A New Synthetic Compound, 2-OH, Enhances Interleukin-2 and Interferon-γ Gene Expression in Human Peripheral Blood Mononuclear Cells

Abstract
:1. Introduction
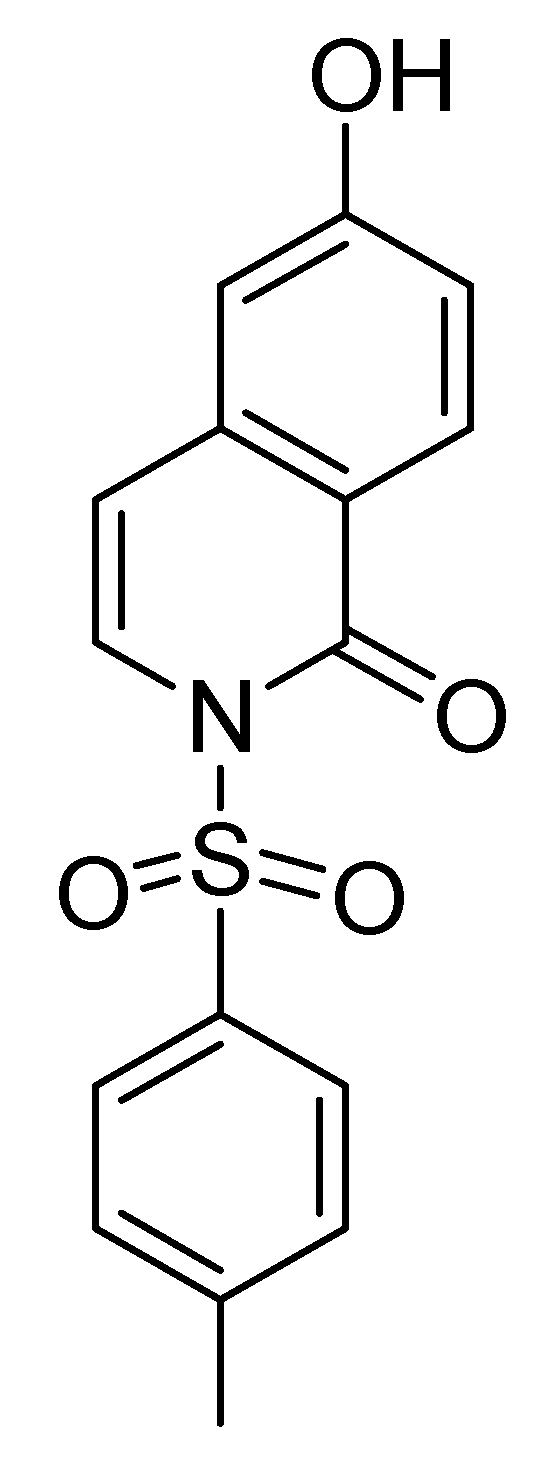
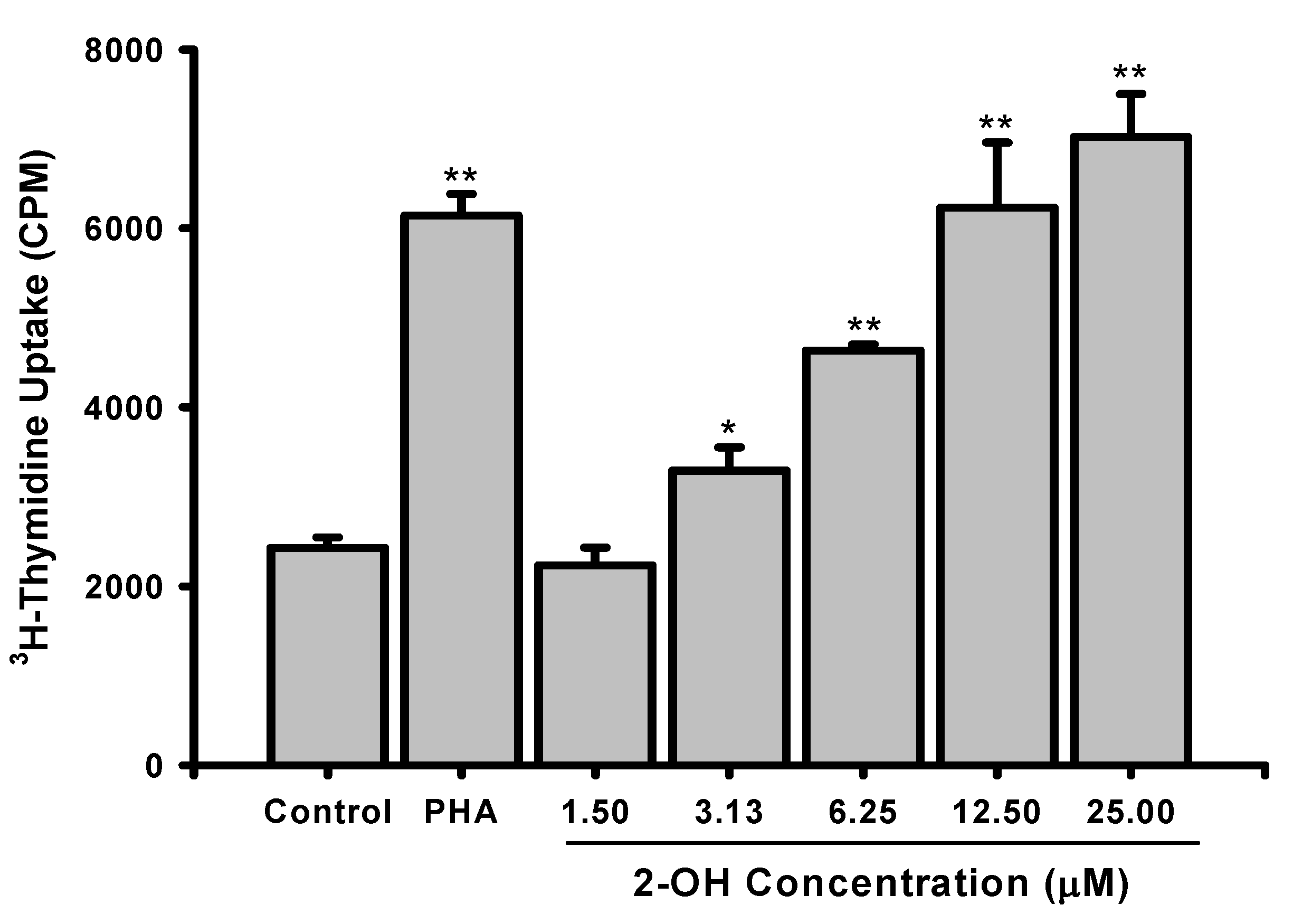
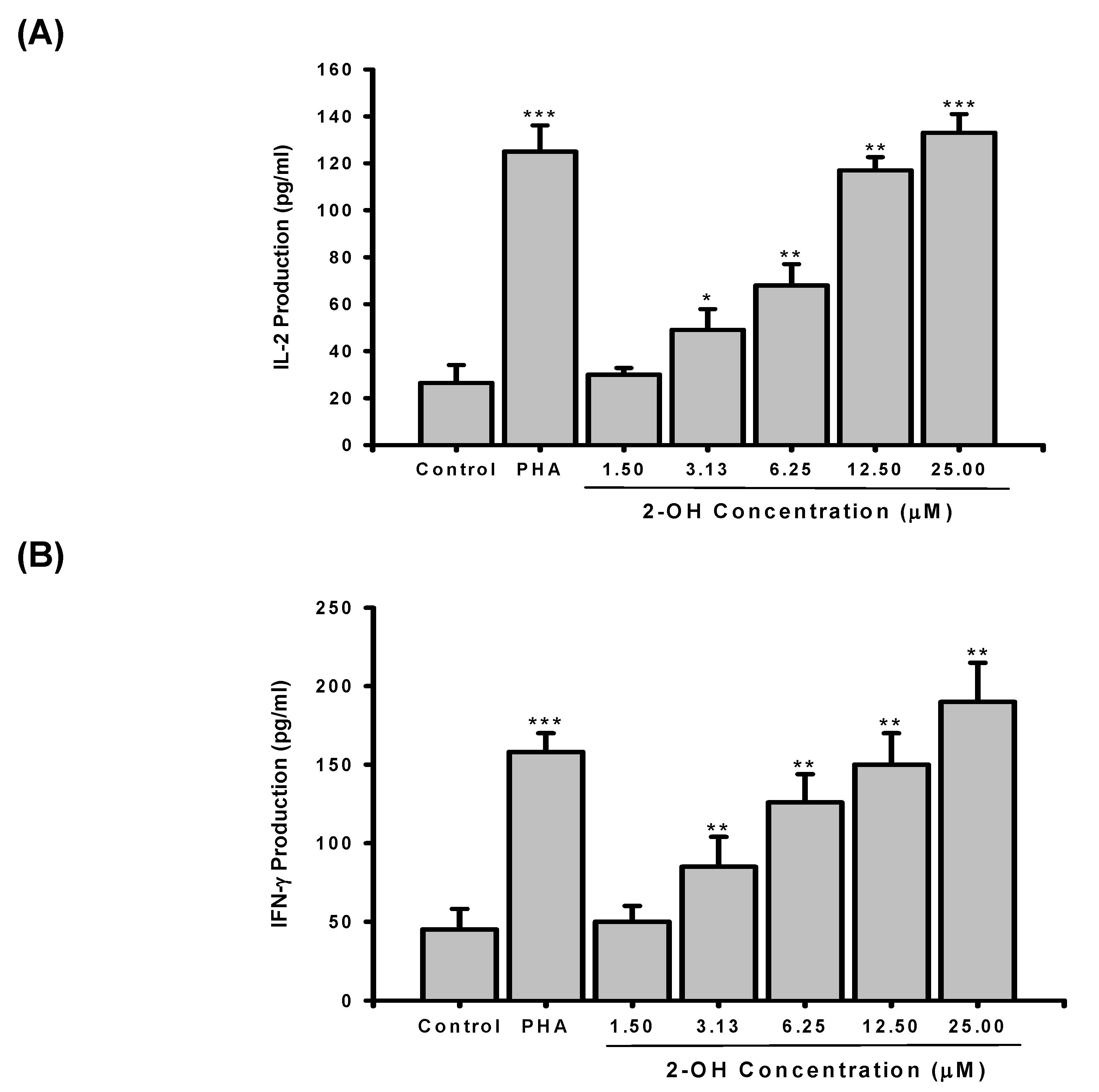
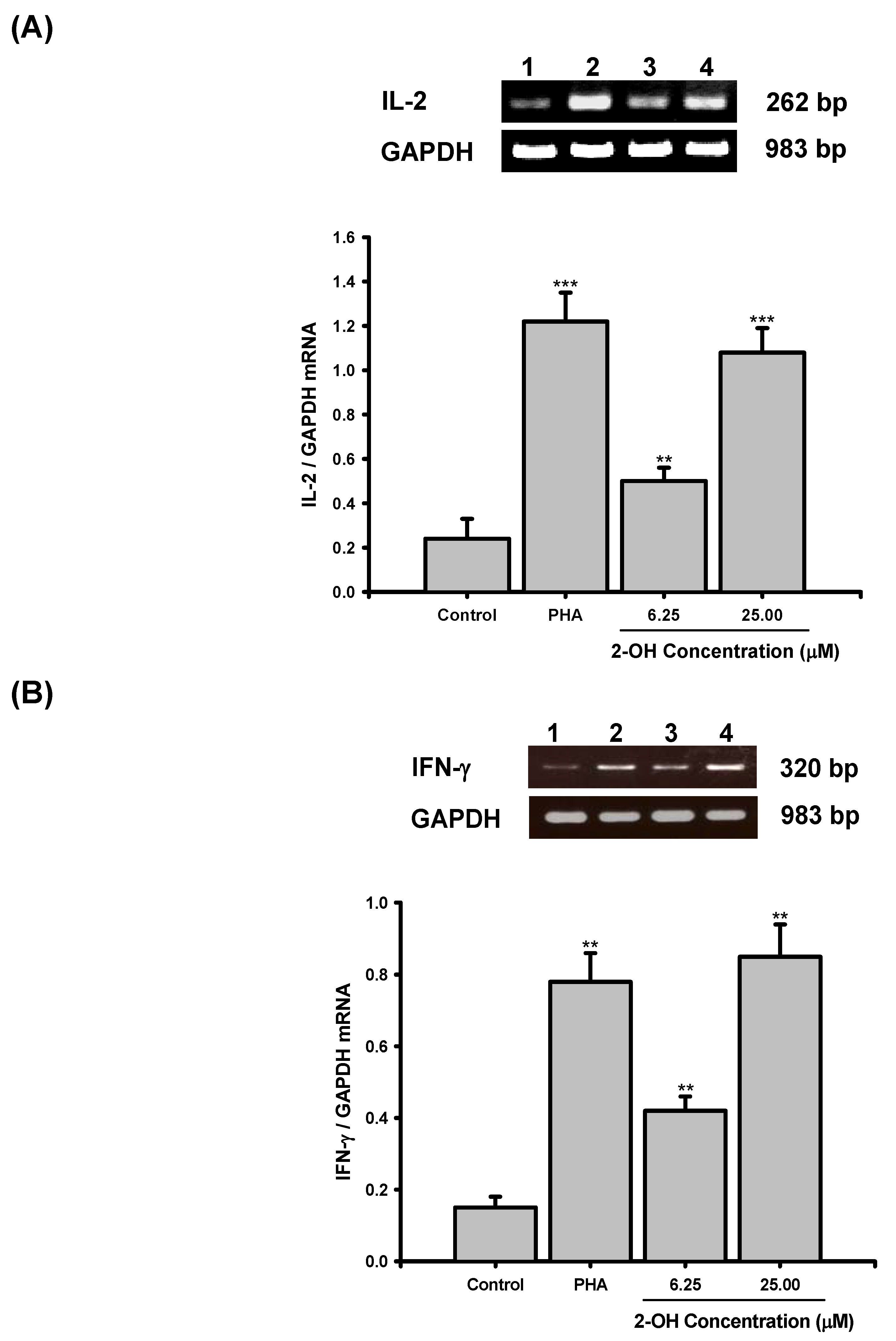
2. Results and Discussion




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | Sequence | Predicted size (bp) |
|---|---|---|
| IL-2 | 5’-GTC ACA AAC AGT GCA CCT AC-3’ 5’-GAA AGT GAA TTC TGG GTC CC-3’ | 262 |
| IFN-γ | 5’-GCA GAG CCA AAT TGT CTC CT-3’ 5’-ATG CTC TTC GAC CTC GAA AC-3’ | 320 |
| GAPDH | 5’- TGA AGG TCG GAG TCA ACG GAT TTG GT-3’ 5’- CAT GTG GGC CAT GAG GTC CAC CAC-3’ | 983 |

| CT | IL-2 ΔCT | IFN-γ ΔCT | |||
|---|---|---|---|---|---|
| IL-2 | IFN-γ | GAPDH | |||
| Control | 34.34±0.69 | 35.05±0.15 | 25.38±0.79 | 8.96±0.10 | 9.62±0.87 |
| PHA (5 μg/mL) | 27.97±0.34 | 28.25±0.19 | 24.57±0.04 | 3.40±0.38** | 3.76±0.11* |
| 2-OH (6.25 μM) | 30.56±0.45 | 31.27±0.04 | 24.87±0.11 | 5.69±0.56* | 6.40±0.16* |
| 2-OH (25 μM) | 34.34±0.69 | 28.60±0.64 | 25.30±0.46 | 4.49±0.12*** | 3.33±0.20** |
3. Experimental
3.1. Preparation of 2-OH
3.2. Preparation of PBMC
3.3. Lymphoproliferation test

3.4. Determination of IL-2 and IFN-γ production
3.5. Extraction of total cellular RNA
3.6. RT-PCR
3.7. Real-time PCR
3.8. Statistical analysis
4. Conclusions
Acknowledgements
References
- Numata, A.; Ibuka, I. Alkaloids from ants and other insects. In The Alkaloids; Brossi, A., Ed.; Academic Press: New York, NY, USA, 1987; p. 193. [Google Scholar]
- Liu, C.P.; Tsai, W.J.; Lin, Y.L.; Liao, J.F.; Chen, C.F.; Kuo, Y.C. The extracts from Nelumbo nucifera suppress cell cycle progression, cytokine genes expression, and cell proliferation in human peripheral blood mononuclear cells. Life Sci. 2004, 75, 699–716. [Google Scholar] [CrossRef]
- Liu, C.P.; Kuo, Y.C.; Lin, Y.L.; Liao, J.F.; Shen, C.C.; Chen, C.F.; Tsai, W.J. (S)-Armepavine inhibits human peripheral blood mononuclear cells activation by regulating Itk and PLCγ activation in a PI3K-dependent manner. J. Leukocyte Biol. 2007, 81, 1276–1286. [Google Scholar] [CrossRef]
- Liu, C.P.; Tsai, W.J.; Lin, Y.L.; Liao, J.F.; Shen, C.C.; Chen, C.F.; Kuo, Y.C. Inhibitory effects of (S)-armepavine from Nelumbo nucifera on autoimmune disease of MRL lpr/lpr mice. Eur. J. Pharmacol. 2006, 531, 270–279. [Google Scholar] [CrossRef]
- San Miguel, J.F.; Schlag, R.; Khuageva, N.K.; Dimopoulos, M.A.; Shpilberg, O.; Kropff, M.; Spicka, I.; Petrucci, M.T.; Palumbo, A.; Samoilova, O.S.; Dmoszynska, A.; Abdulkadyrov, K.M.; Schots, R.; Jiang, B.; Mateos, M.V.; Anderson, K.C.; Esseltine, D.L.; Liu, K.; Cakana, A.; van de Velde, H.; Richardson, P.G. VISTA Trial Investigators. Bortezomib plus melphalan and prednisone for initial treatment of multiple myeloma. N. Engl. J. Med. 2008, 359, 906–917. [Google Scholar]
- Chou, S.S.P.; Chen, P.W. Cycloaddition reactions of 4-sulfur-substituted dihydro-2-pyridones and 2-pyridones with conjugated dienes. Tetrahedron 2008, 64, 1879–1887. [Google Scholar] [CrossRef]
- Lewis, R.S. Calcium signaling mechanisms in T lymphocytes. Annu. Rev. Immunol. 2001, 19, 497–521. [Google Scholar] [CrossRef]
- Ferreia, V.; Sidenius, N.; Tarantino, N.; Hubert, P.; Chatenoud, L.; Blasi, F.; Körner, M. In vivo inhibition of NF-κB in T-lineage cells leads to a dramatic decrease in cell proliferation and cytokine production and to increased cell apoptosis in response to mitogen stimuli, but not to abnormal thymopoiesis. J. Immunol. 1999, 162, 6442–6450. [Google Scholar]
- Robb, R.J. Interleukin-2: the molecule and its function. Immunol. Today 1984, 5, 203–209. [Google Scholar] [CrossRef]
- Young, H.A. Regulation of interferon-gamma gene expression. J. Interf. Cytok. Res. 1996, 16, 563–568. [Google Scholar] [CrossRef]
- Cantrell, D. T cell antigen receptor signal transduction pathways. Ann. Rev. Immunol. 1996, 14, 259–274. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Yang, N.S.; Chou, C.J.; Lin, L.C.; Tsai, W.J. Regulation of cell proliferation, gene expression, production of cytokines, and cell cycle progression in primary human T lymphocytes by piperlactam S isolated from Piper kadsura. Mol. Pharamcol. 2000, 58, 1057–1066. [Google Scholar]
- Chen, Y.C.; Tsai, W.J.; Wu, M.H.; Lin, L.C.; Kuo, Y.C. Suberosin inhibits human peripheral blood mononuclear cells proliferation through the modulation of NF-AT and NF-κB transcription factors. Brit. J. Pharmacol. 2007, 150, 298–312. [Google Scholar] [CrossRef]
- Charles, A.J., Jr.; Paul, T.; Hunt, S.; Walport, M. Immunobiology. Current Biology Ltd: New York, NY, USA, 1997. [Google Scholar]
- Kuby, J. Immunology.; W.H. Freeman and Company: New York, NY, USA, 1997; pp. 285–310. [Google Scholar]
- Chen, Y.C.; Chang, S.C.; Wu, M.H.; Chuang, K.A.; Wu, J.Y.; Tsai, W.J.; Kuo, Y.C. Norcantharidin reduced cyclins and cytokines production in human peripheral blood mononuclear cells. Life Sci. 2009, 84, 218–226. [Google Scholar] [CrossRef]
- Ajchenbaum, F.; Ando, K.; DeCaprio, J.A.; Griffin, J.D. Independent regulation of human D-type cyclin gene expression during G1 phase in primary human T lymphocytes. J. Biol. Chem. 1993, 268, 4113–4119. [Google Scholar]
- Janeway, C.A.; Travers, P.; Hunt, S.; Walport, M. Immunobiology: The immune system in health and disease; Garland Publishing Inc: New York, NY, USA, 1997; pp. 1:1–1:26. [Google Scholar]
- Wang, S.J.; Chou, S.H.; Kuo, Y.C.; Chou, S.S.P.; Tzeng, W.F.; Leu, C.Y.; Huang, R.F.S.; Liew, Y.F. HDT-1, a new synthetic compound, inhibits glutamate release in rat cerebral cortex nerve terminals (synaptosomes). Acta Pharmacol. Sin. 2008, 29, 1289–1295. [Google Scholar] [CrossRef]
- Wu, M.H.; Tsai, W.J.; Don, M.J.; Chen, Y.C.; Kuo, Y.C. Tanshinlactone A from Salvia miltiorrhiza modulates interleukin-2 and interferon-γ gene expression. J. Ethnopharmacol. 2007, 113, 210–217. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Weng, S.C.; Chou, C.J.; Chang, T.T.; Tsai, W.J. Activation and proliferation signals in primary human T lymphocytes inhibited by ergosterol peroxide isolated from Cordyceps cicadae. Bri. J. Pharmacol. 2003, 140, 895–906. [Google Scholar] [CrossRef]
- Tsai, W.J.; Chen, Y.C.; Wu, M.H.; Lin, L.C.; Chuang, K.A.; Chang, S.C.; Kuo, K.C. Seselin from Plumbago zeylanica inhibits phytohemagglutinin (PHA)-stimulated cell proliferation in human peripheral blood mononuclear cells. J. Ethnopharmacol. 2008, 119, 67–73. [Google Scholar] [CrossRef]
- Taniguchi, T. Structure and function of IL-2 and IL-2 receptors. Behring Inst. Mitt. 1992, 91, 87–95. [Google Scholar]
- Gray, P.W.; Goeddel, D.V. Structure of the human immune interferon gene. Nature 1982, 298, 859–863. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chou, S.-H.; Chou, S.-S.P.; Liew, Y.-F.; Leu, J.-Y.; Wang, S.-J.; Huang, R.-F.S.; Tzeng, W.-F.; Kuo, Y.-C. A New Synthetic Compound, 2-OH, Enhances Interleukin-2 and Interferon-γ Gene Expression in Human Peripheral Blood Mononuclear Cells. Molecules 2009, 14, 2345-2355. https://doi.org/10.3390/molecules14072345
Chou S-H, Chou S-SP, Liew Y-F, Leu J-Y, Wang S-J, Huang R-FS, Tzeng W-F, Kuo Y-C. A New Synthetic Compound, 2-OH, Enhances Interleukin-2 and Interferon-γ Gene Expression in Human Peripheral Blood Mononuclear Cells. Molecules. 2009; 14(7):2345-2355. https://doi.org/10.3390/molecules14072345
Chicago/Turabian StyleChou, Shiu-Huey, Shang-Shing P. Chou, Yih-Fong Liew, Jyh-Yih Leu, Su-Jane Wang, Rwei-Fen S. Huang, Woan-Fang Tzeng, and Yuh-Chi Kuo. 2009. "A New Synthetic Compound, 2-OH, Enhances Interleukin-2 and Interferon-γ Gene Expression in Human Peripheral Blood Mononuclear Cells" Molecules 14, no. 7: 2345-2355. https://doi.org/10.3390/molecules14072345