Salivary Aldehyde Dehydrogenase: Activity towards Aromatic Aldehydes and Comparison with Recombinant ALDH3A1

Abstract
:1. Introduction

2. Results and Discussion
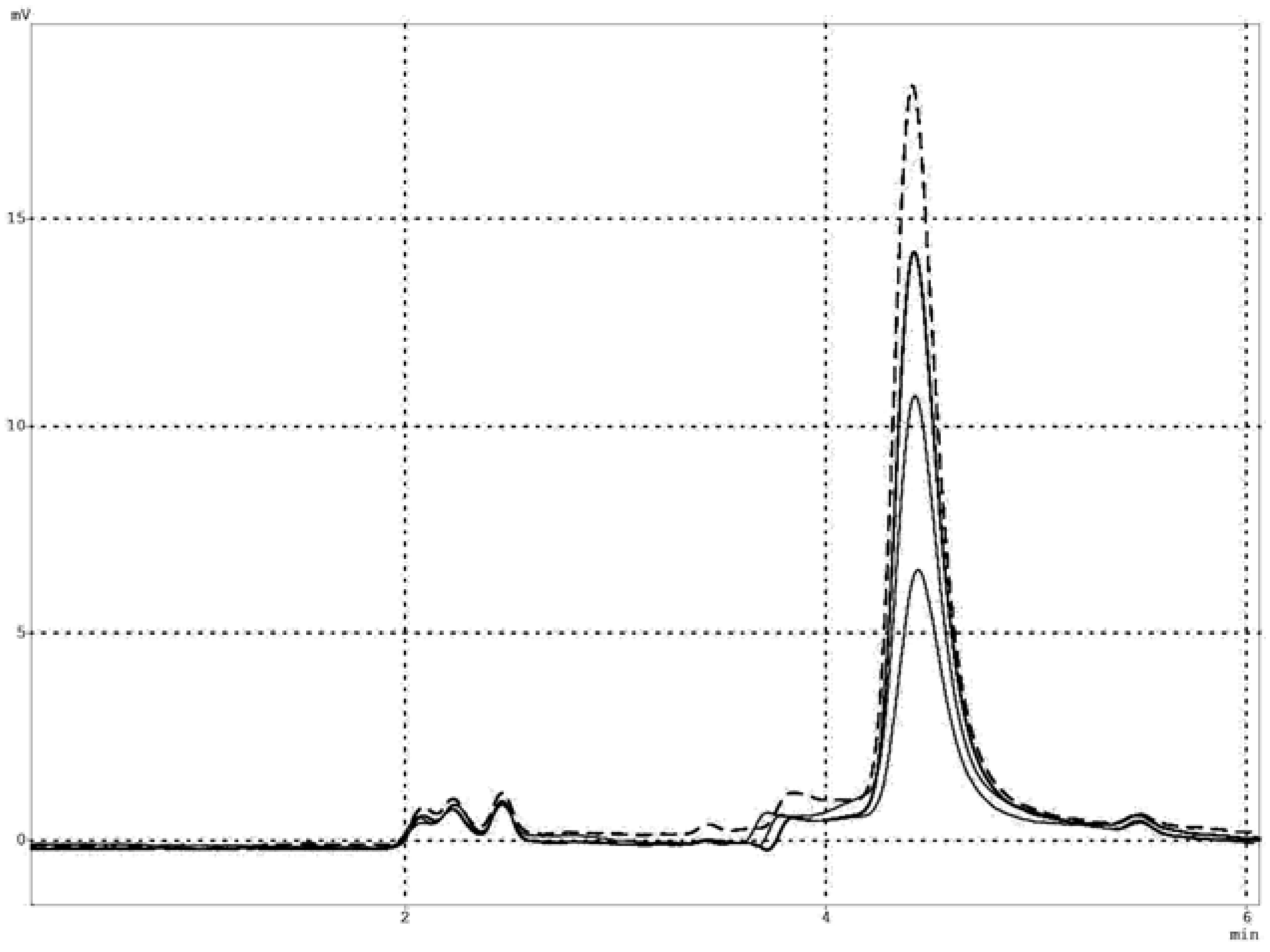
2.1. Salivary ALDH activity – apparent kinetics and product analysis

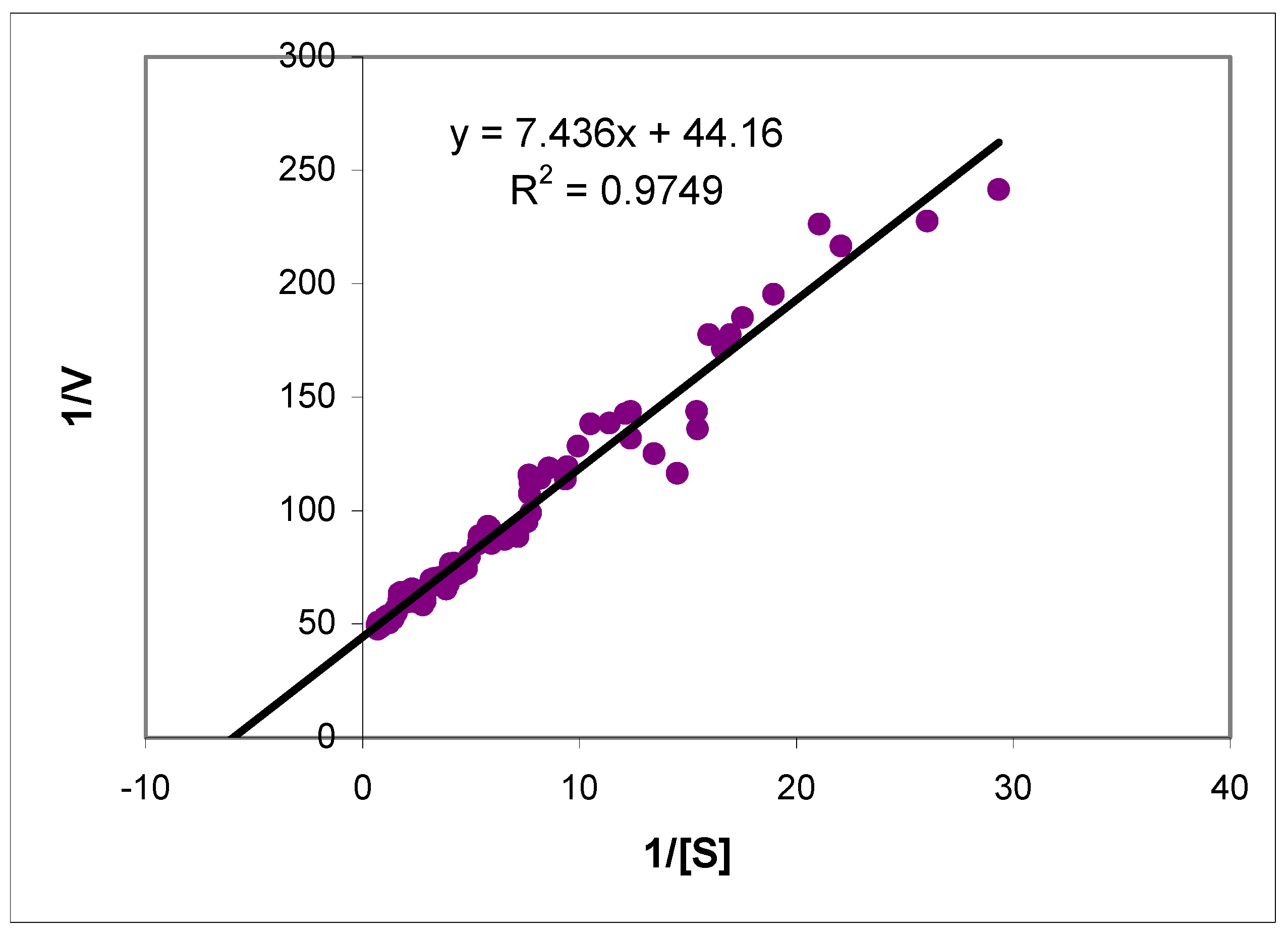
2.2. Kinetic properties of ALDH3A1 towards aromatic aldehydes – comparison with salivary ALDH

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aldehyde | Recombinant ALDH3A1 | salivary ALDH | |||
|---|---|---|---|---|---|
| Km [μM] | Vmax [relative] | Km [μM] | Vmax [relative] | obs [nm] (Δ [M-1cm-1]) | |
| Benzaldehyde | 148 | 100 | 160a | 100 | 340 (6,200) |
| 4-methoxybenzaldehyde (anisaldehyde) | 19 | 73 | nd | ~45 | 340 (6,200) |
| 4-dimethylaminobenzaldehyde | 4.2 | 8 | nd | nd | 350 (-22,000) |
| 4-hydroxy-3-methoxy-benzaldehyde (vanillin) | 155 | 6 | nd | nd | 310 (-7,000) |
| cinnamic aldehyde | 6 | 160 | nd | 190 | 340 (6,200) |
| 2-naphthaldehyde | 0.4 | 101 | 0.46a | 105 | 330 (3,900) |
| 6-methoxy-2-naphthaldehyde (MONAL) | 0.16 | 47 | 0.2a | 52 | 315 (-7,600) |
| 6-dimethylamino-2-naphthaldehyde (DANAL) | ~20 | ~21 | 7.2a | 21a | 380 (-11,000) |
| 1-naphthaldehyde | nd | ~1.5 | - | <25b | 320 (-4,000) |
| 4-methoxy-1-naphthaldehyde | - | <1 | - | <8a | 360 (-9,000) |
| 4-dimethylamino-1-naphthaldehyde | - | <1 | - | <9a | 400 (-10,000) |
| 7-methoxy-1-naphthaldehyde | - | <1 | - | <1a | 360 (-4,000) |


3. Experimental
3.1. General

3.2. Saliva collection
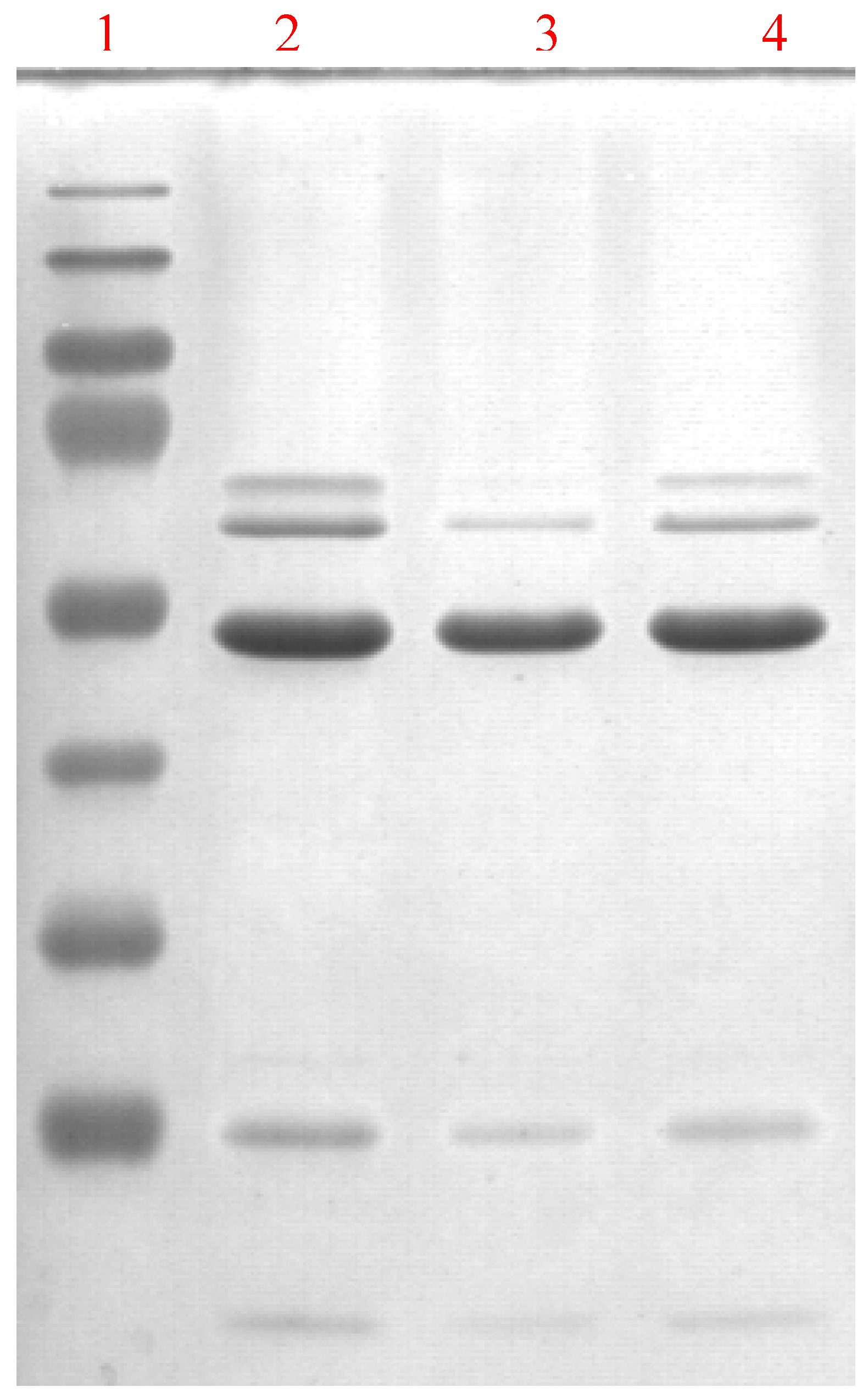
3.3. Cloning of the cDNA for ALDH3A1 and its overexpression

4. Conclusions
Acknowledgements
References and Notes
- Dyck, L.E. Polymorphism of a class 3 aldehyde dehydrogenase present in human saliva and in hair roots. Alcohol. Clin. Exp. Res. 1995, 19, 420–426. [Google Scholar] [CrossRef]
- Feron, V.J.; Til, H.P.; de Vrijer, F.; Woutersen, R.A.; Cassee, F.R.; van Bladeren, P.J. Aldehydes: Occurence, carcinogenic potential, mechanism of action and risk assessment. Mutat. Res. 1991, 259, 363–385. [Google Scholar] [CrossRef]
- Vasiliou, V.; Pappa, A.; Estey, T. Role of human aldehyde dehydrogenases in endobiotic and xenobiotic metabolism. Drug Metab. Rev. 2004, 36, 279–299. [Google Scholar] [CrossRef]
- Sládek, N.E. Human aldehyde dehydrogenases: Potential pathological, pharmacological, and toxicological impact. J. Biochem. Mol. Toxicol. 2003, 17, 7–23. [Google Scholar] [CrossRef]
- Sreerama, L.; Hedge, M.W.; Sladek, N.E. Identification of a class 3 aldehyde dehydrogenase in human saliva and increased levels of this enzyme, glutathione S-transferases, and DT-diaphorase in the saliva of subjects who continually ingest large quantities of coffee or broccoli. Clin. Cancer Res. 1995, 1, 1153–1165. [Google Scholar]
- Ellis, E.M. Reactive carbonyls and oxidative stress: Potential for therapeutic intervention. Pharmacol. Therap. 2007, 115, 13–24. [Google Scholar] [CrossRef]
- Townsend, A.J.; Leone-Kabler, S.; Haynes, R.L.; Wu, Y.; Szweda, L.; Bunting, K.D. Selective protection by stably transfected human ALDH3A1 (but not human ALDH1A1) against toxicity of aliphatic aldehydes in V79 cells. Chem. Biol. Interact. 2001, 130-132, 261–273. [Google Scholar] [CrossRef]
- Pappa, A.; Chen, C.; Koutalos, Y.; Townsend, A.J.; Vasiliou, V. ALDH3A1 protects human corneal epithelial cells from ultraviolet- and 4-hydroxy-2-nonenal-induced oxidative damage. Free Radical Bio. Med. 2003, 34, 1178–1189. [Google Scholar] [CrossRef]
- Estey, T.; Piatigorsky, J.; Lassen, N.; Vasiliou, V. ALDH3A1: a corneal crystallin with diverse functions. Exp. Eye Res. 2007, 84, 3–12. [Google Scholar] [CrossRef]
- Jelski, W.; Szmitkowski, M. Alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) in the cancer diseases. Clin. Chim. Acta 2008, 395, 1–5. [Google Scholar] [CrossRef]
- Sladek, N.E. Aldehyde dehydrogenase-mediated cellular relative insensitivity to the oxazaphosphorines. Curr. Pharm. Des. 1999, 5, 607–625. [Google Scholar]
- Hsu, L.C.; Chang, W.C.; Hiraoka, L.; Hsieh, C.L. Molecular cloning, genomic organization, and chromosomal localization of an additional human aldehyde dehydrogenase gene, ALDH6. Genomics 1994, 24, 333–341. [Google Scholar]
- Aldehyde dehydrogenase gene superfamily database. Available online: www.aldh.org.
- Pappa, A.; Estey, T.; Manzer, R.; Brown, D.; Vasiliou, V. Human aldehyde dehydrogenase 3A1 (ALDH3A1): Biochemical characterization and immunohistochemical localization in the cornea. Biochem. J. 2003, 376, 615–623. [Google Scholar] [CrossRef]
- Wierzchowski, J.; Wroczynski, P.; Laszuk, K.; Interewicz, E. Fluorimetric detection of aldehyde dehydrogenase activity in human blood, saliva and organ biopsies, and kinetic differentiation between class I and class III isozymes. Anal. Biochem. 1997, 245, 68–75. [Google Scholar]
- Wroczyński, P.; Wierzchowski, J. Aromatic aldehydes as fluorogenic indicators for human aldehyde dehydrogenases and oxidases. Analyst 2000, 125, 511–516. [Google Scholar]
- Wroczyński, P.; Wierzchowski, J.; Rakowska, A.; Chimkowska, M.; Targoński, J. Aldehyde dehydrogenase in human saliva – evaluation of its oxidation status. Acta Pol. Pharm. 2004, 61 Suppl., 62–64. [Google Scholar]
- Wierzchowski, J.; Pietrzak, M.; Szeląg, M.; Wroczyński, P. Salivary aldehyde dehydrogenase – reversible oxidation of the enzyme and its inhibition by caffeine, investigated using fluorimetric method. Arch. Oral Biol. 2008, 53, 423–428. [Google Scholar] [CrossRef]
- Wierzchowski, J.; Interewicz, E.; Wroczynski, P.; Orlanska, I. Continuous fluorimetric assay for human aldehyde dehydrogenase and its application to blood analysis. Anal. Chim. Acta 1996, 319, 209–219. [Google Scholar] [CrossRef]
- Glatt, H.; Rost, K.; Frank, H.; Seidel, A.; Kollock, R. Detoxification of promutagenic aldehydes derived from methylpyrenes by human aldehyde dehydrogenases ALDH2 and ALDH3A1. Arch. Biochem. Biophys. 2008, 477, 196–205. [Google Scholar] [CrossRef]
- Seitz, H.K.; Matsuzaki, S.; Yokoyama, A.; Homann, N.; Vaekaevainen, S.; Wang, X.D. Alcohol and cancer. Alcohol. Clin. Exper. Res. 2001, 25, 137–143. [Google Scholar] [CrossRef]
- Sládek, N.E.; Kollander, R; Sreerama, L.; Kiang, D.T. Cellular levels of aldehyde dehydrogenases (ALDH1A1 and ALDH3A1) as predictors of therapeutic responses to cyclophosphamide-based chemotherapy of breast cancer: A retrospective study. Cancer Chemother. Pharmacol. 2002, 49, 309–321. [Google Scholar] [CrossRef]
- Moreb, S.J.; Muhoczy, D.; Ostmark, B.; Zucali, J.R. RNAi-mediated knockdown of aldehyde dehydrogenase class-1A1 and class-3A1 is specific and reveals that each contributes equally to the resistance against 4-hydroperoxycyclophosphamide. Cancer Chemother. Pharmacol. 2007, 59, 127–136. [Google Scholar]
- Ho, K.K.; Mukhopadhyay, A.; Li, Y.F.; Mukhopadhyay, S.; Weiner, H. A point mutation produced a class 3 aldehyde dehydrogenase with increased protective ability against the killing effect of cyclophosphamide. Biochem. Pharmacol. 2008, 76, 690–696. [Google Scholar] [CrossRef]
- Ekhart, C.; Rodenhuis, S.; Smits, P.H.M.; Beijnen, J.H.; Huitema, A.D.R. Relations between polymorphisms in drug-metabolising enzymes and toxicity of chemotherapy with cyclophosphamide, thiotepa and carboplatin. Pharmacogenet. Genomics 2008, 18, 1009–1015. [Google Scholar] [CrossRef]
- Samples Availability: Available from the authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Giebułtowicz, J.; Wolinowska, R.; Sztybor, A.; Pietrzak, M.; Wroczyński, P.; Wierzchowski, J. Salivary Aldehyde Dehydrogenase: Activity towards Aromatic Aldehydes and Comparison with Recombinant ALDH3A1. Molecules 2009, 14, 2363-2372. https://doi.org/10.3390/molecules14072363
Giebułtowicz J, Wolinowska R, Sztybor A, Pietrzak M, Wroczyński P, Wierzchowski J. Salivary Aldehyde Dehydrogenase: Activity towards Aromatic Aldehydes and Comparison with Recombinant ALDH3A1. Molecules. 2009; 14(7):2363-2372. https://doi.org/10.3390/molecules14072363
Chicago/Turabian StyleGiebułtowicz, Joanna, Renata Wolinowska, Anna Sztybor, Monika Pietrzak, Piotr Wroczyński, and Jacek Wierzchowski. 2009. "Salivary Aldehyde Dehydrogenase: Activity towards Aromatic Aldehydes and Comparison with Recombinant ALDH3A1" Molecules 14, no. 7: 2363-2372. https://doi.org/10.3390/molecules14072363