Endemics and Cosmopolitans: Application of Statistical Mechanics to the Dry Forests of Mexico

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The Underlying Model
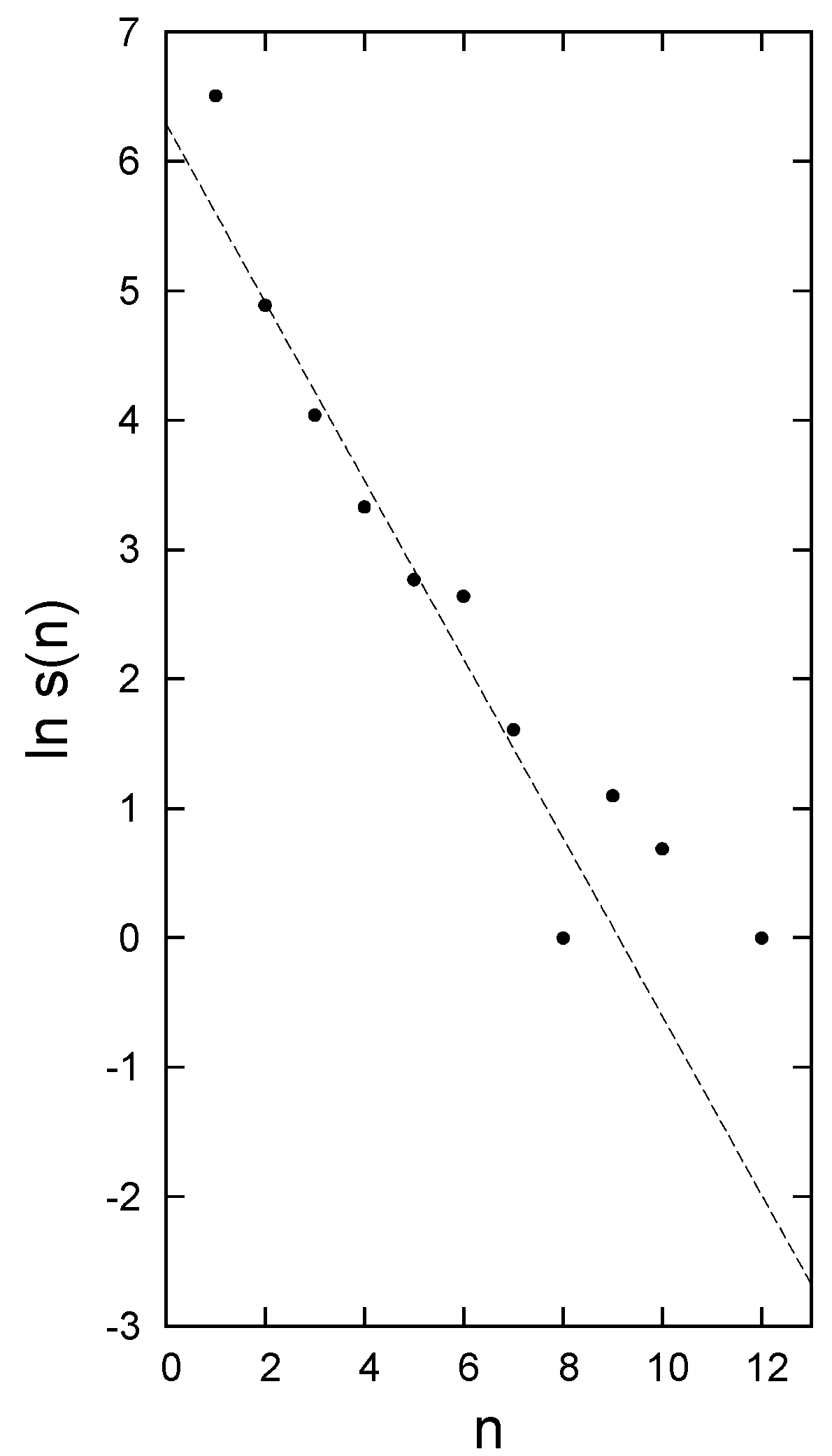
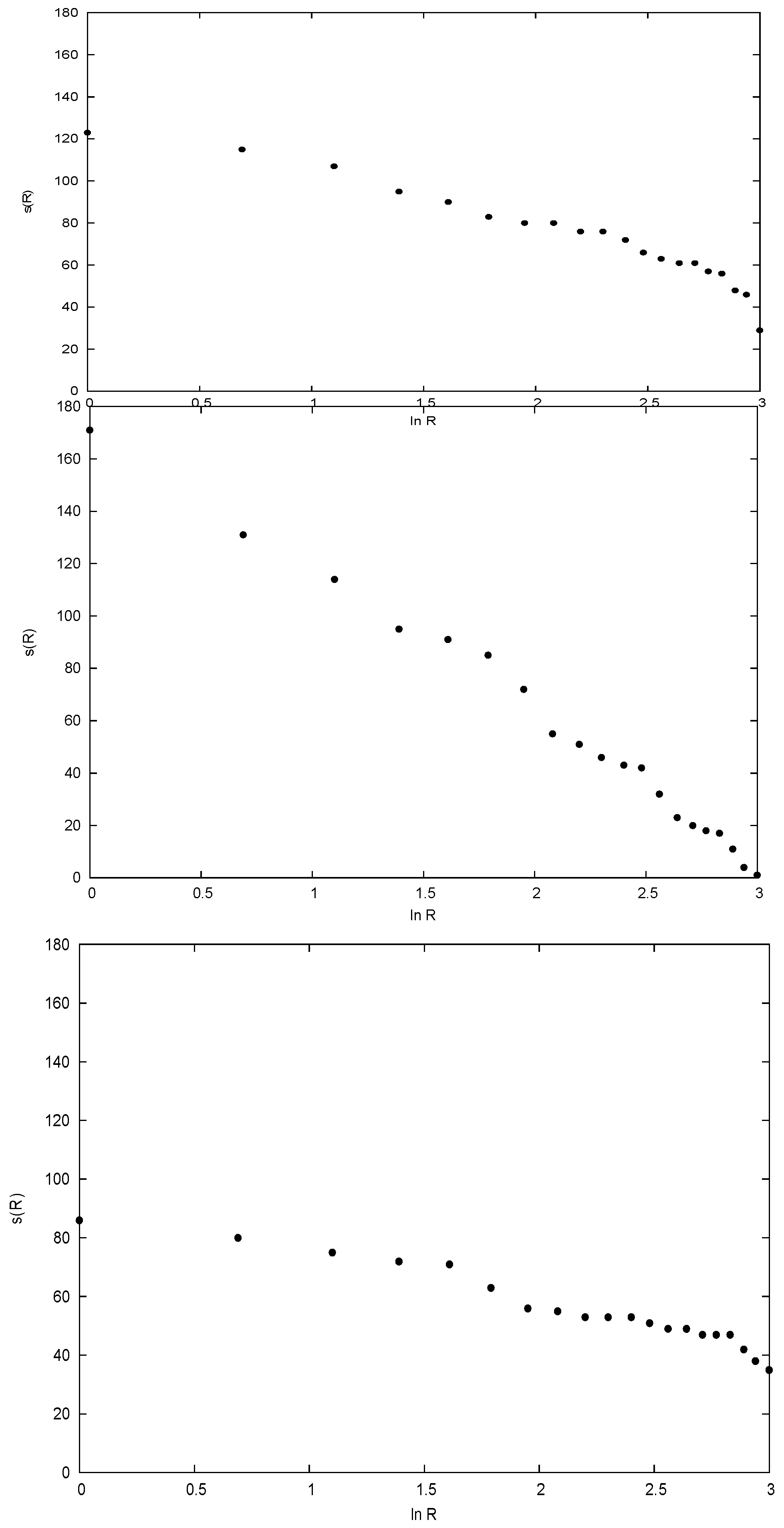
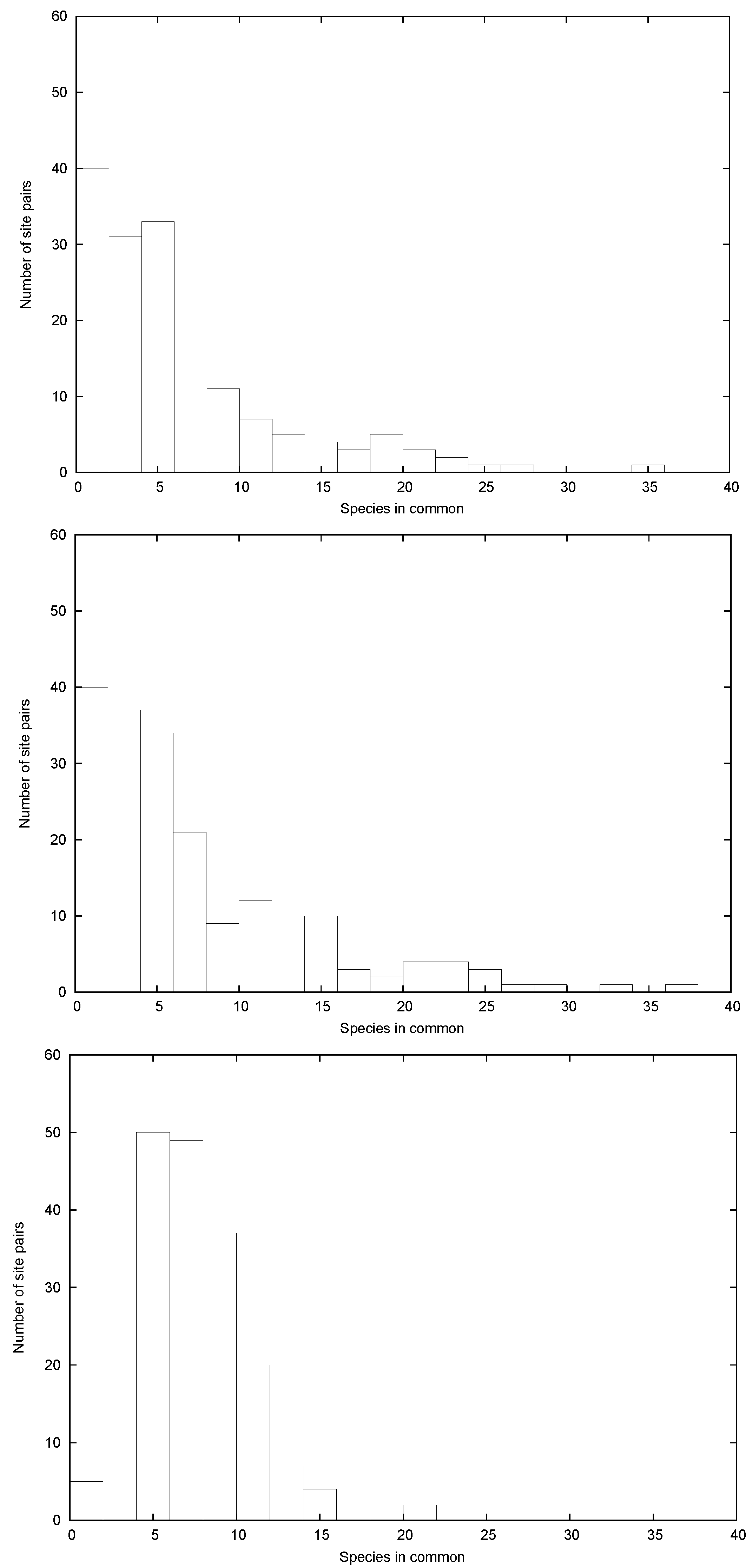
3. Relevant Aspects of the Data
4. Disentangling the Conundrum
5. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Pueyo, S.; He, F.; Zillio, T. The maximum entropy formalism and the idiosyncratic theory of biodiversity. Ecol. Lett. 2007, 10, 1017–1028. [Google Scholar] [CrossRef]
- Harte, J. Maximum Entropy and Ecology; Oxford Series in Ecology and Evolution; Oxford University Press: New York, NY, USA, 2011; ISBN 978-0-19-959341-5. [Google Scholar]
- Bowler, M.G.; Kelly, C.K. On the statistical mechanics of species abundance distributions. Theor. Popul. Biol. 2012, 82, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Bowler, M.G. Species abundance distributions, statistical mechanics and the priors of maxent. Theor. Popul. Biol. 2014, 92, 69–77. [Google Scholar] [CrossRef]
- Kelly, C.K.; Blundell, S.J.; Bowler, M.G.; Fox, G.A.; Harvey, P.H.; Lomas, M.R.; Ian Woodward, F. The statistical mechanics of community assembly and species distribution. N. Phytol. 2011, 191, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Bowler, M.G.; Kelly, C.K. On the statistical mechanics of alien species distribution. Entropy 2017, 19, 674. [Google Scholar] [CrossRef]
- Trejo, I.; Dirzo, R. Floristic diversity of Mexican seasonally dry tropical forests. Biodivers. Conserv. 2002, 11, 2063–2084. [Google Scholar] [CrossRef]
- MacArthur, R.H. On the relative abundance of species. Am. Nat. 1960, 94, 25–36. [Google Scholar] [CrossRef]
- Magurran, A.E.; Henderson, P.A. Explaining the excess of rare species in abundance distributions. Nature 2003, 422, 714–716. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowler, M.G.; Kelly, C.K. Endemics and Cosmopolitans: Application of Statistical Mechanics to the Dry Forests of Mexico. Entropy 2019, 21, 616. https://doi.org/10.3390/e21060616
Bowler MG, Kelly CK. Endemics and Cosmopolitans: Application of Statistical Mechanics to the Dry Forests of Mexico. Entropy. 2019; 21(6):616. https://doi.org/10.3390/e21060616
Chicago/Turabian StyleBowler, Michael G., and Colleen K. Kelly. 2019. "Endemics and Cosmopolitans: Application of Statistical Mechanics to the Dry Forests of Mexico" Entropy 21, no. 6: 616. https://doi.org/10.3390/e21060616
APA StyleBowler, M. G., & Kelly, C. K. (2019). Endemics and Cosmopolitans: Application of Statistical Mechanics to the Dry Forests of Mexico. Entropy, 21(6), 616. https://doi.org/10.3390/e21060616