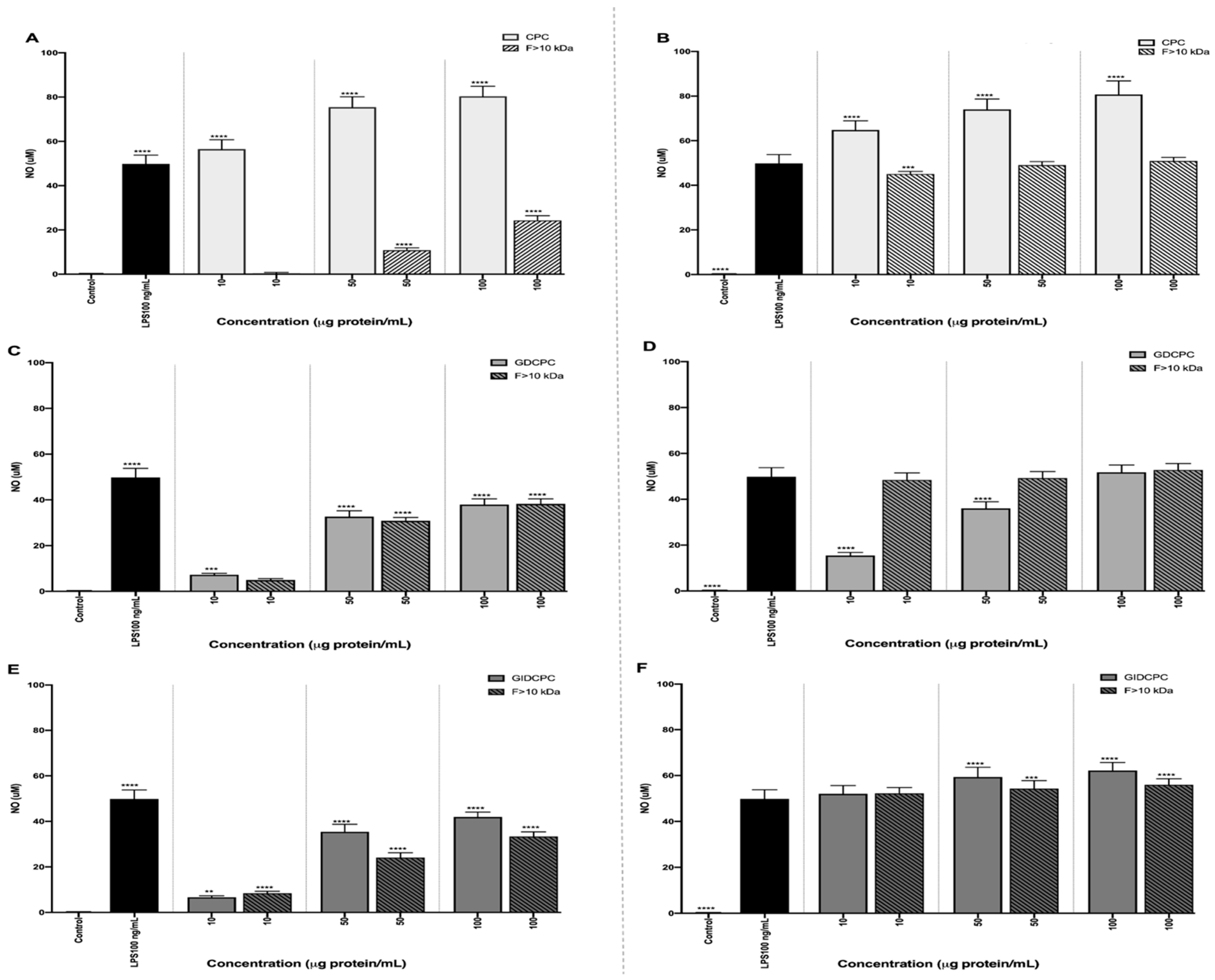
3.3. Effect of Chachafruto Seed Protein Concentrate (CPC) on Macrophage RAW264.7 Cells under Basal and Stimulated Conditions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vermeir, I.; Weijters, B.; De Houwer, J.; Geuens, M.; Slabbinck, H.; Spruyt, A.; Van Kerckhove, A.; Van Lippevelde, W.; De Steur, H.; Verbeke, W. Environmentally Sustainable Food Consumption: A Review and Research Agenda from a Goal-Directed Perspective. Front. Psychol. 2020, 11, 1603. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto, A.; Mattila, P.; Mäkinen, S.; Pajari, A.-M. Bioactivities of Alternative Protein Sources and Their Potential Health Benefits. Food Funct. 2017, 8, 3443–3458. [Google Scholar] [CrossRef] [PubMed]
- Otero, D.M.; Mendes, G.D.L.; Lucas, A.J.D.; Christ-Ribeiro, A.; Ribeiro, C.D.F. Exploring Alternative Protein Sources: Evidence from Patents and Articles Focusing on Food Markets. Food Chem. 2022, 394, 133486. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Chero, M.J.; Sánchez Chero, J.A.; Miranda Zamora, W. Technify and Conserve the Bioactive Components of Pashul (Erythrina edulis) for Human Consumption. UCV HACER Rev. Inv. Cult. 2019, 8, 11–17. [Google Scholar]
- Intiquilla, A.; Jiménez-Aliaga, K.; Zavaleta, A.I.; Arnao, I.; Peña, C.; Chávez-Hidalgo, E.L.; Hernández-Ledesma, B. Erythrina edulis (Pajuro) Seed Protein: A New Source of Antioxidant Peptides. Nat. Prod. Commun. 2016, 11, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intiquilla, A.; Jiménez-Aliaga, K.; Guzmán, F.; Alvarez, C.A.; Zavaleta, A.I.; Izaguirre, V.; Hernández-Ledesma, B. Novel Antioxidant Peptides Obtained by Alcalase Hydrolysis of Erythrina edulis (Pajuro) Protein. J. Sci. Food Agric. 2019, 99, 2420–2427. [Google Scholar] [CrossRef]
- Palma-Albino, C.; Intiquilla, A.; Jiménez-Aliaga, K.; Rodríguez-Arana, N.; Solano, E.; Flores, E.; Zavaleta, A.I.; Izaguirre, V.; Hernández-Ledesma, B. Albumin from Erythrina edulis (Pajuro) as a Promising Source of Multifunctional Peptides. Antioxidants 2021, 10, 1722. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Miralles, B.; Carrillo, W.; Hernández-Ledesma, B. In Vitro Chemopreventive Properties of Peptides Released from Quinoa (Chenopodium quinoa Willd.) Protein under Simulated Gastrointestinal Digestion. Food Res. Int. 2018, 105, 403–411. [Google Scholar] [CrossRef]
- Phongthai, S.; D’Amico, S.; Schoenlechner, R.; Homthawornchoo, W.; Rawdkuen, S. Fractionation and Antioxidant Properties of Rice Bran Protein Hydrolysates Stimulated by In Vitro Gastrointestinal Digestion. Food Chem. 2018, 240, 156–164. [Google Scholar] [CrossRef]
- Nascimento, E.S.D.; Anaya, K.; de Oliveira, J.M.C.; de Lacerda, J.T.J.G.; Miller, M.E.; Dias, M.; Mendes, M.A.; Pallone, J.d.A.L.; Arns, C.W.; Juliano, M.A.; et al. Identification of Bioactive Peptides Released from In Vitro Gastrointestinal Digestion of Yam Proteins (Dioscorea cayennensis). Food Res. Int. 2021, 143, 110–286. [Google Scholar]
- Cipollone, M.A.; Tironi, V.A. Yellow Pea Flour and Protein Isolate as Sources of Antioxidant Peptides after Simulated Gastrointestinal Digestion. Legume Sci. 2020, 2, e59. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Bellesia, A.; Conte, A. Identification of ACE-Inhibitory Peptides from Phaseolus vulgaris after In Vitro Gastrointestinal Digestion. Int. J. Food Sci. Nutr. 2015, 66, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Luna-Vital, D.; de Mejía, E.G. Peptides from Legumes with Antigastrointestinal Cancer Potential: Current Evidence for Their Molecular Mechanisms. Curr. Opin. Food Sci. 2018, 20, 13–18. [Google Scholar] [CrossRef]
- Wang, S.; Rao, P.; Ye, X. Isolation and Biochemical Characterization of a Novel Leguminous Defense Peptide with Antifungal and Antiproliferative Potency. Appl. Microbiol. Biotechnol. 2009, 82, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Pavlicevic, M.; Marmiroli, N.; Maestri, E. Immunomodulatory Peptides—A Promising Source for Novel Functional Food Production and Drug Discovery. Peptides 2022, 148, 170696. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and Anticancer Protein Hydrolysates (Peptides) from Food Proteins: A Review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef]
- Kiewiet, M.B.; Faas, M.M.; De Vos, P. Immunomodulatory Protein Hydrolysates and Their Application. Nutrients 2018, 10, 904. [Google Scholar] [CrossRef] [Green Version]
- Cunniff, P.; Association of Official Analytical Chemists (Eds.) Official Methods of Analysis of AOAC International; AOAC: Washington, DC, USA, 1999; ISBN 978-0-935584-54-7. [Google Scholar]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D. A Standardised Static In Vitro Digestion Method Suitable for Food–An International Consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Ojalvo, D.; Molina, E.; López-Fandiño, R. Hydrolysates of Egg White Proteins Modulate T- and B-Cell Responses in Mitogen-Stimulated Murine Cells. Food Funct. 2016, 7, 1048–1056. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Dávalos, A.; Bartolomé, B.; Amigo, L. Preparation of Antioxidant Enzymatic Hydrolysates from α-Lactalbumin and β-Lactoglobulin. Identification of Active Peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the Probe 2′,7′-Dichlorofluorescin as an Indicator of Reactive Oxygen Species Formation and Oxidative Stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Villaluenga, C.; Dia, V.P.; Berhow, M.; Bringe, N.A.; Gonzalez de Mejia, E. Protein Hydrolysates from β-Conglycinin Enriched Soybean Genotypes Inhibit Lipid Accumulation and Inflammation In Vitro. Mol. Nutr. Food Res. 2009, 53, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Arango Bedoya, O.; Bolaños Patiño, V.; Ricaurte García, D.; Caicedo, M.; Guerrero, Y. Obtención de un extracto proteico a partir de harina de chachafruto (Erythrina edulis). Univ. Salud 2012, 14, 161–167. [Google Scholar]
- Klupšaitė, D.; Juodeikienė, G. Legume: Composition, Protein Extraction and Functional Properties. A Review. Chem. Technol. 2015, 66, 5–12. [Google Scholar] [CrossRef]
- Villafuerte, F.; Pérez, E.; Mahfoud, A.; Valero, Y.; Enríquez, M.; Yanez, K.; Manobanda, P. Characerization of Erythrina Edulis Triana and Obtaining Protein Isolate. In Proceedings of the CICABI, Ambato, Ecuador, 25–29 June 2018; p. 233. [Google Scholar]
- Teka, T.A.; Retta, N.; Bultosa, G.; Admassu, H.; Astatkie, T. Protein Fractions, In Vitro Protein Digestibility and Amino Acid Composition of Select Cowpea Varieties Grown in Ethiopia. Food Biosci. 2020, 36, 100634. [Google Scholar] [CrossRef]
- Pastor-Cavada, E.; Juan, R.; Pastor, J.E.; Alaiz, M.; Vioque, J. Protein and Amino Acid Composition of Select Wild Legume Species of Tribe Fabeae. Food Chem. 2014, 163, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.; Yao, X.; Li, A. Antioxidant Properties of Peptide from Cowpea Seed. Int. J. Food Prop. 2013, 16, 1245–1256. [Google Scholar] [CrossRef]
- Heldt, H.-W.; Piechulla, B. Chapter 12—Products of Nitrogen Fixation and Nitrate Assimilation Are Deposited as Storage Proteins. In Plant Biochemistry, 5th ed.; Heldt, H.-W., Piechulla, B., Eds.; Academic Press: Rostock, Germany, 2021; pp. 299–305. ISBN 978-0-12-818631-2. [Google Scholar]
- Junejo, S.A.; Ding, L.; Fu, X.; Xiong, W.; Zhang, B.; Huang, Q. Pea Cell Wall Integrity Controls the Starch and Protein Digestion Properties in the Infogest In Vitro Simulation. Int. J. Biol. Macromol. 2021, 182, 1200–1207. [Google Scholar] [CrossRef]
- Gómez, A.; Gay, C.; Tironi, V.; Avanza, M.V. Structural and Antioxidant Properties of Cowpea Protein Hydrolysates. Food Biosci. 2021, 41, 101074. [Google Scholar] [CrossRef]
- Zhan, Q.; Wang, Q.; Liu, Q.; Guo, Y.; Gong, F.; Hao, L.; Wu, H.; Dong, Z. The Antioxidant Activity of Protein Fractions from Sacha Inchi Seeds after a Simulated Gastrointestinal Digestion. LWT 2021, 145, 111356. [Google Scholar] [CrossRef]
- Jodayree, S.; Smith, J.C.; Tsopmo, A. Use of Carbohydrase to Enhance Protein Extraction Efficiency and Antioxidative Properties of Oat Bran Protein Hydrolysates. Food Res. Int. 2012, 46, 69–75. [Google Scholar] [CrossRef]
- You, L.J.; Zhao, M.M.; Regenstein, J.M.; Ren, J.Y. Purification and Identification of Antioxidative Peptides from Loach (Misgurnus anguillicaudatus) Protein Hydrolysate by Consecutive Chromatography and Electrospray Ionization-Mass Spectrometry. Food Res. Int. 2010, 43, 1167–1173. [Google Scholar] [CrossRef]
- Feng, L.; Peng, F.; Wang, X.; Li, M.; Lei, H.; Xu, H. Identification and Characterization of Antioxidative Peptides Derived from Simulated In Vitro Gastrointestinal Digestion of Walnut Meal Proteins. Food Res. Int. 2019, 116, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Peighambardoust, S.H.; Karami, Z.; Pateiro, M.; Lorenzo, J.M. A Review on Health-Promoting, Biological, and Functional Aspects of Bioactive Peptides in Food Applications. Biomolecules 2021, 11, 631. [Google Scholar] [CrossRef]
- Sannasimuthu, A.; Kumaresan, V.; Anilkumar, S.; Pasupuleti, M.; Ganesh, M.-R.; Mala, K.; Paray, B.A.; Al-Sadoon, M.K.; Albeshr, M.F.; Arockiaraj, J. Design and Characterization of a Novel Arthrospira Platensis Glutathione Oxido-Reductase-Derived Antioxidant Peptide GM15 and Its Potent Anti-Cancer Activity via Caspase-9 Mediated Apoptosis in Oral Cancer Cells. Free Radic. Biol. Med. 2019, 135, 198–209. [Google Scholar] [CrossRef]
- Ngo, N.T.T.; Shahidi, F. Antioxidant Properties of Camelina (Camelina sativa (L.) Crantz) Protein Hydrolysates. J. Food Bioact. 2021, 16, 75–82. [Google Scholar] [CrossRef]
- Afrin, S.; Gasparrini, M.; Forbes-Hernández, T.Y.; Cianciosi, D.; Reboredo-Rodriguez, P.; Manna, P.P.; Battino, M.; Giampieri, F. Protective Effects of Manuka Honey on LPS-Treated RAW 264.7 Macrophages. Part 1: Enhancement of Cellular Viability, Regulation of Cellular Apoptosis and Improvement of Mitochondrial Functionality. Food Chem. Toxicol. 2018, 121, 203–213. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Han, S.H.; Lee, Y.W.; Lee, M.; Yang, K.H.; Kim, H.M. Dexamethasone Inhibits IL-1β Gene Expression in LPS-Stimulated RAW 264.7 Cells by Blocking NF-ΚB/Rel and AP-1 Activation. Immunopharmacology 2000, 48, 173–183. [Google Scholar] [CrossRef]
- Kim, Y.; Moon, J.S.; Lee, K.S.; Park, S.Y.; Cheong, J.; Kang, H.S.; Lee, H.Y.; Do Kim, H. Ca2+/Calmodulin-Dependent Protein Phosphatase Calcineurin Mediates the Expression of INOS through IKK and NF-ΚB Activity in LPS-Stimulated Mouse Peritoneal Macrophages and RAW 264.7 Cells. Biochem. Biophys. Res. Commun. 2004, 314, 695–703. [Google Scholar] [CrossRef]
- Song, S.; Li, T.; Cheng, X.; Ji, X.; Gao, D.; Du, M.; Jiang, N.; Liud, X.; Mao, X. Sea Cucumber Peptides Exert Anti-Inflammatory Activity Through Suppressing NF-κB and MAPK and Inducing HO-1 in RAW264.7 Macrophages. Food Funct. 2016, 7, 2773. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Dong, L.; Du, H.; Bao, Z.; Lin, S. Potential Mechanisms Underlying the Protective Effects of Tricholoma matsutake Singer Peptides Against LPS-Induced Inflammation in RAW264.7 Macrophages. Food Chem. 2021, 353, 129452. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.D.; Ha, B.J. Paeoniflorin Protects RAW 264.7 Macrophages from LPS-Induced Cytotoxicity and Genotoxicity. Toxicol. In Vitro 2009, 23, 1014–1019. [Google Scholar] [CrossRef]
- Chao, W.-W.; Chung, Y.-C.; Shih, I.-P.; Wang, H.-Y.; Chou, S.-T.; Hsu, C.-K. Red Bean Extract Inhibits Lipopolysaccharide-Induced Inflammation and H2O2-Induced Oxidative Stress in RAW 264.7 Macrophages. J. Med. Food 2015, 18, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, X.; Ren, G.; Wu, C.; Qin, P.; Yao, Y. Peptides from Extruded Lupin (Lupinus albus L.) Regulate Inflammatory Activity via the P38 MAPK Signal Transduction Pathway in RAW 264.7 Cells. J. Agric. Food Chem. 2020, 68, 11702–11709. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Zeng, Y.; Tian, H.; Zhang, Z.; Zhang, H.; Huang, F.; Yu, F. Macrophage Immunomodulatory Effects of Low Molecular Weight Peptides from Mytilus coruscus via NF-κB/MAPK Signaling Pathways. J. Funct. Foods 2021, 83, 104562. [Google Scholar] [CrossRef]
- Frassinetti, S.; Gabriele, M.; Caltavuturo, L.; Longo, V.; Pucci, L. Antimutagenic and Antioxidant Activity of a Selected Lectin-Free Common Bean (Phaseolus vulgaris L.) in Two Cell-Based Models. Plant Foods Hum. Nutr. 2015, 70, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Nishanth, R.P.; Jyotsna, R.G.; Schlager, J.J.; Hussain, S.M.; Reddanna, P. Inflammatory Responses of RAW 264.7 Macrophages upon Exposure to Nanoparticles: Role of ROS-NFκB Signaling Pathway. Nanotoxicology 2011, 5, 502–516. [Google Scholar] [CrossRef]
- Martínez-Leo, E.E.; Martín-Ortega, A.M.; Acevedo-Fernández, J.J.; Moo-Puc, R.; Segura-Campos, M.R. Peptides from Mucuna pruriens L., with Protection and Antioxidant In Vitro Effect on HeLa Cell Line. J. Sci. Food Agric. 2019, 99, 4167–4173. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Antioxidant and Anti-Inflammatory Properties of Cancer Preventive Peptide Lunasin in RAW 264.7 Macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 803–808. [Google Scholar] [CrossRef]
- Piñuel, M.L.; Vilcacundo, E.; Boeri, P.A.; Barrio, D.A.; Morales, D.; Pinto, A.; Moran, R.; Samaniego, I.; Carrillo, W. Extraction of Protein Concentrate from Red Bean (Phaseolus vulgaris L.): Antioxidant Activity and Inhibition of Lipid Peroxidation. J. Appl. Pharm. Sci. 2019, 9, 45–58. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Amino Acid Content (g/100 g Protein) | Amino Acid Content (g/100 g Product) | FAO Recommendations |
|---|---|---|---|
| Essential | |||
| Lysine | 4.00 ± 0.13 | 3.30 ± 0.11 | 5.20 |
| Tryptophan | n.d. | n.d. | 0.70 |
| Phenylalanine | 3.38 ± 0.10 | 2.78 ± 0.08 | 4.60 a |
| Tyrosine | 3.76 ± 0.08 | 3.10 ± 0.06 | |
| Methionine | 1.01 ± 0.08 | 0.83 ± 0.06 | 2.60 b |
| Cysteine | 0.90 ± 0.14 | 0.74 ± 0.12 | |
| Threonine | 2.56 ± 0.06 | 2.11 ± 0.05 | 2.70 |
| Leucine | 6.02 ± 0.17 | 4.97 ± 0.14 | 6.30 |
| Isoleucine | 2.32 ± 0.08 | 1.91 ± 0.06 | 3.10 |
| Valine | 3.23 ± 0.02 | 2.67 ± 0.01 | 4.20 |
| Non-essential | |||
| Aspartic acid + asparagine | 8.49 ± 0.31 | 7.00 ± 0.26 | |
| Glutamic acid + glutamine | 12.34 ± 0.36 | 10.18 ± 0.30 | |
| Serine | 4.69 ± 0.11 | 3.87 ± 0.09 | |
| Histidine | 1.68 ± 0.10 | 1.38 ± 0.08 | |
| Arginine | 3.52 ± 0.18 | 2.90 ± 0.15 | |
| Alanine | 2.80 ± 0.13 | 2.31 ± 0.11 | |
| Proline | 3.83 ± 0.09 | 3.16 ± 0.08 | |
| Glycine | 3.28 ± 0.15 | 2.70 ± 0.12 | |
| TAA | 67.81 | 55.91 | |
| HAA | 27.25 | 22.48 | |
| AAA | 7.13 | 5.89 |
| Sample | TEAC (µmol TE/mg Protein) | ORAC (µmol TE/mg Protein) | ||||||
|---|---|---|---|---|---|---|---|---|
| Whole Sample | F ˃ 10 kDa | F 3–10 kDa | F ˂ 3 kDa | Whole Sample | F ˃ 10 kDa | F 3–10 kDa | F ˂ 3 kDa | |
| CPC | 0.10 ± 0.01 | 0.20 ± 0.00 | 0.47 ± 0.01 | 0.69 ± 0.07 | 0.47 ± 0.02 | 0.70 ± 0.01 | 0.79 ±0.01 | 4.29 ± 0.13 |
| GDCPC | 0.33 ± 0.02 | 0.17 ± 0.01 | 0.34 ± 0.00 | 0.66 ± 0.06 | 1.13 ± 0.02 | 0.47 ± 0.02 | 1.10 ± 0.02 | 1.93 ± 0.13 |
| GIDCPC | 0.46 ± 0.01 | 0.21 ± 0.01 | 0.30 ± 0.02 | 0.79 ± 0.00 | 1.12 ± 0.07 | 0.68 ± 0.02 | 0.96 ± 0.02 | 2.20 ± 0.18 |
| Sample | Concentration (µg/mL) | ROS Generation (% of Control) | |||
|---|---|---|---|---|---|
| Whole Sample | Fraction ˃ 10 kDa | Fraction 3–10 kDa | Fraction ˂ 3 kDa | ||
| Control | - | 100.00 ± 4.87 | |||
| LPS | 0.1 | 186.12 ± 16.43 *** | |||
| CPC | 5.0 | 65.55 ± 3.69 *** | 85.73 ± 6.26 * | 57.03 ± 4.40 *** | 52.84 ± 4.43 *** |
| 10.0 | 102.56 ± 10.56 | 116.25 ± 6.47 * | 58.14 ± 4.44 *** | 50.54 ± 3.28 *** | |
| 50.0 | 140.62 ± 8.44 *** | 188.80 ± 8.90 *** | 60.21 ± 3.85 *** | 57.26 ± 5.61 *** | |
| 100.0 | 163.20 ± 15.91 *** | 225.02 ± 6.63 *** | 66.48 ± 5.97 *** | 61.19 ± 3.34 *** | |
| GDCPC | 5.0 | 41.96 ± 5.04 *** | 70.56 ± 6.00 *** | 61.70 ± 2.30 *** | 58.80 ± 2.69 *** |
| 10.0 | 50.76 ± 6.12 *** | 75.00 ± 5.17 *** | 57.97 ± 2.28 *** | 60.67 ± 2.97 *** | |
| 50.0 | 115.53 ± 10.16 * | 199.00 ± 19.47 *** | 82.02 ± 5.62 *** | 70.23 ± 3.21 *** | |
| 100.0 | 137.26 ± 8.69 *** | 236.38 ± 22.35 *** | 118.59 ± 5.92 ** | 75.19 ± 4.69 *** | |
| GIDCPC | 5.0 | 67.56 ± 3.77 *** | 66.30 ± 4.86 *** | 53.76 ± 4.38 *** | 61.02 ± 2.51 *** |
| 10.0 | 74.00 ± 6.83 ** | 106.74 ± 10.26 | 55.42 ± 5.23 *** | 66.99 ± 2.13 *** | |
| 50.0 | 224.44 ± 19.08 *** | 180.46 ± 14.09 *** | 68.96 ± 3.13 *** | 73.91 ± 4.23 *** | |
| 100.0 | 306.17 ± 26.57 *** | 245.40 ± 8.98 *** | 70.23 ± 5.56 *** | 77.81 ± 2.30 *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correa, J.L.; Zapata, J.E.; Hernández-Ledesma, B. Release of Bioactive Peptides from Erythrina edulis (Chachafruto) Proteins under Simulated Gastrointestinal Digestion. Nutrients 2022, 14, 5256. https://doi.org/10.3390/nu14245256
Correa JL, Zapata JE, Hernández-Ledesma B. Release of Bioactive Peptides from Erythrina edulis (Chachafruto) Proteins under Simulated Gastrointestinal Digestion. Nutrients. 2022; 14(24):5256. https://doi.org/10.3390/nu14245256
Chicago/Turabian StyleCorrea, Jessica L, José Edgar Zapata, and Blanca Hernández-Ledesma. 2022. "Release of Bioactive Peptides from Erythrina edulis (Chachafruto) Proteins under Simulated Gastrointestinal Digestion" Nutrients 14, no. 24: 5256. https://doi.org/10.3390/nu14245256