Vitamin B6: A Molecule for Human Health?

Washington State University, Abelson 435, P.O. Box 66224, Pullman, WA, USA
*
Author to whom correspondence should be addressed.
Molecules 2010, 15(1), 442-459; https://doi.org/10.3390/molecules15010442
Submission received: 5 November 2009
/
Revised: 16 January 2010
/
Accepted: 20 January 2010
/
Published: 20 January 2010
(This article belongs to the Special Issue Vitamins)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Vitamin B6 is an intriguing molecule that is involved in a wide range of metabolic, physiological and developmental processes. Based on its water solubility and high reactivity when phosphorylated, it is a suitable co-factor for many biochemical processes. Furthermore the vitamin is a potent antioxidant, rivaling carotenoids or tocopherols in its ability to quench reactive oxygen species. It is therefore not surprising that the vitamin is essential and unquestionably important for the cellular metabolism and well-being of all living organisms. The review briefly summarizes the biosynthetic pathways of vitamin B6 in pro- and eukaryotes and its diverse roles in enzymatic reactions. Finally, because in recent years the vitamin has often been considered beneficial for human health, the review will also sum up and critically reflect on current knowledge how human health can profit from vitamin B6.
1. Introduction
The B vitamins are a group of water soluble, chemically quite distinct compounds to which other than vitamin B6, vitamin B1 (thiamine), B2 (riboflavin), B3 (niacin or niacin amide), B5 (pantothenic acid), B7 (biotin), B9 (folic acid), and B12 (various cobalamins) also belong [1]. Historically, it was believed that only one vitamin B existed with a critical function for maintenance of growth and health and prevention of characteristic skin lesions in animals and human [2]. However, with ongoing research it became obvious that vitamin B actually comprised a group of compounds that was collectively called the ‘vitamin B complex’.
Vitamin B6 (vitB6 from here on) itself is an enzymatic co-factor required for more than 140 biochemical reactions including transaminations, aldol cleavages, α-decarboxylations, racemizations, β- and γ- eliminations, and replacement reactions. Most of these reactions are related to amino acid biosynthesis and degradation, but vitB6 is also involved in other processes including sugar and fatty acid metabolism [3]. It comprises a set of three different pyridine derivatives called pyridoxine (PN; 1), pyridoxal (PL; 2), and pyridoxamine (PM; 3). They differ in a variable group present at their 4-position with PN carrying a hydroxymethyl group, and PL (2) and PM (3) having an aldehyde and an aminomethyl group, respectively. Furthermore, all three B6 vitamers are phosphorylated by a kinase, which is a requirement for their role as cofactors in enzymatic reactions (Scheme 1). While pyridoxamine-5’-phosphate (PMP; 4) has been reported to function as a co-factor, it is pyridoxal 5’-phosphate (PLP; 5) that is the biologically most active form [4,5].
A growing number of interesting and helpful new resources have been established in the last years that focus primarily on vitB6 related issues. For example, an online database has been launched that allows searching whole genomes for PLP-dependent enzymes, and which also provides information on critical aspects such as the biochemical pathways requiring PLP (5) and the classification of PLP-dependent enzymes (http://bioinformatics.unipr.it/cgi-bin/bioinformatics/B6db/home.pl) [3]. In addition, a database has been established that allows searching for mutated PLP-dependent enzymes in various organisms (http://www.studiofmp.com/plpmdb/home.htm) [6].
2. Suggested Reaction Mechanisms of VitB6 for Amino Acid Metabolism
In most cases PLP (5) is covalently bound to the ε-amino group of a conserved lysine residue in the active center of a PLP-dependent enzyme, with its 5’-phosphate group being buried in a conserved phosphate-binding cup [7]. It is suggested that reactions are initiated by the formation of a geminal diamine intermediate between the aldehydic carbon atom of PLP (5) and an amino group of the substrate. This is followed by its rapid breakdown and the formation of an external aldimine (Schiff base) between PLP (5) and the substrate causing the release of the lysine residue of the enzyme from PLP (5). From this point on subsequent reactions mainly depend on the specific, participating enzymes that guide and modulate the next steps leading to e.g. racemisations, β- and γ- eliminations.
3. Three Different Biosynthetic Pathways for VitB6 Are Known
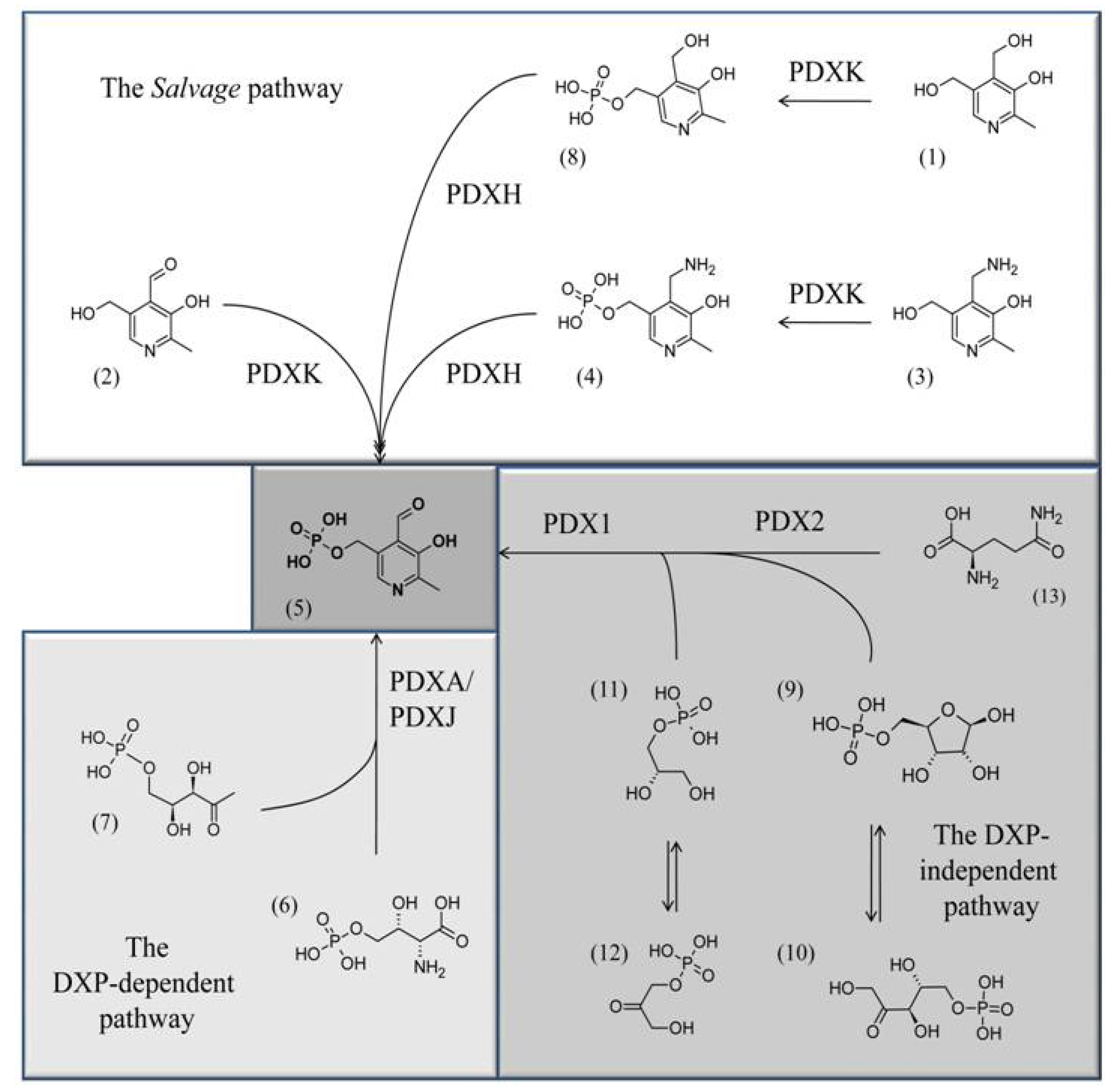
Three different pathways for vitB6 biosynthesis have been described which will be just briefly summarized, as they were topics of other recent reviews [8,9]. In eubacteria like Escherichia coli, the vitamin can be de novo synthesized by the concerted activities of the pyridoxine biosynthesis proteins A and J (PdxA (EC 1.1.1.262) and PdxJ (EC 2.6.99.2), respectively) which use 4-phospohydroxy-L-threonine (4HPT; 6) and deoxyxylose 5’-phosphate (DXP; 7) to synthesize pyridoxine 5’-phosphate (PNP; 8) (Scheme 1) [10,11,12]. In bacteria, archaea, and eukarya a second de novo pathway is known that synthesizes PLP (5) from ribose 5’-phosphate (9) or ribulose 5’-phosphate (10), in combination with either glyceraldehyde 3’-phosphate (11) or dihydroxyacetone phosphate (12) and glutamine (13) (Scheme 1) [13,14,15,16,17].
Scheme 1.
The three known pathways for PLP biosynthesis: one salvage pathway, and two de novo pathways, a DXP-dependent one and a DXP-independent one. Chemical structures: (5) PLP; (7) deoxyxylulose 5’-phosphate, (6) 4-(phosphohydroxy)-L-threonine; (11) glyceraldehyde 3’-phosphate; (12) dihydroxyacetone phosphate; (9) ribose 5’-phosphate; (10) ribulose 5’-phosphate, (13) glutamine, (3) PM, (4) PMP, (1) PN, (8) PNP, (2) PL.
Scheme 1.
The three known pathways for PLP biosynthesis: one salvage pathway, and two de novo pathways, a DXP-dependent one and a DXP-independent one. Chemical structures: (5) PLP; (7) deoxyxylulose 5’-phosphate, (6) 4-(phosphohydroxy)-L-threonine; (11) glyceraldehyde 3’-phosphate; (12) dihydroxyacetone phosphate; (9) ribose 5’-phosphate; (10) ribulose 5’-phosphate, (13) glutamine, (3) PM, (4) PMP, (1) PN, (8) PNP, (2) PL.

Here two pyridoxine biosynthetic enzymes (PDX) are active: while PDX2 functions as a glutaminase that deaminates glutamine to glutamate in order to supply nitrogen for the PLP heterocycle, PDX1 arranges the final ring closure [18,19,20,21,22,23,24]. Because of a different sugar precursor used for the biosynthesis of the vitamin, the de novo pathway from eubacteria is known as the DXP-dependent pathway, while the other is the DXP-independent pathway [14]. In addition to the two de novo pathways, most organisms also have a salvage pathway that converts the different B6 vitamers to PLP (5). This is achieved by the concerted activities of an oxidase, PDXH (EC 1.4.3.5), and a kinase, PDXK (EC 2.7.1.35) (Scheme 1) [8,25]. Most animal organisms, including humans, have a salvage pathway, however, they lack the enzymatic machinery for de novo synthesis and rely on external uptake of the vitamin from food [16].
4. VitB6 and Its Healthy Face
Since its discovery in 1932 by the Japanese scientist S. Ohdake, vitB6 has been discussed in relationship to health issues [26]. In these early works from, for example, Ohdake or the Hungarian scientist P. Györgi, vitB6 was associated with pellagra, a skin disease that is based on multi-vitamin deficiencies that mostly occurs in context with niacin undersupply [27,28,29]. A search through the public literature data basis (http://www.ncbi.nlm.nih.gov/) for health aspects associated with vitB6 yields a surprisingly high number of articles (>900). Furthermore, the current Recommended Dietary Allowance per day by the National Institute of Health (NIH) of the USA is around 2 mg with an upward tolerance of 100 mg per day for adults. A recent U.S. study, which tested the blood PLP levels in around 8,000 patients, demonstrated a widespread deficiency of the vitamin among all tested subgroups, and the authors suggested an increase of the daily allowance from around 2 mg to 3 to 4.9 mg per day [30]. It has been reported for animal models, that continuous uptake of very high doses (e.g. 400 mg/kg) can lead to peripheral sensory neuropathy and nerve degeneration [31,32]. These problems are generally reversible when supplementation is stopped. Additionally some studies have suggested that increased levels of the B6 vitamers and some derivatives can generate toxic photoproducts as a result of UV irradiation [33,34,35]. However, the applied daily dosages were far beyond any physiological concentrations an organism is normally exposed to, making it unlikely that such vitB6 induced impacts will be observed. Because of the great interest in vitB6 as a therapeutic and pharmaceutical compound, its reactive capability, and its potent antioxidative characteristics, we summarize in the following paragraphs some of the relevant topics related to these issues.
4.1. Therapeutic applications by using drugs against PLP-dependent enzymes
PLP-dependent enzymes are highly diverse and the reactions they facilitate are estimated to represent 4% of all known catalytic activities; hence, many of them are being explored as targets for therapeutic agents (for an excellent overview see [5]). We chose three major examples for this review to illustrate the potentials of this approach in disease control: malaria, sleeping sickness, and cancer treatment.
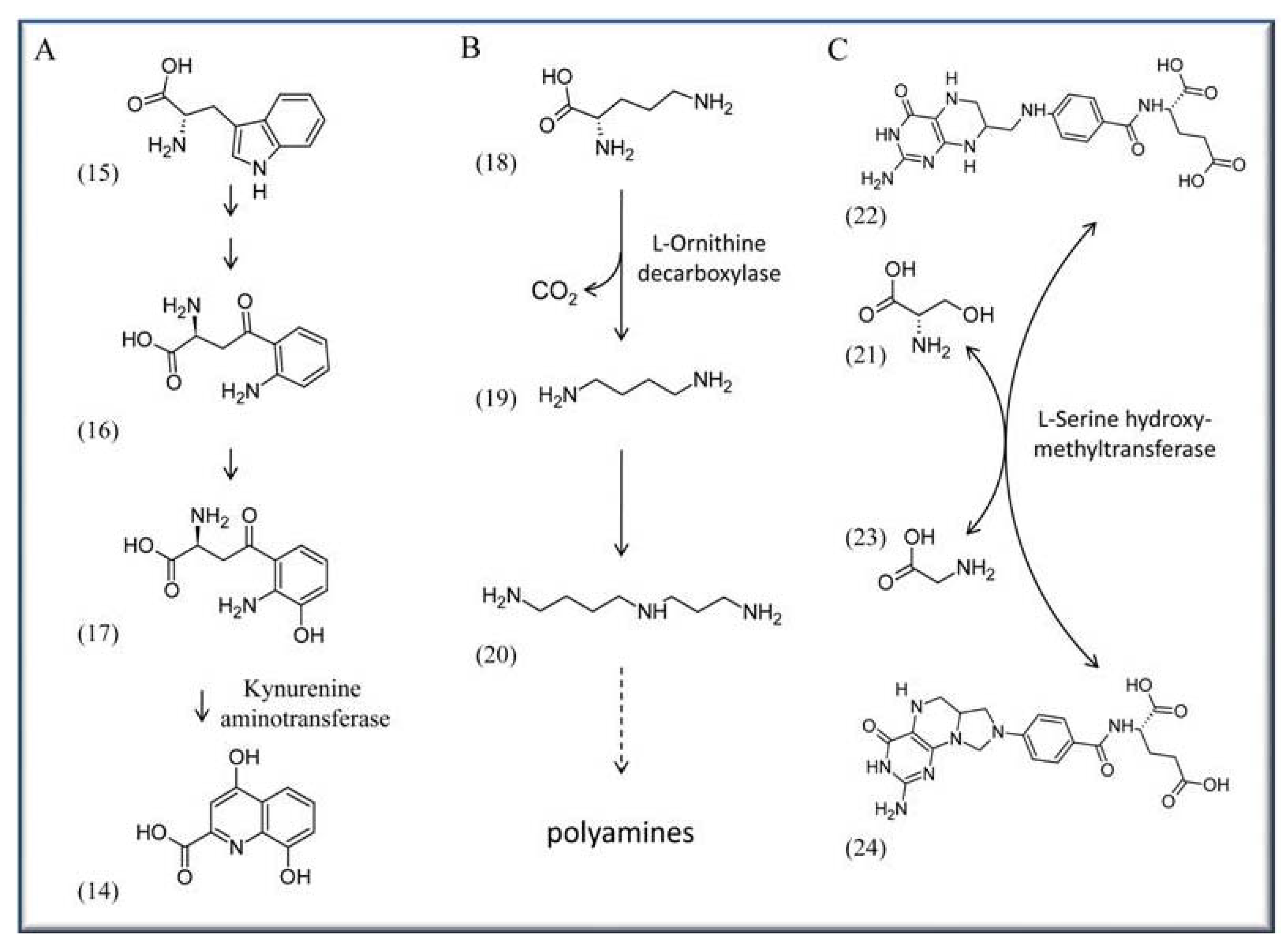
One of the most threatening human diseases is malaria, with more than 300-500 million infected people worldwide and an annual death toll of up to one million people (http://www.unicef.org/health/ index_malaria.html). Several approaches are currently underway in an effort to affect the life cycle or metabolism of the pathogen Plasmodium falciparum, the cause of malaria. One such approach is to impair biosynthesis of xanthurenic acid (14), which is essential for gametogenesis and fertility of the pathogen [36,37,38]. The acid is synthesized as part of the L-tryptophan (15) degradation pathway from L-kynurenine (16) via 3-hydroxykynurenine (17) by the activity of the PLP-dependent kynurenine aminotransferase (EC 2.6.1.7) [39] (Scheme 2A).
A possible strategy involves developing specific drugs that reduce activity of the aminotransferase. This might lower the levels of 3-hydroxykynurenine (17) in P. falciparum infected mosquitoes potentially reducing or even preventing malaria transmission to humans. A similar direction was recently proposed by channeling synthetic pyridoxyl-amino acid adducts into the pathogen, which can phosphorylate these compounds mediated by PDXK kinase [40]. After binding by a PLP-dependent enzyme, such phosphorylated compounds should inhibit these enzymes and affect further metabolism. Müller and co-workers successfully tried pyridoxyl-tryptophan methyl ester to inhibit proliferation of P. falciparum opening up the possibility for a novel malaria treatment in the future [40]. Because P. falciparum expresses PDX1/PDX2 proteins, which humans lack, a potential approach can also be to target these de novo pathway proteins by specific drugs [41]. However, no specific approach has been reported so far.
African sleeping sickness is another severe epidemic disease with an estimated 300–500 thousand people affected in various African countries (http://www.sbri.org/diseases/african.asp). It is caused by the protist Trypanosoma brucei and transmitted by flies of the Genus Glossina. A target to treat sleeping sickness in affected patients is the PLP-dependent enzyme ornithine decarboxylase (ODC; E.C. 4.1.1.17). It catalyzes the step from L-ornithine (18) to the diamine putrescine (19), an initial step in the production of polyamines (Scheme 2B). α-Difluoromethylornithine (DFMO) is a proven irreversible inhibitor of ODC activity and works by forming a covalent bond with a cysteine residue of ODC after decarboxylation [42,43,44]. Although DFMO is an approved drug in treating sleeping sickness caused by T. brucei, the precise reason for its effectiveness is not fully resolved because human and T. brucei ODCs are comparably affected by the agent [45]. It is suggested that this effect is based on metabolic differences: a more rapid turnover of the host’s ODC on the one side, and on the other side T. brucei’s high demand for the synthesis of the polyamine trypanothione, a specific dithiol essential for the detoxification system of Trypanosomes and Leishmania parasites [46,47].
Targeting PLP-dependent enzymes is also discussed in context with cancer. Here an interesting candidate is, for example, serine hydroxymethyltransferase (SHMT; EC 2.1.2.1), which catalyzes the reversible transfer of the Cβ of serine (21) to tetrahydrofolate (22) to form glycine (23) and 5,10-methylenetetrahydrofolate (24) (Scheme 2C). Because of 5,10-methylenetetrahydrofolate (24), which serves as a methyl donor in many reactions, SHMT activity is critical for one-carbon metabolism, the biosynthesis of methionine, lipids, formyl-tRNA and pyrimidine. The latter is of special interest as apparently SHMT activity is coupled to some extent with increased demand for DNA biosynthesis. For example there is evidence in tumors with highly proliferating, mitotically active cells, that serine is preferentially channeled for DNA biosynthesis [48,49]. Consequently SHMT is a proposed target in developing drugs for chemotherapy [50,51].
Scheme 2.
Enzymatic reaction that are targets for pharmaceutical approaches. Shown are only the PLP-dependent enzymes. (A) Synthesis of xanthurenic acid (14) from L-tryptophan (15), via the intermediates L-kynurenine (16) and 3-hydroxykynurenine (17). (B) Synthesis of putrescine (19) from L-ornithine (18), leading subsequently to the synthesis of spermidine (20) and other polyamines. (C) Synthesis of tetrahydrofolate (22) and serine (21) to glycine (23) and 5,10-methylenetetrahydrofolate (24). Note: in this and the next Schemes only the PLP-dependent enzymes are shown.
Scheme 2.
Enzymatic reaction that are targets for pharmaceutical approaches. Shown are only the PLP-dependent enzymes. (A) Synthesis of xanthurenic acid (14) from L-tryptophan (15), via the intermediates L-kynurenine (16) and 3-hydroxykynurenine (17). (B) Synthesis of putrescine (19) from L-ornithine (18), leading subsequently to the synthesis of spermidine (20) and other polyamines. (C) Synthesis of tetrahydrofolate (22) and serine (21) to glycine (23) and 5,10-methylenetetrahydrofolate (24). Note: in this and the next Schemes only the PLP-dependent enzymes are shown.

4.2. VitB6 in context with cardiovascular disease and blood pressure
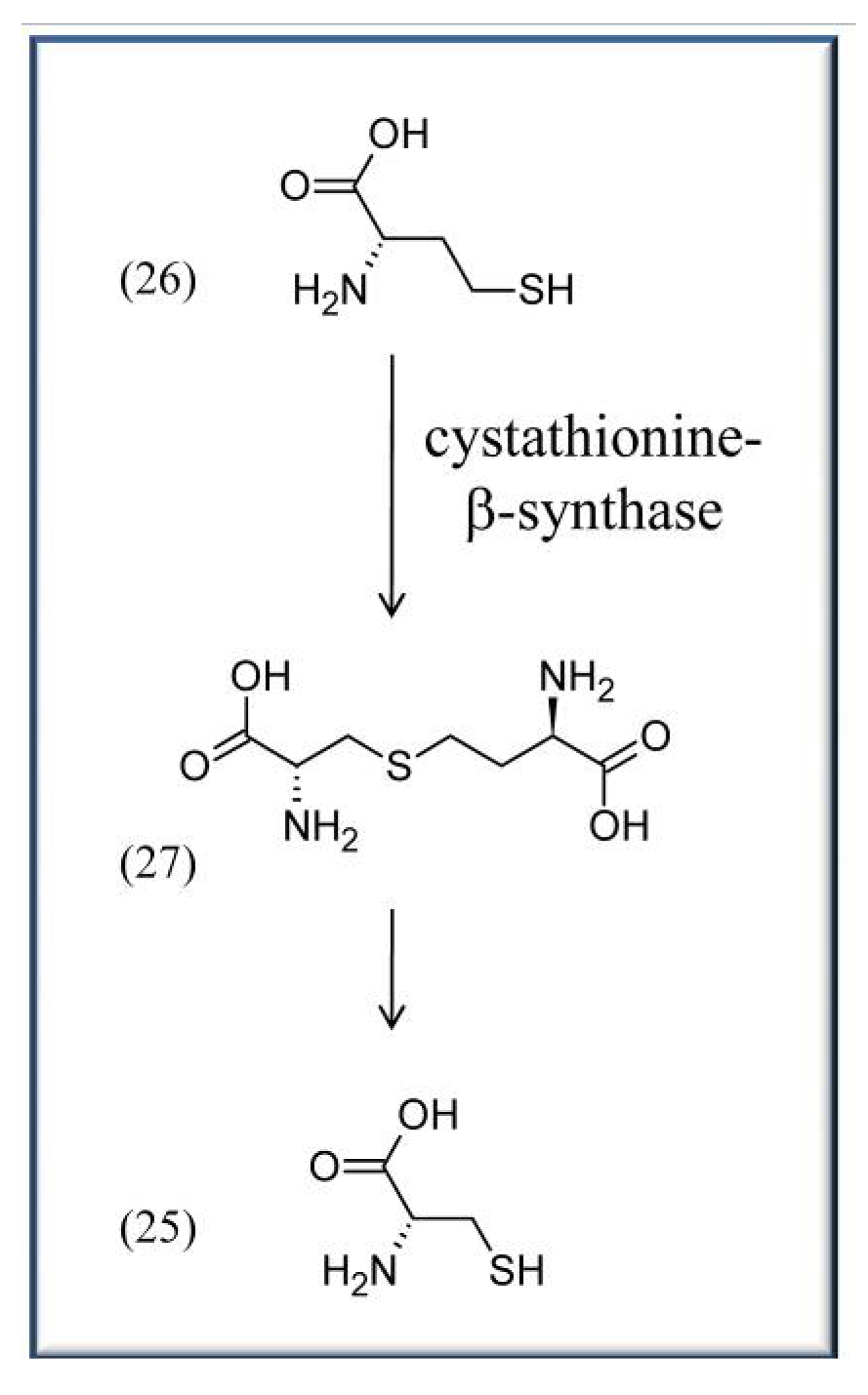
Other aspects in which vitamin B6 is directly discussed to play an important role are cardiovascular disease and high blood pressure. Coronary heart disease (CHD) is one the major reasons for death worldwide. It is caused by atheromata, which are swollen artery walls due to the accumulation of cell debris containing e.g., fatty acids and cholesterols that negatively affect blood flow. Though the impact of vitB6 is controversially discussed (compare for example [52] and [53]), a variety of works indicate positive effects of vitB6 on CHD. For instance, a large study in Japan, comprising 40,803 subjects, recently showed that vitB6 has the potential to reduce the risk of CHD, and especially nonfatal myocardial infarction (MI), among middle-aged (40–59 years) non-multivitamin supplement users [53]. Here, an increase of daily supplementary vitB6 intake from 1.3 to 1.6 mg already significantly reduced the number of affected patients with reported CHDs and MIs [53]. Similarly, the Coronary Health Project and other studies indicate a correlation between increased vitB6 intake and reduced risk of CHD [25,54,55,56]. It is noteworthy that often other vitamins like folates or cobalamins are tested in these studies as well with similar positive effects in reducing the risk of CHD. The precise reason(s) for the beneficial impact of vitB6 are unclear. One suggested reason is that vitB6, like folates and cobalamins, can lower homocysteine (26) levels in the blood by converting the amino acid to cysteine (25) or methionine, respectively. VitB6 is required as a cofactor for cystathionine-β-synthase (EC 4.2.1.22), a PLP-dependent enzyme that converts homocysteine (26) to cysteine (25) via a cystathionine (27) intermediate (Scheme 3) [57]. Because high levels of homocysteine are often associated with an increased chance for atherosclerotic diseases, it is considered a risk factor like e.g. high blood pressure, active smoking, or adverse blood lipid profiles [58]. But, as stressed above, it is not generally accepted whether vitB6, folates, or cobalamins do indeed reduce the blood homocysteine levels, as a recent review indicates, thus still awaiting additional proof [39].
Scheme 3.
Synthesis of L-cysteine (25) from L-homo-cysteine (26) via the intermediate L-cystathionine (27).
Scheme 3.
Synthesis of L-cysteine (25) from L-homo-cysteine (26) via the intermediate L-cystathionine (27).

VitB6 appears also to have a beneficial role in reducing hypertension or high blood pressure. Several articles showed that supplementary treatments with the vitamin could lower blood pressure [59,60,61,62,63,64]. Like for CHD the biochemical or physiological reasons are unresolved. However, it is suggested that the blood pressure lowering role of vitB6 might be connected with the level of blood aldehydes. These are highly reactive compounds that, potentially by binding to sulfhydryl groups of membrane proteins, activate Ca2+-channels and increase cytosolic free calcium in the blood, ultimately leading to an increase in peripheral vascular resistance and blood pressure [65]. Consequently, it is not uncommon for people with excessive alcohol consumption to have increased levels of acetaldehyde, which is often accompanied by high blood pressure [66]. Treatment with N-acetylcysteine can normalize blood pressure in spontaneous hypertensive rats, most likely because the amino acid competes with membrane proteins for the reaction with the aldehydes, causing a reduced Ca2+ flux [67]. In addition, it is well established that acetaldehyde is detrimental to PLP (5) stability [68]. Because PLP (5) is needed for the biosynthesis of cysteine (25), it is suggested that the mechanism of PLP (5) on the blood pressure is either a direct one, by buffering the detrimental activity of aldehydes, or occurs indirectly, by influencing the rate of cysteine (25) biosynthesis [61,63]. Still, it is worth mentioning that the connection of PLP (5) with lowered blood pressure is curious as it is required for dopamine biosynthesis (see Section 4.3 below), a known vasopressor that actually stimulates contraction of the muscular tissue of the capillaries and arteries [69]. A correlation between high blood pressure and PDXH/Pnpo gene expression was found in hypertensive Dahl-S rats [64]. These rats are sensitive to a high salt diet and develop high blood pressure in response to such foods [70]. Okuda and co-workers could show for Dahl-S rats supplied with a high salt diet that, in comparison to a control group, gene expression of the oxidase is down-regulated. These findings indicate that high blood PLP (5) levels are needed for coping with salt uptake and argue that the vitamin is beneficial for preventing high blood pressure [64]. However, it would be interesting to know to what extent neurotransmitter production in these rats is affected as well and whether this correlates with the hypertensive status.
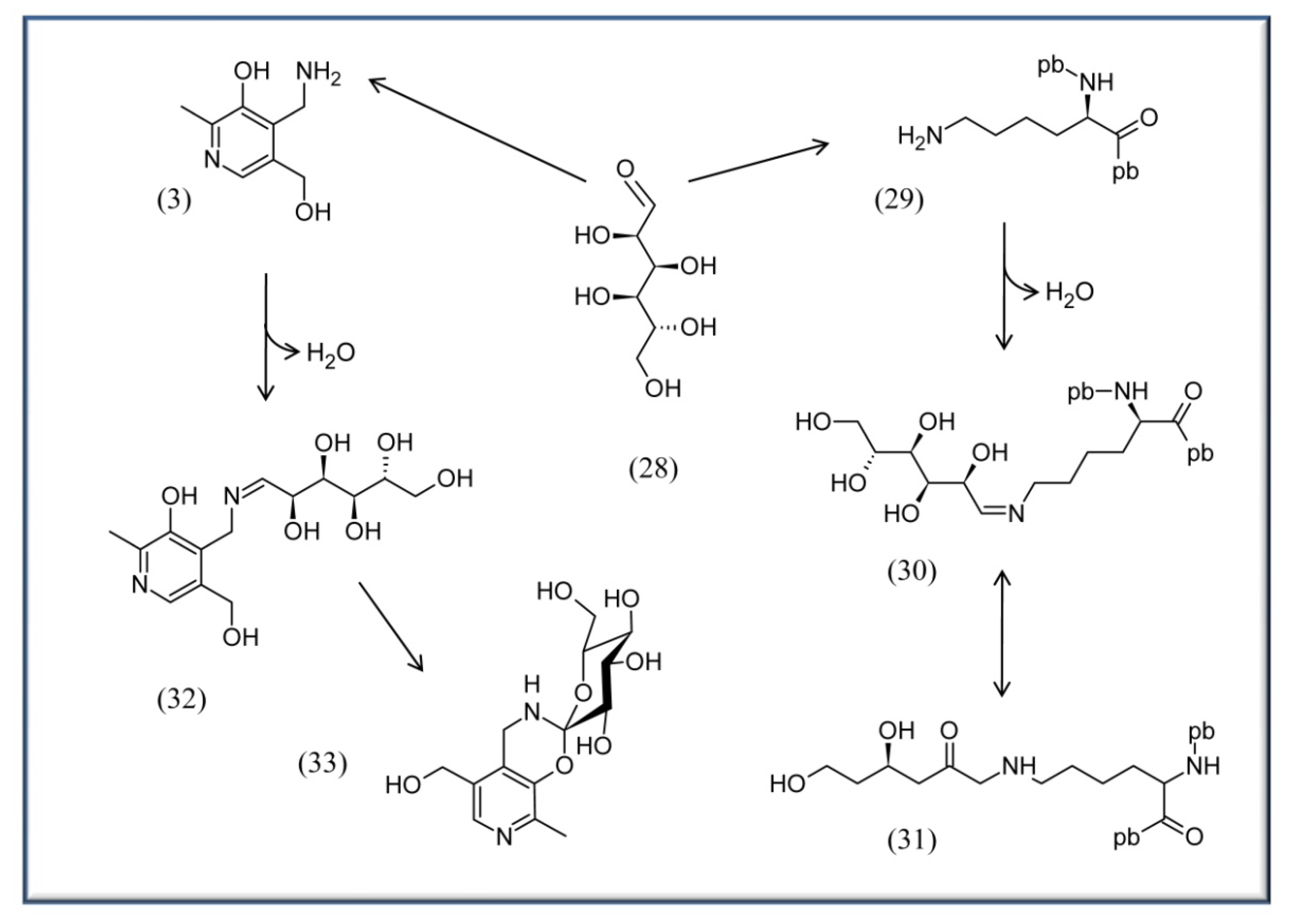
Scheme 4.
Example for Advanced Glycation Endproduct Reaction of glucose with proteins or PM. On the right half: glucose (28) can react with lysine residues of proteins 29 to form an imine (Schiff base) 30 and an Amadori product 31. This latter can be further oxidized by metals to form the final AGE. pb, peptide bond. On the left half: PM (3) has been proven to be the most potent B6 vitamer to compete for the formation of AGEs [73]. It is suggested to form first an imine 32 with the glucose, followed by a double cyclization to afford 33 rather than formation of an Amadori product [73].
Scheme 4.
Example for Advanced Glycation Endproduct Reaction of glucose with proteins or PM. On the right half: glucose (28) can react with lysine residues of proteins 29 to form an imine (Schiff base) 30 and an Amadori product 31. This latter can be further oxidized by metals to form the final AGE. pb, peptide bond. On the left half: PM (3) has been proven to be the most potent B6 vitamer to compete for the formation of AGEs [73]. It is suggested to form first an imine 32 with the glucose, followed by a double cyclization to afford 33 rather than formation of an Amadori product [73].

4.3. VitB6 in context with diabetes, AGE and ALE
A variety of articles about diabetes mellitus focus on the impact of vitB6 on blood sugar levels and arteriosclerosis [60,71,72]. For example, a recent study could show that endothelial dysfunction is normalized by treatment with folates and vitB6 in children with type 1 diabetes [71]. Endothelial dysfunction is an indicator for the progression of arteriosclerosis that is often developed early in diabetes mellitus patients. Endothelial function can be assessed as flow-mediated dilation of the brachial artery with high-resolution ultrasound. MacKenzie and co-workers treated patients over eight weeks with vitB6 or folate, which resulted in improved flow-mediated dilation from an average 3.5% to 8.3% and 2.6% to 9.7%, respectively, and with a combination of both to over 10% [71].
Other work also supports the notion of a positive impact of vitB6 on endothelial cells, indicating that the vitamin is indeed affecting the status of this tissue [74,75,76]. Additionally vitB6 seems to have a positive role against progressive kidney disease, which is frequently associated with diabetic nephropathy [72,77,78]. A possible reason for the advantageous results of vitB6 on mammalian tissues is discussed to be the vitamin’s ability to react with reducing sugar and lipids in the blood to prevent formation of advanced glycation or lipoxygenation endproducts (AGE and ALE, respectively) (Scheme 4) [39,72,73,79].
Such products can accumulate when reducing sugars like glucose (28) or fructose or polyunsaturated fatty acids are highly abundant in the blood or in cells. This can be the case under stress conditions (e.g. oxidative stress) or for patients that suffer from diabetes or arteriosclerosis, respectively. The accumulation of AGEs and ALEs are on the long run detrimental and can lead to severe tissue damage in the body. Here, vitB6 might effectively prevent AGE and ALE formation making it a good candidate as a therapeutic agent in treating side effects in diabetes and arteriosclerosis patients [80,81].
4.4. VitB6 in context with neurological activity
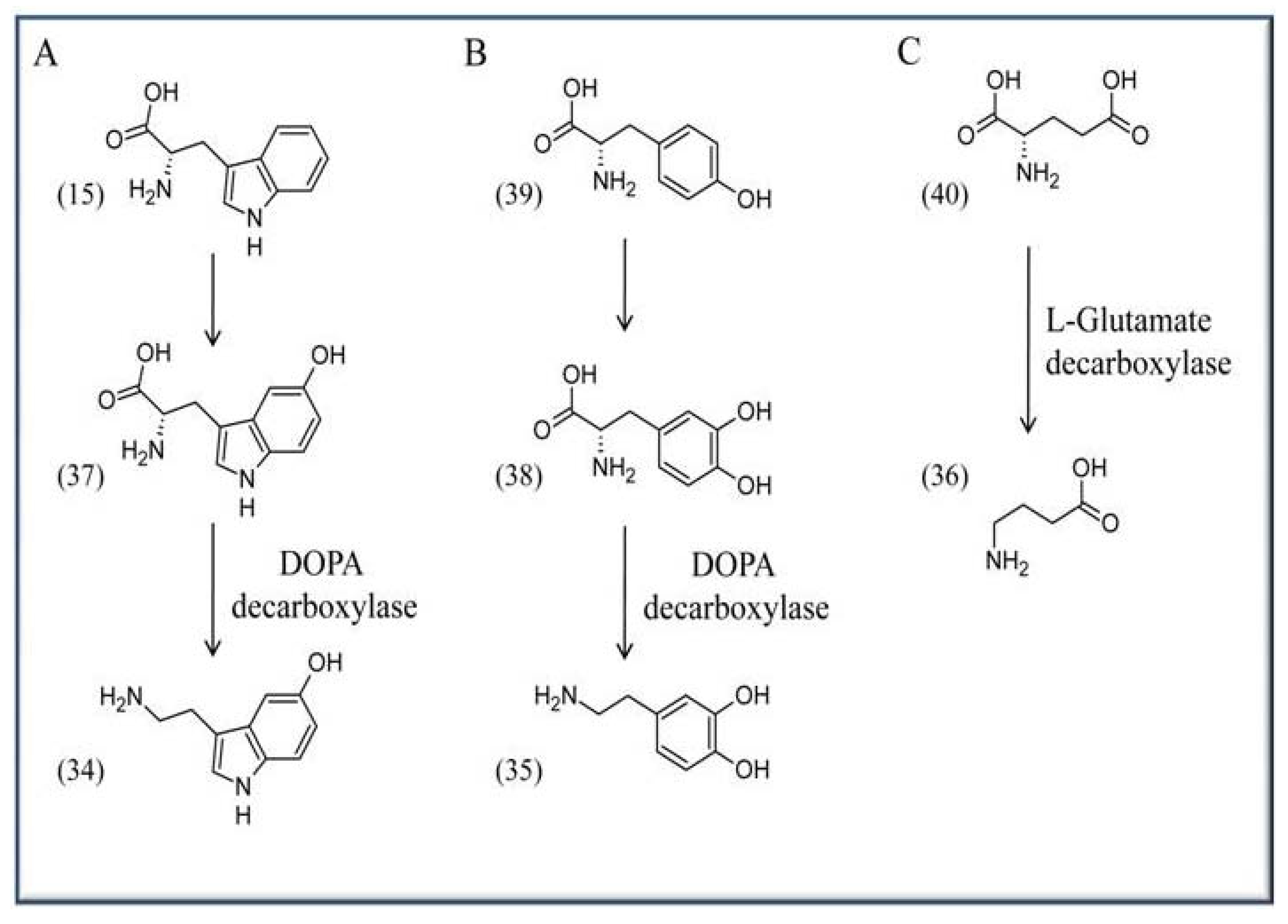
VitB6 is required for the biosynthesis of several neurotransmitters like serotonin (34), dopamine (35), and γ-aminobutyric acid (GABA) (36). Serotonin (34), or 5-hydroxytryptamine, is synthesized from L-tryptophan (15) and requires the activities of tryptophan hydroxylase (EC 1.14.16.4) and the PLP-dependent enzyme DOPA (L-dihydroxyphenylalanine) decarboxylase [(common synonyms are: L-aromatic amino acid decarboxylase, tryptophan decarboxylase, 5-hydroxytryptophan decarboxylase; (EC 4.1.1.28)], which catalyzes the step from 5-hydroxy-L-tryptophan (37) to serotonin (34). The enzyme also catalyzes the biosynthesis of dopamine (35) from L-DOPA (38). Here the initial precursor is L-tyrosine (37), which is converted to L-DOPA (38) by the activity of L-tyrosine hydroxylase (EC 1.14.16.2) (Scheme 5A, B). GABA (36) in turn is synthesized by a decarboxylation reaction from L-glutamate (40) based on the activity of L-glutamate decarboxylase (Scheme 5C) (EC 4.1.1.15).
Serotonin (34) acts on the central nervous system where it affects a diverse range of conditions including appetite, sleep, or cognitive functions, and it is also well known for its ability to improve the overall mood [82]. In comparison, dopamine (35) affects the sympathetic nervous system where it is involved in the regulation of blood pressure and heart rate, while GABA (36) is a major inhibitory neurotransmitter in mammals that widely controls the excitability of neurons [83,84]. Consequently, low levels of vitB6 have been associated with depression and also dysfunction of the brain (e.g., epilepsy), and it is even considered by some authors as an ‘anti-stress’ agent [85,86,87,88]. In this context it is interesting to note that some plants, like Ginkgo biloba, synthesize derivatives of vitB6 that are suggested to inhibit the salvage pathway enzyme PDXK and thereby to impair neurotransmitter biosynthesis in the brain [8,10,89,90].
Scheme 5.
Synthesis of neurotransmitters in the brain. (A) Serotonin (34) is synthesized from L-tryptophan (15) via the intermediate 5-hydroxytryptophan (37). (B) Dopamine (35) is formed from L-tyrosine (39) via DOPA (38), and (C) GABA (36) is formed in a decarboxylation reaction from L-glutamate (40).
Scheme 5.
Synthesis of neurotransmitters in the brain. (A) Serotonin (34) is synthesized from L-tryptophan (15) via the intermediate 5-hydroxytryptophan (37). (B) Dopamine (35) is formed from L-tyrosine (39) via DOPA (38), and (C) GABA (36) is formed in a decarboxylation reaction from L-glutamate (40).

4.5. The antioxidative capacity of VitB6
Only recently was the potent antioxidant ability of vitB6 recognized. Here groundbreaking work from the group of Margaret Daub showed that the vitamin is highly efficient in quenching reactive oxygen species with a similar potential like described for carotenes and tocopherols [91,92,93]. Consequently, results from different organisms showed that reduced levels of the vitamin are connected with severe susceptibility to abiotic stress (oxidative, salt, drought, UV-B) [25,93,94,95]. Given the great consideration for other antioxidants like vitamins C and E or phenolics as ‘anti-aging’ compounds by the food industry and consumers, it will be interesting to see whether this relatively novel antioxidant will be embraced in similar ways in the future.
5. Conclusions
The biosynthesis of vitB6 and its function as a co-factor have been well resolved in the last years, leaving currently open what drives the activities of the different participating enzymes in the cell. Overall the latest studies indicate that vitB6 can be beneficial as a nutritional supplement, but can also be used as a pharmacological agent for disease treatment. Similarly, the diversity of PLP-dependent enzymes and the reactions they catalyze yield a wide range of targets for therapeutic approaches. However, the precise mechanisms of how vitB6 is beneficial are often still elusive, and to solidly define them is probably one of the most challenging tasks in the near future.
Acknowledgments
We would like to thank our Russian collaborator Cleatus for critical reading. We also would like to thank WSU for supporting this work.
- Samples Availability: Not applicable.
References and Notes
- Roje, S. Vitamin B biosynthesis in plants. Phytochemistry 2007, 68, 1904–1921. [Google Scholar] [CrossRef]
- Birch, T.W.; Gyorgy, P.; Harris, L.J. The vitamin B(2) complex. Differentiation of the antiblacktongue and the "P.-P." factors from lactoflavin and vitamin B(6) (so-called "rat pellagra" factor). Parts I-VI. Biochem. J. 1935, 29, 2830–2850. [Google Scholar]
- Percudani, R.; Peracchi, A. The B6 database: a tool for the description and classification of vitamin B6-dependent enzymatic activities and of the corresponding protein families. BMC Bioinformatics 2009, 10, 273. [Google Scholar] [CrossRef]
- Brouwer, D.A.; Welten, H.T.; van Doormaal, J.J.; Reijngoud, D.J.; Muskiet, F.A. Recommended dietary allowance of folic acid is insufficient for optimal homocysteine levels. Ned. Tijdschr. Geneeskd. 1998, 142, 782–786. [Google Scholar]
- Amadasi, A.; Bertoldi, M.; Contestabile, R.; Bettati, S.; Cellini, B.; di Salvo, M.L.; Borri-Voltattorni, C.; Bossa, F.; Mozzarelli, A. Pyridoxal 5'-phosphate enzymes as targets for therapeutic agents. Curr. Med. Chem. 2007, 14, 1291–1324. [Google Scholar] [CrossRef]
- Di Giovine, P. PLPMDB: pyridoxal-5'-phosphate dependent enzymes mutants database. Bioinformatics 2004, 20, 3652–3653. [Google Scholar] [CrossRef]
- Denesyuk, A.I.; Denessiouk, K.A.; Korpela, T.; Johnson, M.S. Functional attributes of the phosphate group binding cup of pyridoxal phosphate-dependent enzymes. J. Mol. Biol. 2002, 316, 155–172. [Google Scholar] [CrossRef]
- Mooney, S.; Leuendorf, J.E.; Hendrickson, C.; Hellmann, H. Vitamin B6: a long known compound of surprising complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Amrhein, N.; Kappes, B.; Macheroux, P.; Tews, I.; Raschle, T. Two independent routes of de novo vitamin B6 biosynthesis: not that different after all. Biochem. J. 2007, 407, 1–13. [Google Scholar] [CrossRef]
- Drewke, C.; Leistner, E. Biosynthesis of vitamin B6 and structurally related derivatives. Vitam. Horm. 2001, 61, 121–155. [Google Scholar] [CrossRef]
- Drewke, C.; Klein, M.; Clade, D.; Arenz, A.; Muller, R.; Leistner, E. 4-O-phosphoryl-L-threonine, a substrate of the pdxC(serC) gene product involved in vitamin B6 biosynthesis. FEBS Lett. 1996, 390, 179–182. [Google Scholar] [CrossRef]
- Notheis, C.; Drewke, C.; Leistner, E. Purification and characterization of the pyridoxol-5'-phosphate:oxygen oxidoreductase (deaminating) from Escherichia coli. Biochim. Biophys. Acta 1247, 265–271. [Google Scholar]
- Mittenhuber, G. Phylogenetic analyses and comparative genomics of vitamin B6 (pyridoxine) and pyridoxal phosphate biosynthesis pathways. J. Mol. Microbiol. Biotechnol. 2001, 3, 1–20. [Google Scholar]
- Tambasco-Studart, M.; Titiz, O.; Raschle, T.; Forster, G.; Amrhein, N.; Fitzpatrick, T.B. Vitamin B6 biosynthesis in higher plants. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 13687–13692. [Google Scholar]
- Burns, K.E.; Xiang, Y.; Kinsland, C.L.; McLafferty, F.W.; Begley, T.P. Reconstitution and biochemical characterization of a new pyridoxal-5'-phosphate biosynthetic pathway. J. Am. Chem. Soc. 2005, 127, 3682–3683. [Google Scholar]
- Tanaka, T.; Tateno, Y.; Gojobori, T. Evolution of vitamin B6 (pyridoxine) metabolism by gain and loss of genes. Mol. Biol. Evol. 2005, 22, 243–2450. [Google Scholar]
- Wagner, S.; Bernhardt, A.; Leuendorf, J. E.; Drewke, C.; Lytovchenko, A.; Mujahed, N.; Gurgui, C.; Frommer, W.B.; Leistner, E.; Fernie, A.R.; Hellmann, H. Analysis of the Arabidopsis rsr4-1/pdx1-3 mutant reveals the critical function of the PDX1 protein family in metabolism, development, and vitamin B6 biosynthesis. Plant Cell 2006, 18, 1722–1735. [Google Scholar] [CrossRef]
- Neuwirth, M.; Strohmeier, M.; Windeisen, V.; Wallner, S.; Deller, S.; Rippe, K.; Sinning, I.; Macheroux, P.; Tews, I. X-ray crystal structure of Saccharomyces cerevisiae Pdx1 provides insights into the oligomeric nature of PLP synthases. FEBS Lett. 2009, 583, 2179–2186. [Google Scholar] [CrossRef]
- Raschle, T.; Speziga, D.; Kress, W.; Moccand, C.; Gehrig, P.; Amrhein, N.; Weber-Ban, E.; Fitzpatrick, T.B. Intersubunit crosstalk in pyridoxal 5'-phosphate synthase, co-ordinated by the C-terminus of the synthase subunit. J. Biol. Chem. 2008, 284, 7706–7718. [Google Scholar]
- Gdynia, H.J.; Muller, T.; Sperfeld, A.D.; Kuhnlein, P.; Otto, M.; Kassubek, J.; Ludolph, A.C. Severe sensorimotor neuropathy after intake of highest dosages of vitamin B6. Neuromuscul. Disord. 2008, 18, 156–158. [Google Scholar] [CrossRef]
- Neuwirth, M.; Flicker, K.; Strohmeier, M.; Tews, I.; Macheroux, P. Thermodynamic characterization of the protein-protein interaction in the heteromeric Bacillus subtilis pyridoxalphosphate synthase. Biochemistry 2007, 46, 5131–5139. [Google Scholar] [CrossRef]
- Strohmeier, M.; Raschle, T.; Mazurkiewicz, J.; Rippe, K.; Sinning, I.; Fitzpatrick, T.B.; Tews, I. Structure of a bacterial pyridoxal 5'-phosphate synthase complex. Proc. Natl. Acad. Sci. USA 2006, 103, 19284–19289. [Google Scholar]
- Gengenbacher, M.; Fitzpatrick, T.B.; Raschle, T.; Flicker, K.; Sinning, I.; Muller, S.; Macheroux, P.; Tews, I.; Kappes, B. Vitamin B6 biosynthesis by the malaria parasite Plasmodium falciparum: biochemical and structural insights. J. Biol. Chem. 2006, 281, 3633–3641. [Google Scholar]
- Zein, F.; Zhang, Y.; Kang, Y.N.; Burns, K.; Begley, T.P.; Ealick, S.E. Structural insights into the mechanism of the PLP synthase holoenzyme from Thermotoga maritima. Biochemistry 2006, 45, 14609–14620. [Google Scholar]
- Gonzalez, E.; Danehower, D.; Daub, M.E. Vitamer levels, stress response, enzyme activity, and gene regulation of Arabidopsis lines mutant in the pyridoxine/pyridoxamine 5'-phosphate oxidase (PDX3) and the pyridoxal kinase (SOS4) genes involved in the vitamin B6 salvage pathway. Plant Physiol. 2007, 145, 985–996. [Google Scholar] [CrossRef]
- Ohdake, S. Isolation of "Oryzanin" (Antineuritic Vitamin) from Rice-polishings. Bull Agri. Chem. Soc. Jpn. 1932, 8, 11–46. [Google Scholar] [CrossRef]
- Oka, T. Vitamin B6. Nippon Rinsho 1999, 57, 2199–2204. [Google Scholar]
- Nogueira, A.; Duarte, A.F.; Magina, S.; Azevedo, F. Pellagra associated with esophageal carcinoma and alcoholism. Dermatol. Online J. 2009, 15, 8. [Google Scholar]
- Hendricks, W.M. Pellagra and pellagralike dermatoses: etiology, differential diagnosis, dermatopathology, and treatment. Semin. Dermatol. 1991, 10, 282–292. [Google Scholar]
- Morris, M.S.; Picciano, M.F.; Jacques, P.F.; Selhub, J. Plasma pyridoxal 5'-phosphate in the US population: the National Health and Nutrition Examination Survey, 2003–2004. Am. J. Clin. Nutr. 2008, 87, 1446–1454. [Google Scholar]
- Perry, T.; Holloway, H.W.; Weerasuriya, A.; Mouton, P.R.; Duffy, K.; Mattison, J.A.; Greig, N.H. Evidence of GLP-1-mediated neuroprotection in an animal model of pyridoxine-induced peripheral sensory neuropathy. Exp. Neurol. 2007, 203, 293–301. [Google Scholar] [CrossRef]
- Albin, R.L.; Albers, J.W.; Greenberg, H.S.; Townsend, J.B.; Lynn, R.B.; Burke, J.M. Jr.; Alessi, A.G. Acute sensory neuropathy-neuronopathy from pyridoxine overdose. Neurology 1987, 37, 1729–1732. [Google Scholar] [CrossRef]
- Lu, T.; Xu, Y.; Monttinen, E.S.; Kato, N. Supplementing vitamin B6 to a low vitamin B6 diet exaggerates UVB-induced skin tumorigenesis in DMBA-treated hairless mice. J. Nutr. Sci. Vitaminol. (Tokyo) 2008, 54, 262–265. [Google Scholar]
- Wondrak, G.T.; Roberts, M.J.; Jacobson, M.K.; Jacobson, E.L. 3-hydroxypyridine chromophores are endogenous sensitizers of photooxidative stress in human skin cells. J. Biol. Chem. 2004, 279, 30009–30020. [Google Scholar]
- Maeda, T.; Taguchi, H.; Minami, H.; Sato, K.; Shiga, T.; Kosaka, H.; Yoshikawa, K. Vitamin B6 phototoxicity induced by UVA radiation. Arch. Dermatol. Res. 2000, 292, 562–567. [Google Scholar] [CrossRef]
- Muhia, D.K.; Swales, C.A.; Deng, W.; Kelly, J.M.; Baker, D.A. The gametocyte-activating factor xanthurenic acid stimulates an increase in membrane-associated guanylyl cyclase activity in the human malaria parasite Plasmodium falciparum. Mol. Microbiol. 2001, 42, 553–560. [Google Scholar] [CrossRef]
- Rossi, F.; Garavaglia, S.; Giovenzana, G.B.; Arca, B.; Li, J.; Rizzi, M. Crystal structure of the Anopheles gambiae 3-hydroxykynurenine transaminase. Proc. Natl. Acad. Sci. USA 2006, 103, 5711–576. [Google Scholar]
- Rossi, F.; Lombardo, F.; Paglino, A.; Cassani, C.; Miglio, G.; Arca, B.; Rizzi, M. Identification and biochemical characterization of the Anopheles gambiae 3-hydroxykynurenine transaminase. Febs.J. 2005, 272, 5653–5662. [Google Scholar] [CrossRef]
- Marti-Carvajal, A.J.; Sola, I.; Lathyris, D.; Salanti, G. Homocysteine lowering interventions for preventing cardiovascular events. Cochrane Database Syst. Rev. 2009, CD006612. [Google Scholar]
- Muller, I.B.; Wu, F.; Bergmann, B.; Knockel, J.; Walter, R.D.; Gehring, H.; Wrenger, C. Poisoning pyridoxal 5-phosphate-dependent enzymes: a new strategy to target the malaria parasite Plasmodium falciparum. PLoS One 2009, 4, 4406. [Google Scholar]
- Wrenger, C.; Eschbach, M.L.; Muller, I.B.; Warnecke, D.; Walter, R.D. Analysis of the vitamin B6 biosynthesis pathway in the human malaria parasite Plasmodium falciparum. J. Biol. Chem. 2005, 280, 5242–5248. [Google Scholar]
- Pegg, A.E.; McGovern, K.A.; Wiest, L. Decarboxylation of alpha-difluoromethylornithine by ornithine decarboxylase. Biochem. J. 1987, 241, 305–307. [Google Scholar]
- Poulin, R.; Lu, L.; Ackermann, B.; Bey, P.; Pegg, A.E. Mechanism of the irreversible inactivation of mouse ornithine decarboxylase by alpha-difluoromethylornithine. Characterization of sequences at the inhibitor and coenzyme binding sites. J. Biol. Chem. 1992, 267, 15015–15018. [Google Scholar]
- Kern, A.D.; Oliveira, M.A.; Coffino, P.; Hackert, M.L. Structure of mammalian ornithine decarboxylase at 1.6 A resolution: stereochemical implications of PLP-dependent amino acid decarboxylases. Structure 1999, 7, 567–581. [Google Scholar]
- Kernebeck, T.; Lohse, A.W.; Grotzinger, J. A bioinformatical approach suggests the function of the autoimmune hepatitis target antigen soluble liver antigen/liver pancreas. Hepatology 2001, 34, 230–233. [Google Scholar]
- Krauth-Siegel, L.R.; Comini, M.A.; Schlecker, T. The trypanothione system. Subcell. Biochem. 2007, 44, 231–251. [Google Scholar] [CrossRef]
- Fairlamb, A.H.; Blackburn, P.; Ulrich, P.; Chait, B.T.; Cerami, A. Trypanothione: a novel bis(glutathionyl)spermidine cofactor for glutathione reductase in trypanosomatids. Science 1985, 227, 1485–1487. [Google Scholar]
- Snell, K.; Natsumeda, Y.; Eble, J.N.; Glover, J.L.; Weber, G. Enzymic imbalance in serine metabolism in human colon carcinoma and rat sarcoma. Br. J. Cancer 1988, 57, 87–90. [Google Scholar] [CrossRef]
- Snell, K.; Natsumeda, Y.; Weber, G. The modulation of serine metabolism in hepatoma 3924A during different phases of cellular proliferation in culture. Biochem. J. 1987, 245, 609–612. [Google Scholar]
- Chang, S.J. Vitamin B6 antagonists alter the function and ultrastructure of mice endothelial cells. J. Nutr. Sci. Vitaminol. (Tokyo) 2000, 46, 149–153. [Google Scholar]
- Renwick, S.B.; Skelly, J.V.; Chave, K.J.; Sanders, P.G.; Snell, K.; Baumann, U. Purification, crystallization and preliminary X-ray analysis of human recombinant cytosolic serine hydroxymethyltransferase. Acta Crystallogr. D. Biol. Crystallogr. 1998, 54, 1030–1031. [Google Scholar] [CrossRef]
- Pires, A.R.; Araujo, A.N.; Montenegro, M.C.; Chocholous, P.; Solich, P. New ionophores for vitamin B1 and vitamin B6 potentiometric sensors for multivitaminic control. J. Pharm. Biomed. Anal. 2008, 46, 683–691. [Google Scholar] [CrossRef]
- Ishihara, J.; Iso, H.; Inoue, M.; Iwasaki, M.; Okada, K.; Kita, Y.; Kokubo, Y.; Okayama, A.; Tsugane, S. Intake of folate, vitamin B6 and vitamin B12 and the risk of CHD: the Japan Public Health Center-Based Prospective Study Cohort I. J. Am. Coll. Nutr. 2008, 27, 127–136. [Google Scholar]
- Czeizel, A.E.; Puho, E.; Banhidy, F.; Acs, N. Oral pyridoxine during pregnancy : potential protective effect for cardiovascular malformations. Drugs R. D. 2004, 5, 259–269. [Google Scholar] [CrossRef]
- Merrill, R.M.; Taylor, P.; Aldana, S.G. Coronary Health Improvement Project (CHIP) is associated with improved nutrient intake and decreased depression. Nutrition 2008, 24, 314–321. [Google Scholar] [CrossRef]
- Booth, G.L.; Wang, E.E. Preventive health care, 2000 update: screening and management of hyperhomocysteinemia for the prevention of coronary artery disease events. The Canadian Task Force on Preventive Health Care. CMAJ 2000, 163, 21–29. [Google Scholar]
- Nozaki, T.; Shigeta, Y.; Saito-Nakano, Y.; Imada, M.; Kruger, W.D. Characterization of transsulfuration and cysteine biosynthetic pathways in the protozoan hemoflagellate, Trypanosoma cruzi. Isolation and molecular characterization of cystathionine beta-synthase and serine acetyltransferase from Trypanosoma. J. Biol. Chem. 2001, 276, 6516–6523. [Google Scholar]
- Graham, I.M.; Daly, L.E.; Refsum, H.M.; Robinson, K.; Brattstrom, L.E.; Ueland, P.M.; Palma-Reis, R.J.; Boers, G.H.; Sheahan, R.G.; Israelsson, B.; Uiterwaal, C.S.; Meleady, R.; McMaster, D.; Verhoef, P.; Witteman, J.; Rubba, P.; Bellet, H.; Wautrecht, J.C.; de Valk, H.W.; Sales Luis, A.C.; Parrot-Rouland, F.M.; Tan, K.S.; Higgins, I.; Garcon, D.; Andria, G. Plasma homocysteine as a risk factor for vascular disease. The European Concerted Action Project. JAMA 1997, 277, 1775–1781. [Google Scholar]
- Lal, K.J.; Dakshinamurti, K. The relationship between low-calcium-induced increase in systolic blood pressure and vitamin B6. J. Hypertens. 1995, 13, 327–332. [Google Scholar]
- Lal, K.J.; Dakshinamurti, K.; Thliveris, J. The effect of vitamin B6 on the systolic blood pressure of rats in various animal models of hypertension. J. Hypertens. 1996, 14, 355–363. [Google Scholar] [CrossRef]
- Dakshinamurti, K.; Lal, K.J.; Ganguly, P.K. Hypertension, calcium channel and pyridoxine (vitamin B6). Mol. Cell Biochem. 1998, 188, 137–148. [Google Scholar] [CrossRef]
- Aybak, M.; Sermet, A.; Ayyildiz, M.O.; Karakilcik, A.Z. Effect of oral pyridoxine hydrochloride supplementation on arterial blood pressure in patients with essential hypertension. Arzneimittelforschung 1995, 45, 1271–1273. [Google Scholar]
- Dakshinamurti, K.; Dakshinamurti, S. Blood pressure regulation and micronutrients. Nutr. Res. Rev. 2001, 14, 3–44. [Google Scholar] [CrossRef]
- Okuda, T.; Sumiya, T.; Iwai, N.; Miyata, T. Pyridoxine 5'-phosphate oxidase is a candidate gene responsible for hypertension in Dahl-S rats. Biochem. Biophys. Res. Commun. 2004, 313, 647–53. [Google Scholar] [CrossRef]
- Lieber, C.S. Mechanism of ethanol induced hepatic injury. Pharmacol. Ther. 1990, 46, 1–41. [Google Scholar] [CrossRef]
- Vasdev, S.; Whalen, M.; Ford, C. A.; Longerich, L.; Prabhakaran, V.; Parai, S. Ethanol- and threonine-induced hypertension in rats: a common mechanism. Can. J. Cardiol. 1995, 11, 807–815. [Google Scholar]
- Vasdev, S.; Wadhawan, S.; Ford, C.A.; Parai, S.; Longerich, L.; Gadag, V. Dietary vitamin B6 supplementation prevents ethanol-induced hypertension in rats. Nutr. Metab. Cardiovasc. Dis. 1999, 9, 55–63. [Google Scholar]
- Gloria, L.; Cravo, M.; Camilo, M.E.; Resende, M.; Cardoso, J.N.; Oliveira, A.G.; Leitao, C.N.; Mira, F.C. Nutritional deficiencies in chronic alcoholics: relation to dietary intake and alcohol consumption. Am. J. Gastroenterol. 1997, 92, 485–489. [Google Scholar]
- Huber, K.H.; Rexroth, W.; Werle, E.; Koeth, T.; Weicker, H.; Hild, R. Sympathetic neuronal activity in diabetic and non-diabetic subjects with peripheral arterial occlusive disease. Klin. Wochenschr. 1991, 69, 233–238. [Google Scholar] [CrossRef]
- Cicila, G.T. Strategy for uncovering complex determinants of hypertension using animal models. Curr. Hypertens. Rep. 2000, 2, 217–226. [Google Scholar] [CrossRef]
- MacKenzie, K.E.; Wiltshire, E.J.; Gent, R.; Hirte, C.; Piotto, L.; Couper, J.J. Folate and vitamin B6 rapidly normalize endothelial dysfunction in children with type 1 diabetes mellitus. Pediatrics 2006, 118, 242–253. [Google Scholar] [CrossRef]
- Nakamura, S.; Li, H.; Adijiang, A.; Pischetsrieder, M.; Niwa, T. Pyridoxal phosphate prevents progression of diabetic nephropathy. Nephrol. Dial. Transplant 2007, 22, 2165–2174. [Google Scholar] [CrossRef]
- Adrover, M.; Vilanova, B.; Munoz, F.; Donoso, J. Inhibition of glycosylation processes: the reaction between pyridoxamine and glucose. Chem. Biodivers. 2005, 2, 964–975. [Google Scholar] [CrossRef]
- Nagore, E.; Insa, A.; Sanmartin, O. Antineoplastic therapy-induced palmar plantar erythrodysesthesia ('hand-foot') syndrome. Incidence, recognition and management. Am. J. Clin. Dermatol. 2000, 1, 225–234. [Google Scholar]
- Taylor, B.V.; Oudit, G.Y.; Evans, M. Homocysteine, vitamins, and coronary artery disease. Comprehensive review of the literature. Can. Fam. Physician 2000, 46, 2236–2245. [Google Scholar]
- Matsubara, K.; Matsumoto, H.; Mizushina, Y.; Lee, J.S.; Kato, N. Inhibitory effect of pyridoxal 5'-phosphate on endothelial cell proliferation, replicative DNA polymerase and DNA topoisomerase. Int. J. Mol. Med. 2003, 12, 51–55. [Google Scholar]
- Bostom, A.G.; Carpenter, M.A.; Kusek, J.W.; Hunsicker, L.G.; Pfeffer, M.A.; Levey, A.S.; Jacques, P.F.; McKenney, J. Rationale and design of the Folic Acid for Vascular Outcome Reduction In Transplantation (FAVORIT) trial. Am. Heart J. 2006, 152, 448. [Google Scholar]
- Menon, V.; Wang, X.; Greene, T.; Beck, G.J.; Kusek, J.W.; Selhub, J.; Levey, A.S.; Sarnak, M.J. Homocysteine in chronic kidney disease: Effect of low protein diet and repletion with B vitamins. Kidney Int. 2005, 67, 1539–1546. [Google Scholar] [CrossRef]
- Adrover, M.; Vilanova, B.; Frau, J.; Munoz, F.; Donoso, J. A comparative study of the chemical reactivity of pyridoxamine, Ac-Phe-Lys and Ac-Cys with various glycating carbonyl compounds. Amino Acids 2009, 36, 437–448. [Google Scholar] [CrossRef]
- Metz, T.O.; Alderson, N.L.; Thorpe, S.R.; Baynes, J.W. Pyridoxamine, an inhibitor of advanced glycation and lipoxidation reactions: a novel therapy for treatment of diabetic complications. Arch. Biochem. Biophys. 2003, 419, 41–49. [Google Scholar] [CrossRef]
- Voziyan, P.A.; Metz, T.O.; Baynes, J.W.; Hudson, B.G. A post-Amadori inhibitor pyridoxamine also inhibits chemical modification of proteins by scavenging carbonyl intermediates of carbohydrate and lipid degradation. J. Biol. Chem. 2002, 277, 3397–3403. [Google Scholar]
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–66. [Google Scholar] [CrossRef]
- Matxain, J.M.; Padro, D.; Ristila, M.; Strid, A.; Eriksson, L.A. Evidence of high *OH radical quenching efficiency by vitamin B6. J. Phys. Chem. B. 2009, 113, 9629–9632. [Google Scholar] [CrossRef]
- Belelli, D.; Harrison, N.L.; Maguire, J.; Macdonald, R.L.; Walker, M.C.; Cope, D.W. Extrasynaptic GABAA receptors: form, pharmacology, and function. J. Neurosci. 2009, 29, 12757–12763. [Google Scholar]
- Hvas, A.M.; Juul, S.; Bech, P.; Nexo, E. Vitamin B6 level is associated with symptoms of depression. Psychother. Psychosom. 2004, 73, 340–343. [Google Scholar] [CrossRef]
- Hoffmann, G.F.; Schmitt, B.; Windfuhr, M.; Wagner, N.; Strehl, H.; Bagci, S.; Franz, A. R.; Mills, P.B.; Clayton, P.T.; Baumgartner, M.R.; Steinmann, B.; Bast, T.; Wolf, N.I.; Zschocke, J. Pyridoxal 5'-phosphate may be curative in early-onset epileptic encephalopathy. J. Inherit. Metab. Dis. 2007, 30, 96–99. [Google Scholar] [CrossRef]
- Lott, I.T.; Coulombe, T.; Di Paolo, R.V.; Richardson, E.P. Jr.; Levy, H.L. Vitamin B6-dependent seizures: pathology and chemical findings in brain. Neurology 1978, 28, 47–54. [Google Scholar] [CrossRef]
- McCarty, M.F. High-dose pyridoxine as an 'anti-stress' strategy. Med. Hypotheses 2000, 54, 803–807. [Google Scholar] [CrossRef]
- Leuendorf, J.E.; Genau, A.; Szewczyk, A.; Mooney, S.; Drewke, C.; Leistner, E.; Hellmann, H. The Pdx1 family is structurally and functionally conserved between Arabidopsis thaliana and Ginkgo biloba. FEBS J. 2008, 275, 960–969. [Google Scholar]
- Kästner, U.; Hallmen, C.; Wiese, M.; Leistner, E.; Drewke, C. The human pyridoxal kinase, a plausible target for ginkgotoxin from Ginkgo biloba. FEBS J. 2007, 274, 1036–1045. [Google Scholar] [CrossRef]
- Denslow, S.A.; Walls, A.A.; Daub, M.E. Regulation of biosynthetic genes and antioxidant properties of vitamin B6 vitamers during plant defense responses. Physiol. Mol. Plant Path. 2005, 66, 244–255. [Google Scholar] [CrossRef]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Jenns, A.E.; Chung, K.R.; Daub, M.E. SOR1, a gene required for photosensitizer and singlet oxygen resistance in Cercospora fungi, is highly conserved in divergent organisms. Mol. Cell 1998, 1, 603–609. [Google Scholar] [CrossRef]
- Chen, H.; Xiong, L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005, 44, 396–408. [Google Scholar] [CrossRef]
- Chumnantana, R.; Yokochi, N.; Yagi, T. Vitamin B6 compounds prevent the death of yeast cells due to menadione, a reactive oxygen generator. Biochim. Biophys. Acta 2005, 1722, 84–91. [Google Scholar]
- DiSorbo, D.M.; Wagner, R. Jr.; Nathanson, L. In vivo and in vitro inhibition of B16 melanoma growth by vitamin B6. Nutr. Cancer 1985, 7, 43–52. [Google Scholar] [CrossRef]
- DiSorbo, D.M.; Nathanson, L. High-dose pyridoxal supplemented culture medium inhibits the growth of a human malignant melanoma cell line. Nutr. Cancer 1983, 5, 10–15. [Google Scholar] [CrossRef]
- Komatsu, S.; Yanaka, N.; Matsubara, K.; Kato, N. Antitumor effect of vitamin B6 and its mechanisms. Biochim. Biophys. Acta 2003, 1647, 127–130. [Google Scholar]
- Stolzenberg-Solomon, R.Z.; Albanes, D.; Nieto, F.J.; Hartman, T.J.; Tangrea, J.A.; Rautalahti, M.; Sehlub, J.; Virtamo, J.; Taylor, P.R. Pancreatic cancer risk and nutrition-related methyl-group availability indicators in male smokers. J. Natl. Cancer Inst. 1999, 91, 535–541. [Google Scholar] [CrossRef]
- Harbige, L.S. Nutrition and immunity with emphasis on infection and autoimmune disease. Nutr. Health 1996, 10, 285–312. [Google Scholar] [CrossRef]
- Delport, R.; Ubbink, J.B.; Bosman, H.; Bissbort, S.; Vermaak, W.J. Altered vitamin B6 homeostasis during aminophylline infusion in the beagle dog. Int. J. Vitam. Nutr. Res. 1990, 60, 35–40. [Google Scholar]
- Delport, R.; Ubbink, J.B.; Serfontein, W.J.; Becker, P.J.; Walters, L. Vitamin B6 nutritional status in asthma: the effect of theophylline therapy on plasma pyridoxal-5'-phosphate and pyridoxal levels. Int. J. Vitam. Nutr. Res. 1988, 58, 67–72. [Google Scholar]
- van den Berg, H.; Louwerse, E.S.; Bruinse, H.W.; Thissen, J.T.; Schrijver, J. Vitamin B6 status of women suffering from premenstrual syndrome. Hum. Nutr. Clin. Nutr. 1986, 40, 441–450. [Google Scholar]
- Bernstein, A.L. Vitamin B6 in clinical neurology. Ann. N. Y. Acad. Sci. 1990, 585, 250–260. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Hellmann, H.; Mooney, S. Vitamin B6: A Molecule for Human Health? Molecules 2010, 15, 442-459. https://doi.org/10.3390/molecules15010442
AMA Style
Hellmann H, Mooney S. Vitamin B6: A Molecule for Human Health? Molecules. 2010; 15(1):442-459. https://doi.org/10.3390/molecules15010442
Chicago/Turabian StyleHellmann, Hanjo, and Sutton Mooney. 2010. "Vitamin B6: A Molecule for Human Health?" Molecules 15, no. 1: 442-459. https://doi.org/10.3390/molecules15010442