Arginine as a Synergistic Virucidal Agent

Abstract
:1. Introduction
2. Summary of Virucidal Activities of Arginine
2.1. In vitro virus inactivation



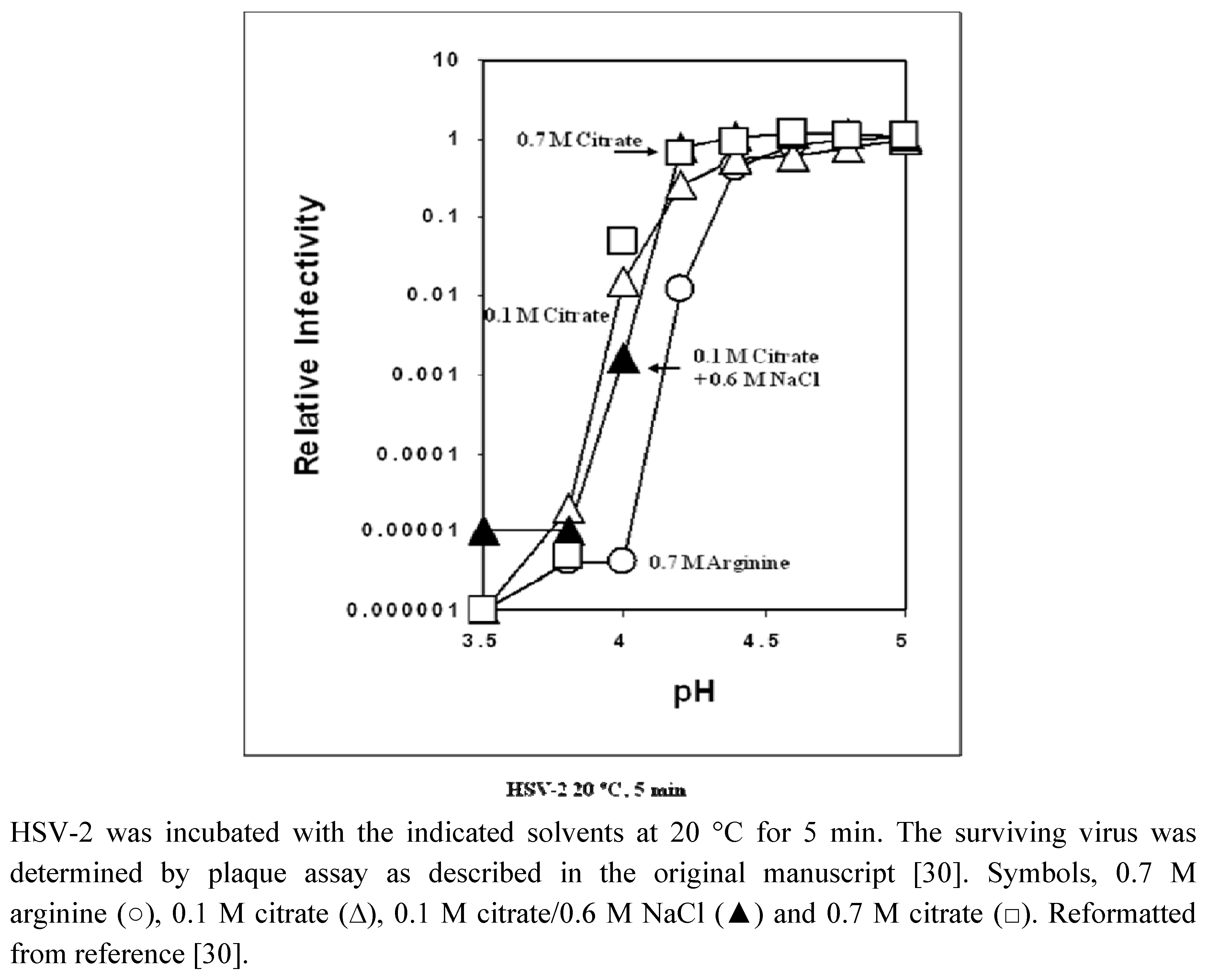{kind=link}
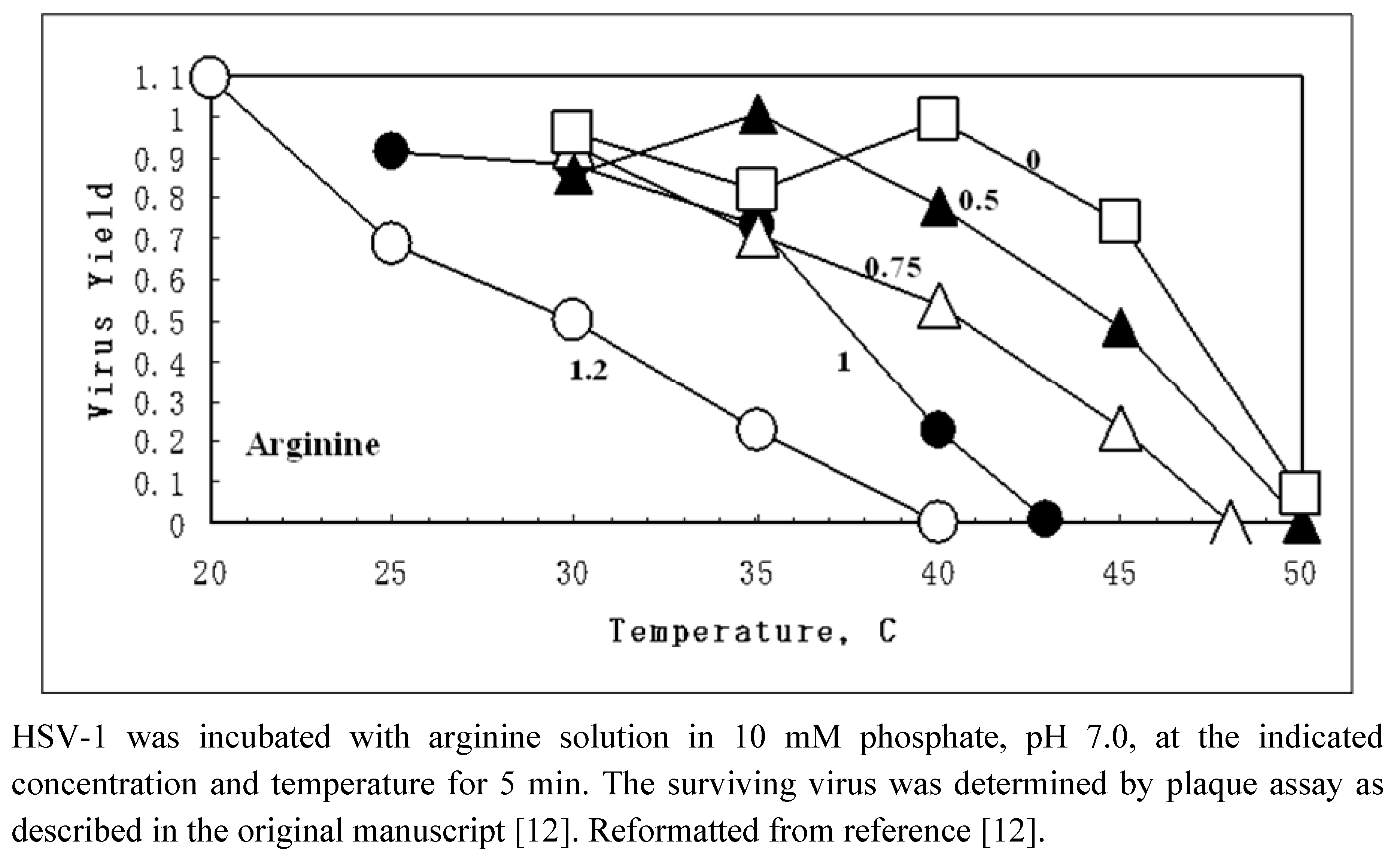{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | Temperature at 50% inactivation | Temperature at 90% inactivation | ||
|---|---|---|---|---|
| 0 (PBS) | 47 | 50 | ||
| Arginine | NaCl | Arginine | NaCl | |
| 0.5 M | -2 | 0 | -3 | |
| 0.75 M | -7 | -4 | -5 | -2 |
| 1.0 M | -10 | -6 | -10 | |
| 1.2 M | -17 | -7 | -16 | -4 |
2.2. In vivo virus inactivation
3. Mechanism of Virus Inactivation by Physical and Chemical Stresses



3.1. Mechanism-1
3.2. Mechanism-2
3.3. Mechanism-3
4. Mechanism of the Virus Inactivation Effects of Arginine
| Arginine Concentration / M | Lysozyme/°C | RNase/°C |
|---|---|---|
| 0.1 | 0 | 0 |
| 0.2 | -1 | |
| 0.5 | -1 | -1 |
| 1.0 | -1 | -3 |
| 2.0 | 0 | -3 |

5. Conclusions
References
- Ellebedy, A.H.; Webby, R.J. Influenza vaccines. Vaccine 2009, 27, D65–D68. [Google Scholar] [CrossRef]
- Toplak, N.; Avcin, T. Influenza and autoimmunity. Ann. N. Y. Acad. Sci. 2009, 1173, 619–626. [Google Scholar] [CrossRef]
- Mathews, J.D.; Chesson, J.M.; McCaw, J.M.; McVerson, J. Understanding influenza transmission, immunity and pandemic threats. Influenza Other Respir. Viruses 2009, 3, 143–149. [Google Scholar] [CrossRef]
- Wanaratana, S.; Tantilertcharoen, R.; Sasipreeyajan, J.; Pakpinyo, S. The inactivation of avian virus subtype H5N1 isolated from chickens in Thailand by chemical and physical treatments. Vet. Microbiol. 2009, 140, 43–48. [Google Scholar]
- Patnayak, D.P.; Prasad, A.M.; Malik, Y.S.; Ramakrishnan, M.A.; Goyal, S.M. Efficacy of disinfectants and hand sanitizers against respiratory viruses. Avian Dis. 2008, 52, 199–202. [Google Scholar] [CrossRef]
- Grayson, M.L.; Melvani, S.; Druce, J.; Barr, I.G.; Ballard, S.A.; Johnson, P.D.; Mastorakos, T.; Birch, C. Efficacy of soap and water and alcohol-based hand-rub preparations against live H1N1 influenza virus on the hands of human volunteers. Clin. Infect. Dis. 2009, 48, 285–291. [Google Scholar] [CrossRef]
- Lombardi, M.E.; Ladman, B.S.; Alphin, R.L.; Benson, E.R. Inactivation of avian influenza virus using common detergents and chemicals. Avian Dis. 2008, 52, 118–123. [Google Scholar] [CrossRef]
- Alphin, R.L.; Johnson, K.J.; Ladman, B.S.; Benson, E.R. Inactivation of avian influenza virus using four common chemicals and one detergent. Poult. Sci. 2009, 88, 1181–1185. [Google Scholar] [CrossRef]
- Rennie, O.; Bowtell, P.; Hull, D.; Charbonneau, D.; Lambkin-Williams, R.; Oxford, J. Low pH gel intranasal sprays inactivate influenza viruses in vitro and protect ferrets against influenza infection. Respir. Res. 2007, 8, 38. [Google Scholar] [CrossRef]
- Yamasaki, H.; Tsujimoto, K.; Koyama, A.H.; Ejima, D.; Arakawa, T. Arginine facilitates inactivation of enveloped viruses. J. Pharm. Sci. 2008, 97, 3063–3073. [Google Scholar]
- Katsuyama, Y.; Yamasaki, H.; Tsujimoto, K.; Koyama, A.H.; Ejima, D.; Arakawa, T. Butyroyl-arginine as a potent virus inactivation agent. Int. J. Pharm. 2008, 361, 92–98. [Google Scholar] [CrossRef]
- Utsunimoya, H.; Ichinose, M.; Tsujimoto, K.; Katsuyama, Y.; Yamasaki, H.; Koyama, A.H.; Ejima, D.; Arakawa, T. Co-operative thermal inactivation of herpes simplex virus and influenza virus by arginine ans NaCl. Int. J. Pharm. 2009, 366, 99–102. [Google Scholar] [CrossRef]
- Arakawa, T.; Kita, Y.; Koyama, A.H. Synergistic virus inactivation effects of arginine. Biotechnol. J. 2009, 4, 174–178. [Google Scholar]
- Arakawa, T.; Tsumoto, K. The effects of argnine on refolding of aggregated proteins: not facilitate refolding, but suppress aggregation. Biochem. Biophys. Res. Commun. 2003, 304, 148–152. [Google Scholar] [CrossRef]
- Tsumoto, K.; Ejima, D.; Kita, Y.; Arakawa, T. Review: why is arginine effective in suppressing aggregation? Protein Pept. Lett. 2005, 12, 613–619. [Google Scholar] [CrossRef]
- Arakawa, T.; Kita, Y.; Tsumoto, K.; Fukada, H.; Ejima, D. Aggregation suppression of proteins by arginine during thermal unfolding. Protein Pept. Lett. 2006, 13, 921–927. [Google Scholar] [CrossRef]
- Shiraki, K.; Kudou, M.; Fujiwara, S.; Imanaka, T.; Takagi, M. Biophysical effect of amino acids on the prevention of protein aggregation. J. Biochem. 2002, 132, 591–595. [Google Scholar] [CrossRef]
- Tsumoto, K.; Umetsu, M.; Kumaga, I.; Ejima, D.; Arakawa, T. Solubilization of active green fluorescent protein from insoluble particles by guanidine and arginine. Biochem. Biophys. Res. Commun. 2003, 312, 1382–1386. [Google Scholar]
- Sakai, N.; Takeuchi, T.; Futaki, S.; Matile, S. Direct observation of anion-mediated translocation of fluorescent oligoarginine carriers into and across liquid and anionic bilayer membranes. ChemBioChem 2005, 6, 114–122. [Google Scholar] [CrossRef]
- Hirai, A.; Kawasaki, H.; Tanaka, S.; Nemoto, N.; Suzuki, M.; Maeda, H. Effects of L-arginine on aggregate of fatty-acid/potassium soap in the aqueous media. Colloid. Polym. Sci. 2006, 284, 520–528. [Google Scholar] [CrossRef]
- Arakawa, T.; Philo, J.S.; Tsumoto, K.; Yumioka, R.; Ejima, D. Elution of antibodies from a Protein-A column by aqueous arginine solution. Protein Expression Purif. 2004, 36, 244–248. [Google Scholar] [CrossRef]
- Ejima, D.; Yumioka, R.; Tsumoto, K.; Arakawa, T. Effective elution of antibodies by arginine and arginine derivatives in affinity colum chromatography. Anal. Biochem. 2005, 345, 250–257. [Google Scholar]
- Tsumoto, K.; Ejima, D.; Nagase, K.; Arakawa, T. Arginine improves protein elution in hydrophobic interaction chromatography. The cases of human interleukin-6 and activin-A. J. Chromatogr. A 2007, 1154, 81–86. [Google Scholar] [CrossRef]
- Ejima, D.; Yumioka, R.; Arakawa, T.; Tsumoto, K. Arginine as an effective additive in gel permeation chromatography. J. Chromatogr. A 2005, 1094, 49–55. [Google Scholar] [CrossRef]
- Arakawa, T.; Tsumoto, K.; Nagase, K.; Ejima, D. The effects of arginine on protein binding and elution in hydrophobic interaction and ion-exchange chromatography. Protein Expr. Purif. 2007, 54, 110–116. [Google Scholar]
- Abe, R.; Kudou, M.; Tanaka, Y.; Arakawa, T.; Tsumoto, K. Immobilized metal affinity chromatography in the presence of arginine. Biochem. Biophys. Res. Commun. 2009, 381, 306–310. [Google Scholar] [CrossRef]
- Futatsumori-Sugai, M.; Abe, R.; Watanabe, M.; Kudou, M.; Yamamoto, T.; Ejima, D.; Arakawa, T.; Tsumoto, K. Utilization of Arg-elution method for FLAG-tag based chromatography. Protein Expr. Purif. 2009, 67, 148–155. [Google Scholar] [CrossRef]
- Brorson, K.; Norling, L. Current and future approaches to ensure the viral safety of biopharmaceuticals. Dev. Biol. 2004, 118, 17–29. [Google Scholar]
- Yap, P.L. The viral safety of intravenous immune globulin. Clin. Exp. Immunol. 1996, 104, 35–42. [Google Scholar]
- Tsujimoto, K.; Uozaki, M.; Yamazaki, H.; Utsunomiya, H.; Ichinose, M.; Koyama, A.H.; Arakawa, T. Solvent-induced virus inactivation by acidic arginine solution. Int. J. Mol. Med. 2010, 25, 433–437. [Google Scholar]
- Ikeda, K.; Yamasaki, H.; Minami, S.; Naito, T.; Irie, H.; Arakawa, T.; Koyama, A.H. Virucidal ability of arginine and its possible application as an antiherpetic agent. In From the Hallowed Halls of Herpesvirology; Baines, J., Blaho, J., Eds.; Imperial College Press: London, UK, 2009; in press. [Google Scholar]
- Kissmann, J.; Ausar, S.F.; Rudolph, A.; Braun, C.; Cape, S.P.; Sievers, R.E.; Federspiel, M.J.; Joshi, S.B.; Middaugh, C.R. Stabilization of measles virus for vaccine formulation. Human Vaccines 2008, 4, 350–359. [Google Scholar] [CrossRef]
- Ausar, S.F.; Rexroad, J.; Frolov, V.G.; Look, J.L.; Konar, N.; Middaugh, C.R. Analysis of the thermal and pH stability of human respiratory syncytial virus. Molecular Pharma. 2005, 2, 491–499. [Google Scholar] [CrossRef]
- Rexroad, J.; Martin, T.T.; McNeilly, D.; Godwin, S.; Middaugh, C.R. Thermal stability of adenovirus 2 as a function of pH. J. Pharm. Sci. 2006, 95, 1469–1479. [Google Scholar] [CrossRef]
- Rexroad, J.; Evans, R.K.; Middaugh, C.R. Effect of pH and ionic strength on the physical stability of adenovirus type 5. J. Pharm. Sci. 2006, 95, 237–247. [Google Scholar] [CrossRef]
- Ausar, S.F.; Foubert, T.R.; Hudson, M.H.; Vedvick, T.S.; Middaugh, C.R. Conformational stability and disassembly of Norwalk virus like particles: effect of pH and temperature. J. Biol. Chem. 2006, 281, 19478–19488. [Google Scholar]
- Thomas, A.A.M.; Brioen, P.; Boeye, A. A monoclonal antibody that neutralizes poliovirus by cross-linking virions. J. Virol. 1985, 54, 7–13. [Google Scholar]
- Daher, K.A.; Selsted, M.E.; Lehrer, R. Direct inactivation of viruses by human granulocyte defensins. J. Virol. 1986, 60, 1068–1074. [Google Scholar]
- Wimley, W.C.; Selsted, M.E.; White, S.H. Interactions between human defensins and lipid bilayers: evidence for formation of multimeric pores. Protein Sci. 1994, 3, 1362–1373. [Google Scholar] [CrossRef]
- Peetermans, J.; Colinet, G.; Stephenne, J.; Bouillet, A. Stability of freeze-dried and reconstituted measles vaccines. Dev. Biol. Stand. 1978, 41, 259–264. [Google Scholar]
- Ohtake, S.; Martin, R.A.; Yee, L.; Chen, D.; Kristensen, D.D.; Lechuga-Ballesteros, D.; Truong-Le, V. Heat-stable measles vaccine produced by spray drying. Vaccine. In Press.
- Ver, B.A.; Melnick, J.L.; Wallis, C. Efficient filtration and sizing of viruses with membrane filters. J. Virol. 1968, 2, 21–25. [Google Scholar]
- Thomas, A.A.M.; Vrijsen, R.; Boeye, A. Relationship between poliovirus neutralization and aggregation. J. Virol. 1986, 59, 479–485. [Google Scholar]
- Wallis, C.; Melnick, J.L. Virus aggregation as the cause of the non-neutralizable persisted fraction. J. Virol. 1967, 1, 478–488. [Google Scholar]
- Pinto, F.; Maillard, J.Y.; Denyer, S.P.; McGeechan, P. Polyhexamethylene biguanide exposure leads to viral aggregation. J. Appl. Microbiol. In Press.
- Kim, Y.C.; Quan, F.S.; Compans, R.W.; Kang, S.M.; Prausnitz, M.R. Formulation and coating of microneedles with inactivated influenza virus to improve vaccine stability and immunogenicity. J. Control. Release 2010, 142, 187–195. [Google Scholar] [CrossRef]
- Michalsky, R.; Pfromm, P.H.; Czermak, P.; Sorensen, C.M.; Passarelli, A.L. Effects of temperature and shear force on infectivity of the baculovirus Autographa californica M nucleopolyhechovirus. J. Virol. Methods 2008, 153, 90–96. [Google Scholar] [CrossRef]
- Melnick, J.L. Preservation of viruses by freezing. Federation Proc. 1965, 15, 280–283. [Google Scholar]
- Watson, D.H.; Wildy, O. Some serological properties of herpesvirus particles studied by the electron microscope. Virology 1963, 21, 100–110. [Google Scholar] [CrossRef]
- Merryman, H.T. Mechanics of freezing in living cells and tissues. Science 1956, 124, 515–521. [Google Scholar]
- Wallis, C.; Melnick, J.L. Stabilization of enveloped virus by dimethyl sulfoxide. J. Virol. 1968, 2, 953–954. [Google Scholar]
- Lebon, P.; Protat, A.; Molinie, P. L-arginine elution of measles virus adsorbed on monkey erythrocytes. Infect. Immunity 1975, 11, 1407–1408. [Google Scholar]
- Yumioka, R.; Sato, H.; Tomizawa, H.; Yamasaki, Y.; Ejima, D. Mobile phase containing arginine provides more reliable SEC condition for aggregation analysis. J. Pharm. Sci. 2010, 99, 618–620. [Google Scholar]
- Arakawa, T.; Ejima, D.; Li, T.; Philo, J.S. The critical role of mobile phase composition in size exclusion chromatography of protein pharmaceuticals. J. Pharm. Sci. 2010, 99, 1674–1692. [Google Scholar]
- Herce, H.D.; Garcia, A.E.; Litt, K.; Kane, R.S.; Martin, P.; Enrique, N.; Rebolledo, A.; Milesi, V. Arginine-rich peptides destabilize the plasma membrane, consistent with a pore formation translocation mechanism of cell penetrating peptides. Biophys. J. 2009, 97, 1917–1925. [Google Scholar]
- Bouchet, A.; Lairion, F.; Disalvo, E.A. Role of guanidinium group in the interaction of l-arginine in DMPE and DMPC lipid interphases. Biochim. Biophys. Acta 2009, 1798, 616–623. [Google Scholar]
- Schmidt, N.; Mishra, A.; Lai, G.H.; Wong, G.C. Arginine-rich cell-penetrating peptides. FEBS Lett. 2009. [Google Scholar] [CrossRef]
- Fukuyama, K.; Abdel-Mequid, S.S.; Johnson, J.E.; Rossmann, M.G. Structure of a T = 1 aggregate of alfalfa mosaic virus coat protein seen at 4.5 A resolution. Mol. Biol. 1983, 167, 873–894. [Google Scholar] [CrossRef]
- Rould, M.A.; Perona, J.J.; Steitz, T.A. Structure of E. coli glutaminyl-tRNA synthetase complexed with tRNA(Gln) and ATP at 2.8 A resolution. Science 1989, 246, 135–1142. [Google Scholar]
- Xu, S.; Zheng, W.; Radisic, D.; Bowen, K.H., Jr. The stabilization of arginine’s zwitterions by dipole-binding of an excess electron. J. Chem. Phys. 2005, 122, 091103. [Google Scholar] [CrossRef]
- Yoo, J.; Cui, Q. Does arginine remain protonated in the lipid membrane? Insight from microscopic pKa calculations. Biophys. J. 2008, 94, L61–L63. [Google Scholar]
- Kita, Y.; Arakawa, T.; Lin, T.Y.; Timasheff, S.N. Contribution of the surface free energy perturbation to protein-solvent interactions. Biochemistry 1994, 33, 15178–15189. [Google Scholar]
- Schneider, C.P.; Trout, B.L. Investigation of cosolute-protein preferential interaction coefficients: new insight into the mechanism by which arginine inhibits aggregation. J. Phys. Chem. 2009, 113, 2050. [Google Scholar]
- Arakawa, T.; Timasheff, S.N. Preferential interactions of proteins with salts in concentrated solutions. Biochemistry 1982, 21, 6545–6552. [Google Scholar]
- Arakawa, T.; Timasheff, S.N. The mechanism of action of Na glutamate, lysine HCl, and piperazine-N,N’-bis(2-ethanesulfonic acid) in the stabilization of tubulin and microtuble formation. J. Biol. Chem. 1984, 259, 4979–4986. [Google Scholar]
- Arakawa, T.; Timasheff, S.N. The stabilization of proteins by osmolytes. Biophys. J. 1985, 47, 411–414. [Google Scholar] [CrossRef]
- Ausar, S.F.; Espina, M.; Brock, J.; Thyagarayapuran, N.; Repetto, R.; Khandke, L.; Middaugh, C.R. High-throughput screening of stabilizers for respiratory syncytial virus. Human Vaccines 2007, 3, 68–77. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ohtake, S.; Arakawa, T.; Koyama, A.H. Arginine as a Synergistic Virucidal Agent. Molecules 2010, 15, 1408-1424. https://doi.org/10.3390/molecules15031408
Ohtake S, Arakawa T, Koyama AH. Arginine as a Synergistic Virucidal Agent. Molecules. 2010; 15(3):1408-1424. https://doi.org/10.3390/molecules15031408
Chicago/Turabian StyleOhtake, Satoshi, Tsutomu Arakawa, and A. Hajime Koyama. 2010. "Arginine as a Synergistic Virucidal Agent" Molecules 15, no. 3: 1408-1424. https://doi.org/10.3390/molecules15031408