Pharmacogenomic Identification of c-Myc/Max-Regulated Genes Associated with Cytotoxicity of Artesunate towards Human Colon, Ovarian and Lung Cancer Cell Lines


Abstract
:Abbreviations
| ABC | = ATP-binding cassette transporter |
| ACT | = artemisinin-based combination therapy |
| ART | = artesunate |
| CYP | = cytochrome P450 |
| DEPC | = diethylpyrocarbonat |
| DMSO | = dimethylsulfoxide |
| EGFR | = Epidermal Growth Factor Receptor |
| iNOS | = inducible nitric oxide synthase |
| IPA | = Ingenuity Pathway Analysis |
| LPS | = lipopolysaccharide |
| MAX | = Myc-associated factor x |
| MDR | = multidrug resistance |
| MTT | = 3-(4,5-dimethyl-thizol-2-yl)-2,5-diphenyltetrazolium bromide |
| NCI | = National Cancer Institute |
| NF-κB | = nuclear factor 'kappa-light-chain-enhancer' of activated B-cells |
| NSCLC | = non-small cell lung cancer |
| PCR | = polymerase chain reaction |
| PDK | = pyruvate dehydrogenase kinase |
| Pgp | = P-glycoprotein |
| PI3K | = phosphatidylinositol 3'-kinase |
| PK | = protein kinase |
| PKC | = protein kinase C |
| ROS | = reactive oxygen species |
| qRT-PCR | = quantitative real-time PCR |
| SDS | = sodium dodecyl sulphate |
| SSC | = standard saline citrate |
| TCM | = traditional Chinese medicine |
| TGF-β1 | = transforming growth factor-beta 1 |
| TNF | = tumor necrosis factor |
| WHO | = World Health Organization |
1. Introduction
2. Results
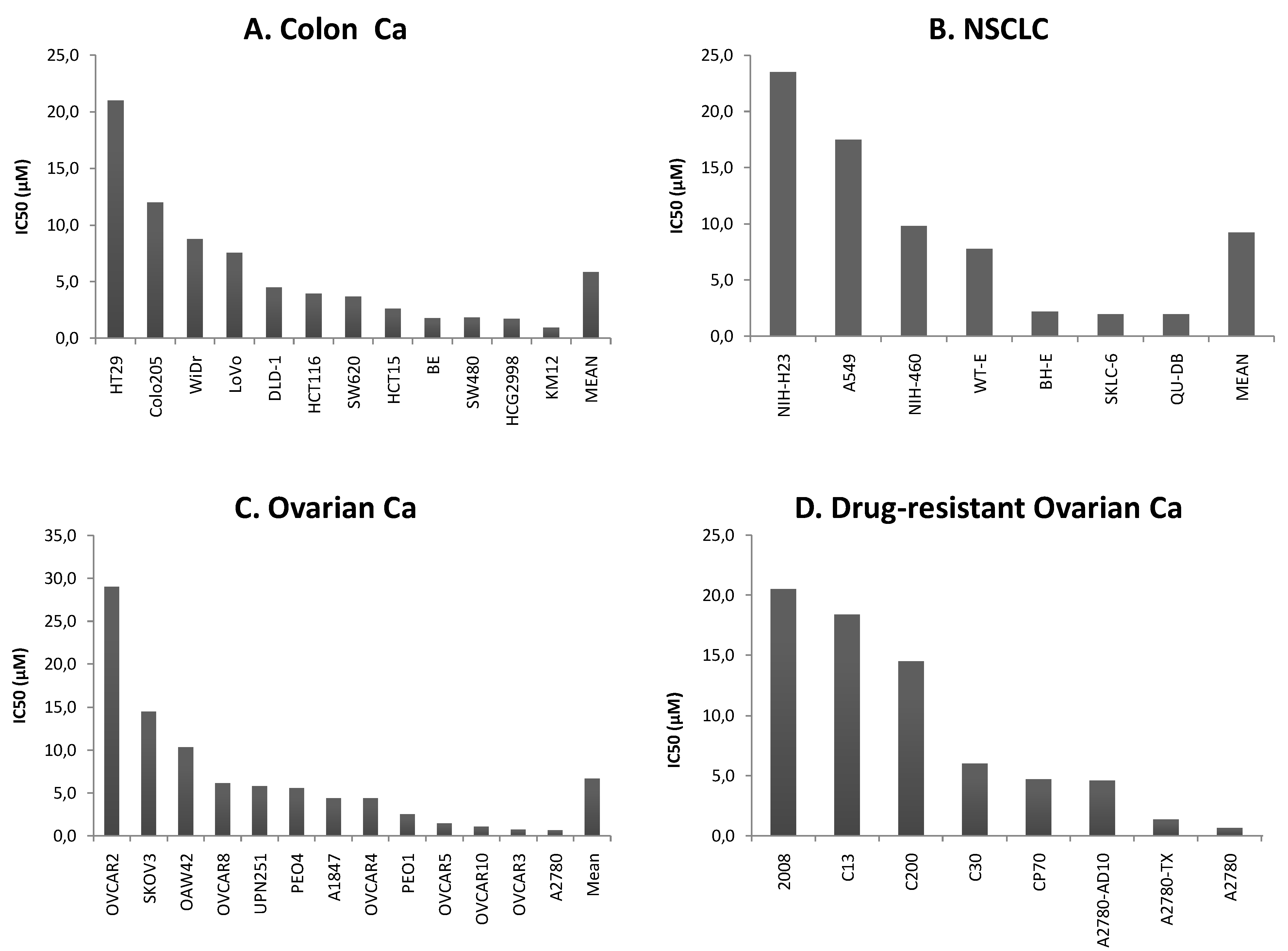
2.1. Cytotoxicity of cell lines

2.2. Microarray hybridization

{kind=link}
{kind=link}
{kind=link}
| RESISTANCE GENES | ||||
|---|---|---|---|---|
| Gene | UNIQID | Pearson | Name | Function |
| SLC30A1 | R31110 | 0.552 | Human hbc647 mRNA sequence | Involved in zinc transport out of the cell. |
| GFAP | AA069414 | 0.552 | Glial fibrillary acidic protein | Class-III intermediate filament |
| BDKRB2 | AA194043 | 0.513 | Bradykinin receptor B2 | Receptor for bradykinin |
| AKR1C3 | AA916325 | 0.511 | Aldo-keto reductase family 1, member C3 | Member of the aldo/keto reductase superfamily |
| SNAP23 | N32278 | 0.472 | Synaptosomal-associated protein, 23kD | Regulator of membrane fusion machinery and transport vesicle docking |
| KYNU | H87471 | 0.471 | Kynureninase (L-kynurenine hydrolase) | Cleaves L-kynurenine and L-3-hydroxykynurenine |
| AKR1C1 | R93124 | 0.453 | Aldo-keto reductase family 1, member C1 | Member of the aldo/keto reductase superfamily |
| ITPK1 | AA463931 | 0.452 | Inositol 1,3,4-triphosphate 5/6 kinase | Phosphorylates various inositol polyphosphates |
| not specified | H44051 | 0.443 | not specified | Member of the keratin family |
| ALDH3A1 | AA069024 | 0.435 | Aldehyde dehydrogenase 3 family, member A1 | ALDH oxidizes various aldehydes to the corresponding acids. |
| GYS2 | N52282 | 0.432 | Glycogen synthase 2 (liver) | Transfers the glycosyl residue from UDP-Glc to the non-reducing end of alpha-1,4-glucan |
| KRT4 | AA629189 | 0.425 | Keratin 4 | Member of the keratin gene family |
| CD24 | H59916 | 0.423 | CD24 antigen (small cell lung carcinoma) | Signal transducer; modulates B-cell activation |
| SMAD3 | W72201 | 0.423 | Similar to mothers against decapentaplegic homolog 3 | Transcriptional modulator; plays a role in carcinogenesis |
| CREB3L3 | AI952482 | 0.415 | Sirtuin 6 | Glycosyl transferase activity |
| SLC12A7 | AA427732 | 0.402 | Solute carrier 12 (potassium/chloride transporters) | Important for the survival of cochlear outer and inner hair cells |
| SLC22A5 | AI933276 | 0.398 | Solute carrier 22 (organic cation transporter) | Involved in the active cellular uptake of carnitine. |
| FETUB | AA705308 | 0.396 | Fetuin B | Member of the fetuin family |
| TRAM1 | H15707 | 0.366 | Translocating chain-associating membrane protein | Translocation of secretory proteins across the ER membrane |
| WNT7A | AI885451 | 0.365 | Wingless-type MMTV integration site family | Involved in oncogenesis |
| DCDC2 | AA127741 | 0.360 | Doublecortin domain containing 2 | Enhances microtubule polymerization |
| ERRFI1 | AA400258 | 0.359 | Gene 33/Mig-6 | Involved in cell signaling and cell stress |
| ABCC3 | AA429895 | 0.357 | ATP-binding cassette, sub-family C (CFTR/MRP) | Member of the superfamily of ABC-Transporters |
| IKBKG | R56102 | 0.357 | Homo sapiens cDNA FLJ20586 fis, clone KAT09466 | Regulatory subunit of the inhibitor of kappaB kinase (IKK) |
| S100A10 | AA444051 | 0.354 | S100 calcium-binding protein A10 | Regulation of cellular processes such as cell cycle progression and differentiation |
| GSR | AA777289 | 0.352 | Glutathione reductase | Maintains high levels of reduced glutathione in the cytosol |
| ST8SIA1 | AA169183 | 0.349 | Sialyltransferase 8 A | Cell adhesion and growth of cultured malignant cells |
| CUL5 | AA086475 | 0.337 | Cullin 5 | May form a cell surface vasopressin receptor |
| SCAP | R54823 | 0.335 | Srebp cleavage-activating protein | Regulates sterol biosynthesis |
| AKR1B1 | AA701963 | 0.330 | Aldo-keto reductase family 1, member B1 | Member of the aldo/keto reductase superfamily |
| ATP1B1 | AA598814 | 0.329 | ATPase, Na+/K+ transporting, beta 1 polypeptide | Catalyzes ATP hydrolysis and ion exchange across plasma membranes |
| VCAN | AA101875 | 0.326 | Chondroitin sulfate proteoglycan 2 (versican) | May play a role in intercellular signaling, cell mobility and differentiation |
| TRIM21 | N45131 | 0.309 | Sjögren syndrome antigen A1, tripartite motif-containing 21 | Ribonucleoprotein particle which binds DNA, RNA, protein and zinc |
| GLRX | AA291163 | 0.307 | Glutaredoxin (thioltransferase) | Reduces low molecular weight disulfides and proteins |
| SLC23A1 | AI934925 | 0.306 | Solute carrier family 23 (nucleobase transporters) | This gene encodes one of the Vitamin C transporters |
| SENSITIVITY GENES | ||||
| Gene | UNIQID | Pearson | Name | Function |
| RPL29 | AW073449 | -0.300 | Ribosomal protein L29 | Cytoplasmic ribosomal protein of the 60S subunit |
| PSMB5 | AA864479 | -0.304 | Proteasome (prosome, macropain) subunit, β type | May catalyze basal processing of intracellular antigens |
| DDR2 | AA243749 | -0.308 | Discoidin domain receptor family, member 2 | Tyrosine kinase receptor mediating fibroblast migration and proliferation |
| ASNS | AA894927 | -0.309 | Asparagine synthetase | Synthesis of asparagine |
| LDHB | AI969670 | -0.310 | Lactate dehydrogenase B | Oxidoreductase activity |
| RAD23A | H99170 | -0.310 | RAD23 homolog A (S. cerevisiae) | Molecular calcium binding chaperone |
| CSE1L | N69204 | -0.312 | Chromosome segregation 1 (yeast homolog)-like | May play a role in apoptosis and cell proliferation |
| INSIG1 | H59620 | -0.314 | Insulin induced gene 1 | May play a role in growth and differentiation of tissues involved in metabolic control |
| PABPCP5 | AA486531 | -0.315 | Poly(A)-binding protein, cytoplasmic 1 | May be involved in translationally coupled mRNA turnover |
| ATOX1 | AA418694 | -0.315 | Antioxidant protein 1 (yeast) homolog 1 | May play a role in carcinogenesis |
| PRKCSH | AA496810 | -0.316 | Protein kinase C substrate 80K-H | Acidic phospho-protein known to be a substrate for protein kinase C |
| AA878561 | -0.319 | Ubiquitin A-52 residue ribosomal protein product | Regulation of gene expression | |
| ISG15 | AA406020 | -0.319 | Interferon-stimulated protein, 15 kDa | May regulate proteins involved in the release of prostaglandin F2-alpha (PGF) |
| EEF2 | R43766 | -0.322 | Eukaryotic translation elongation factor 2 | Essential factor for protein synthesis |
| R37276 | -0.322 | Eukaryotic translation initiation factor 4 gamma, 1 | Recognition of the mRNA cap | |
| PCDH17 | AA969048 | -0.326 | Protocadherin 17 | May play a role for cell-cell connections in the brain |
| PHB2 | AA464567 | -0.328 | B-cell associated protein | Functions as an estrogen receptor |
| TIMM17A | AA708446 | -0.335 | Translocase of inner mitochondrial membrane 17 | Translocation of transit proteins across mitochondrial membrane |
| RRAS | AI368184 | -0.337 | Related RAS viral (r-ras) oncogene homolog | Regulates the organization of the actin cytoskeleton |
| SYNCRIP | AA186327 | -0.338 | Synaptotagmin binding, cytoplasmic RNA binding protein | Involved in mRNA processing |
| AA448261 | -0.345 | High-mobility group protein isoforms I and Y | Gene transcription, integration of retroviruses into chromosomes and metastasis | |
| RPL18A | W81118 | -0.356 | EST, similar to human 60S Ribosomal Protein L18A | unknown |
| RPS10 | AI611010 | -0.357 | Ribosomal protein S10 | Catalyzes protein synthesis. Variable expression of this gene in colorectal cancers |
| UBB | AW078798 | -0.358 | Ubiquitin B | Regulation of gene expression |
| RPSA | AA629897 | -0.367 | Laminin receptor 1 (67kD, ribosomal protein SA) | Up-regulation in cancer cells associated with invasion and metastasis |
| PRMT1 | N55480 | -0.381 | HMT1 (hnRNP methyltransferase)-like 2 | Arginine methyltransferase |
| LMAN1 | AA446103 | -0.386 | Lectin, mannose-binding, 1 | Type I integral membrane protein |
| YBX1 | AA599175 | -0.418 | Nuclease sensitive element binding protein 1 | May play a role in DNA repair |
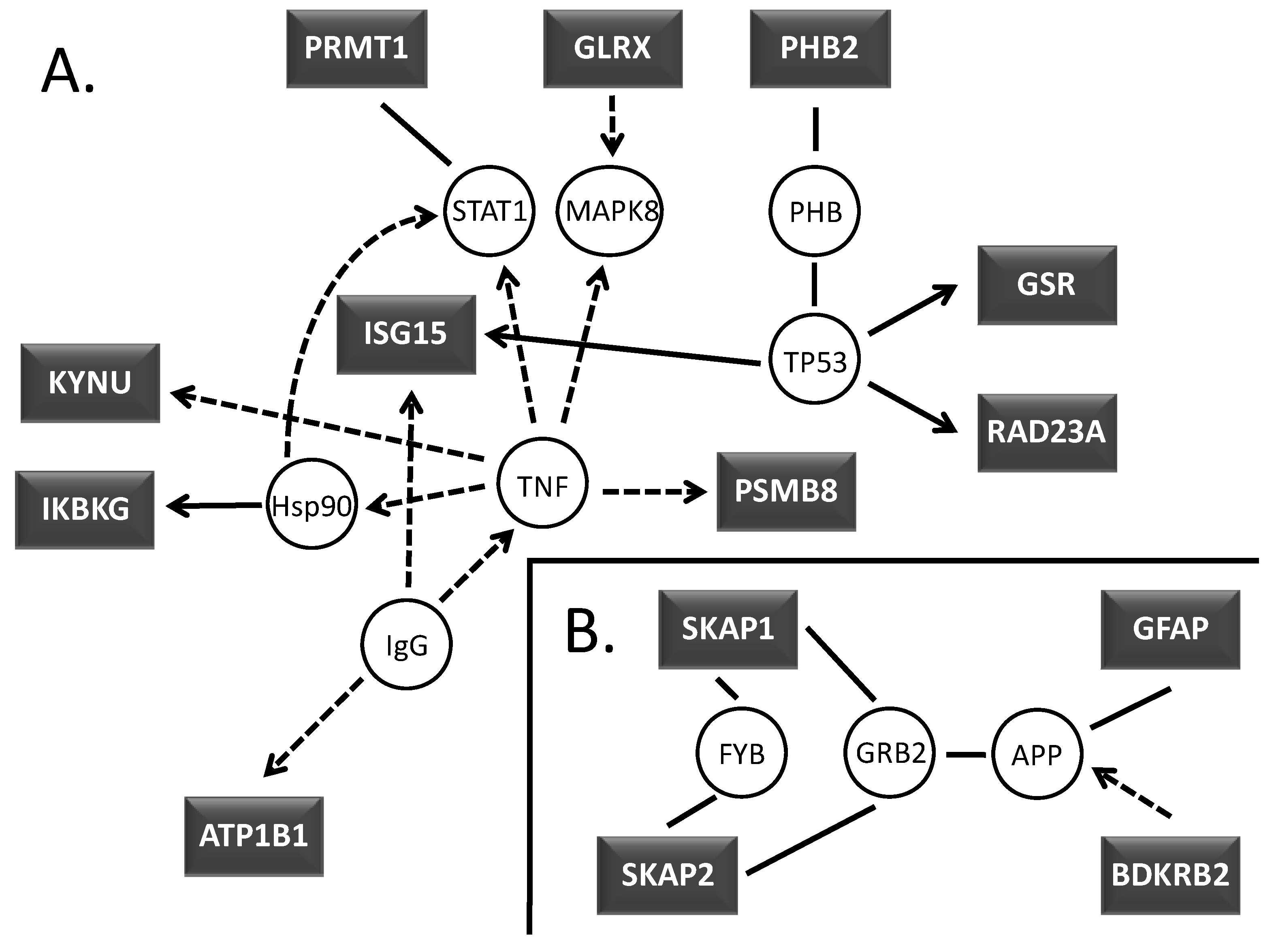
2.3. Signaling pathway profiling

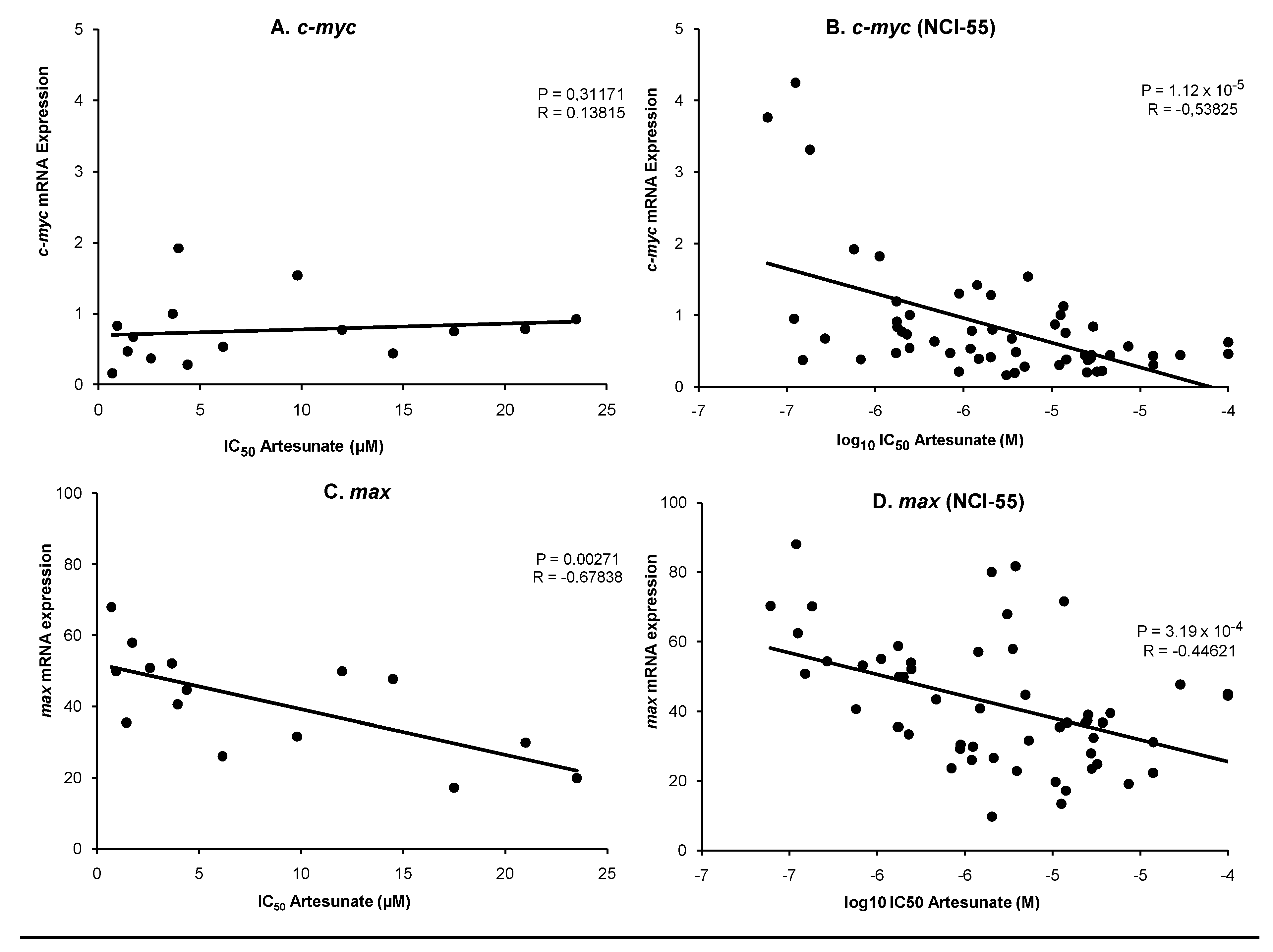
2.4. Transcription factor downstream gene analyses
| Gene | Number of potential binding sites | Position (Score) | Strand |
|---|---|---|---|
| SLC30A1 | 3 | -2645 (7,989), -910 (8,955), -909 (9,341) | +, +, - |
| GFAP | 2 | -4972 (8,199), -161 (9,706) | +, - |
| BDKRB2 | 7 | -4885 (9,431), -4884 (8,449), -3439 (8,229), -2826 (8,977), -2825 (8,229), -1493 (8,820), -1466 (8,372) | +, -, +, +, -, +, + |
| AKR1C3 | 4 | -3486 (9,956), -3182 (8,252), -1029 (7,972), -71 (9,667) | +, -, -, - |
| SNAP23 | 3 | -4877 (8,036), -857 (8,449), -856 (9,431) | -, +, - |
| KYNU | No binding sites detected | ||
| AKR1C1 | 1 | -4606 (8,039) | + |
| ITPK1 | 1 | -2885 (8,890) | - |
| GYS2 | 1 | -4280 (8,960) | - |
| KRT4 | 12 | -4540 (9,810), -4539 (8,393), -3905 (8,504), -3532 (9,421), -3531 (10,337), -3442 (8,372), -2264 (9,431), -2263 (8,449), -1942 (10,480), -1941 (13,638), -1016 (8,841), -165 (9,118) | +, +, -, +, -, +, +, -, +, -, -, + |
| CD24 | 3 | -2956 (9,104), -2955 (9,909), -2906 (8,055) | +, -, - |
| SMAD3 | 5 | -3843 (7,989), -3135 (9,044), -2558 (8,372), -418 (9,341), -417 (9,341) | -, -, -, +, - |
| CREB3L3 | 9 | -4911 (10,153), -3674 (8,186), -32229 (8,162), -2802 (8,449), -2801 (9,431), -2073 (10,396), -1327 (9,585), -485 (9,431), -484 (8,449) | -, +, +, +, -, +, +, +, - |
| SLC12A7 | 11 | -4608 (10,961), -4607 (9,095), -2657 (8,252), -2584 (10,769), -2204 (8,504), -1970 (9,134), -1862 (8,820), -1209 (9,238), -1016 (8,229), -61 (10,468), -60 (9,869) | +, -, -, -, -, +, -, -, -, +, - |
| SLC22A5 | No binding sites detected | ||
| FETUB | 6 | -3871 (8,074), -1418 (8,464), -804 (10,572), -803 (8,441), -418 (9,706), -69 (9,983) | -, +, +, -, +, - |
| TRAM1 | 3 | -4348 (8,890), -4347 (9,552), -3118 (8,515) | +, -, + |
| WNT7A | 3 | -4745 (9,706), -2862 (9,491), -2664 (8,960) | -, +, + |
| DCDC2 | 1 | -2660 (8,259) | + |
| ERRFI1 | 2 | -4501 (8,372), -4141 (8,761) | +, + |
| ABCC3 | 5 | -4552 (8,120), -4326 (7,989), -4097 (8,229), -4064 (9,044), -4063 (8,229) | +, +, +, +, - |
| IKBKG | 3 | -952 (9,431), -951 (8,449), -543 (9,421) | +, -, - |
| S100A10 | 8 | -4729 (8,687), -4728 (9,828), -4050 (8,178), -3913 (10,807), -3912 (9,350), -2303 (9,666), -2272 (8,449), -2271 (9,431) | +, -, +, +, -, -, +, - |
| GSR | 6 | -3066 (9,667), -3065 (8,199), -2431 (9,726), -2043 (8,017), -905 (10,023), -904 (8,229) | +, -, -, -, +, - |
| ST8SIA1 | 3 | -3532 (11,197), -3531 (8,039), -2001 (8,841) | +, -, + |
| CUL5 | 3 | -4798 (8,960), -643 (8,504), -642 (8,754) | +, +, - |
| SCAP | 2 | -3904 (8,449), -3903 (9,431) | +, - |
| AKR1B1 | 5 | -3654 (8,372), -3594 (8,820), -3429 (9,409), -2950 (8,229), -2636 (8,960) | -, +, -, -, + |
| ATP1B1 | 6 | -4514 (11,479), -4513 (10,785), -4151 (9,902), -2867 (9,095), -1545 (8,252), -1136 (8,960) | +, -, -, +, +, - |
| VCAN | 2 | -1256 (9,350), -224 (8,055) | +, - |
| TRIM21 | 7 | -3372 (10,470), -3371 (9,706), -2169 (9,118), -2043 (8,322), -407 (8,055), -207 (8,515), -206 (10,627) | +, -, +, +, +, +, - |
| GLRX | 7 | -2391 (9,431), -2390 (8,449), -1766 (8,687), -1765 (8,566), -1108 (9,095), -1107 (8,485), -115 (8,213) | +, -, +, -, +, -, - |
| SLC23A1 | 3 | -4955 (8,047), -3370 (12,419), -2782 (8,372) | +, -, + |
| RPL29 | 7 | -4179 (8,147), -2366 (8,372), -703 (9,552), -702 (8,890), -624 (9,277), -593 (8,449), -592 (9,431) | -, -, +, -, -, +, - |
| PSMB5 | 6 | -3113 (8,449), -3112 (9,431), -1295 (9,431), -1294 (8,449), -1105 (9,431), -1104 (8,449) | +, -, +, -, +, - |
| ASNS | 3 | -4092 (9,277), -2283 (9,104), -2087 (10,491) | +, -, + |
| LDHB | 4 | -4479 (9,491), -830 (9,902), -563 (9,431), -562 (8,449) | -, +, +, - |
| RAD23A | 5 | -2525 (8,449), -2524 (9,431), -156 (9,095), -155 (11,278), -37 (8,607) | +, -, +, -, - |
| INSIG1 | 5 | -4207 (8,252), -3881 (8,259), -3535 (8,687), -2754 (9,421), -161 (8,754) | +, -, -, +, - |
| PABPCP5 | 3 | -1789 (8,449), -1788 (9,431), -454 (9,341) | +, -, + |
| ATOX1 | 9 | -4359 (8,199), -4358 (11,356), -2830 (9,134), -2656 (9,706), -2618 (8,017), -2313 (9,431), -2312 (8,449), -947 (9,949), -121 (9,956) | +, -, +, +, +, +, -, - |
| PRKCSH | 9 | -4688 (8,372), -4621 (9,491), -3737 (9,431), -3736 (8,449), -2854 (10,889), -2765 (8,298), -2764 (10,092), -121 (8,830), -120 (11,519) | -, -, +, -, +, +, -, +, - |
| ISG15_ | 5 | -3964 (9,431), -3963 (8,449), -1932 (10,627), -1931 (12,578), -1603 (8,218) | +, -, +, -, - |
| EEF2 | 6 | -3734 (10,572), -3278 (9,552), -3277 (8,890), -2791 (9,431), -2790 (8,449), -1565 (8,515) | -, +, -, +, -, - |
| PCDH17 | 1 | -501 (8,199) | - |
| TIMM17A | 11 | -3771 (8,047), -3657 (8,952), -2621 (9,431), -2620 (8,449), -2157 (8,923), -1445 (9,431), -1444 (9,552), -1084 (9,431), -623 (9,431), -622 (8,449), -170 (8,047) | +, +, +, -, +, +, -, +, +, -, + |
| RRAS | 4 | -4729 (8,055), -3576 (9,104), -3575 (8,441), -230 (10,403) | -, +, -, + |
| SYNCRIP | 4 | -4703 (7.977), -1609 (10,480), -1608 (12,721), -77 (8,607) | +, +, -, - |
| RPS10 | 2 | -3402 (7,980), -1980 (7,980) | +, - |
| UBB | 4 | -4351 (9,431), -4350 (8,449), -1719 (8,305), -1268 (9,350) | +, -, -, + |
| RPSA | No binding sites detected | ||
| PRMT1 | 6 | -4832 (8,305), -4831 (8,372), -4430 (8,441), -1711 (8,393), -763 (10,153), -762 (9,706) | +, -, +, -, +, - |
| LMAN1 | 1 | -511 (8,444) | - |
| YBX1 | 6 | -3039 (8,464), -2000 (9,585), -1590 (8,960), -1589 (9,277), -848 (11,455), -847 (10,755) | -, +, +, -, +, - |

3. Discussion
3.1. Identification of candidate genes by microarray and COMPARE analyses
3.2. Signaling pathway analyses
3.3. Transcription factor downstream gene analyses
4. Experimental
4.1. Drugs and reagents
4.2. Cell lines
4.3. MTT assay
4.4. RNA isolation
4.5. Preparation of cDNA probes
4.6. cDNA microarray screening
4.7. Data analyses
4.8. Statistical analyses
4.9. Identification of signaling pathways
4.10. Binding motif analyses in gene promoters
5. Conclusions
Conflict of Interest
- Sample Availability: Samples of ART are available from Saokim Co. Ltd. (Hanoi, Vietnam).
References
- Klayman, D.L. Qinghaosu (artemisinin): An antimalarial drug from China. Science 1985, 228, 1049–1055. [Google Scholar]
- Tu, Y. The development of new antimalarial drugs: Qinghaosu and dihydro-qinghaosu. Chin. Med. J. (Engl.) 1999, 112, 976–977. [Google Scholar]
- Li, Y.; Wu, Y.L. How Chinese scientists discovered qinghaosu (artemisinin) and developed its derivatives? What are the future perspectives? Med. Trop. (Mars) 1998, 58, 9–12. [Google Scholar]
- Efferth, T. Willmar Schwabe Award 2006: Antiplasmodial and antitumor activity of artemisinin-from bench to bedside. Planta Med. 2007, 73, 299–309. [Google Scholar] [CrossRef]
- Efferth, T.; Fu, Y.J.; Zu, Y.G.; Schwarz, G.; Konkimalla, V.S.; Wink, M. Molecular target-guided tumor therapy with natural products derived from traditional Chinese medicine. Curr. Med. Chem. 2007, 14, 2024–2032. [Google Scholar]
- Efferth, T.; Giaisi, M.; Merling, A.; Krammer, P.H.; Li-Weber, M. Artesunate induces ROS-mediated apoptosis in doxorubicin-resistant T leukemia cells. PLoS One 2007, 2, e693. [Google Scholar] [CrossRef]
- Efferth, T.; Li, P.C.; Konkimalla, V.S.; Kaina, B. From traditional Chinese medicine to rational cancer therapy. Trends Mol. Med. 2007, 13, 353–361. [Google Scholar] [CrossRef]
- Efferth, T.; Rucker, G.; Falkenberg, M.; Manns, D.; Olbrich, A.; Fabry, U.; Osieka, R. Detection of apoptosis in KG-1a leukemic cells treated with investigational drugs. Arzneimittelforschung 1996, 46, 196–200. [Google Scholar]
- Efferth, T. Microarray-based prediction of cytotoxicity of tumor cells to cantharidin. Oncol. Rep. 2005, 13, 459–463. [Google Scholar]
- Efferth, T.; Dunstan, H.; Sauerbrey, A.; Miyachi, H.; Chitambar, C.R. The anti-malarial artesunate is also active against cancer. Int. J. Oncol. 2001, 18, 767–773. [Google Scholar]
- Efferth, T.; Briehl, M.M.; Tome, M.E. Role of antioxidant genes for the activity of artesunate against tumor cells. Int. J. Oncol. 2003, 23, 1231–1235. [Google Scholar]
- Efferth, T. Mechanistic perspectives for 1,2,4-trioxanes in anti-cancer therapy. Drug Resist. Updat. 2005, 8, 85–97. [Google Scholar] [CrossRef]
- Berger, T.G.; Dieckmann, D.; Efferth, T.; Schultz, E.S.; Funk, J.O.; Baur, A.; Schuler, G. Artesunate in the treatment of metastatic uveal melanoma--first experiences. Oncol. Rep. 2005, 14, 1599–1603. [Google Scholar]
- Efferth, T. Molecular pharmacology and pharmacogenomics of artemisinin and its derivatives in cancer cells. Curr. Drug Targets 2006, 7, 407–421. [Google Scholar] [CrossRef]
- Kelter, G.; Steinbach, D.; Konkimalla, V.B.; Tahara, T.; Taketani, S.; Fiebig, H.H.; Efferth, T. Role of transferrin receptor and the ABC transporters ABCB6 and ABCB7 for resistance and differentiation of tumor cells towards artesunate. PLoS One 2007, 2, e798. [Google Scholar] [CrossRef]
- Scherf, U.; Ross, D.T.; Waltham, M.; Smith, L.H.; Lee, J.K.; Tanabe, L.; Kohn, K.W.; Reinhold, W.C.; Myers, T.G.; Andrews, D.T.; Scudiero, D.A.; Eisen, M.B.; Sausville, E.A.; Pommier, Y.; Botstein, D.; Brown, P.O.; Weinstein, J.N. A gene expression database for the molecular pharmacology of cancer. Nat. Genet. 2000, 24, 236–244. [Google Scholar] [CrossRef]
- Staunton, J.E.; Slonim, D.K.; Coller, H.A.; Tamayo, P.; Angelo, M.J.; Park, J.; Scherf, U.; Lee, J.K.; Reinhold, W.O.; Weinstein, J.N.; Mesirov, J.P.; Lander, E.S.; Golub, T.R. Chemosensitivity prediction by transcriptional profiling. Proc. Natl. Acad. Sci. USA 2001, 98, 10787–10792. [Google Scholar]
- Paull, K.D.; Lin, C.M.; Malspeis, L.; Hamel, E. Identification of novel antimitotic agents acting at the tubulin level by computer-assisted evaluation of differential cytotoxicity data. Cancer Res. 1992, 52, 3892–3900. [Google Scholar]
- Leteurtre, F.; Kohlhagen, G.; Paull, K.D.; Pommier, Y. Topoisomerase II inhibition and cytotoxicity of the anthrapyrazoles DuP 937 and DuP 941 (Losoxantrone) in the National Cancer Institute preclinical antitumor drug discovery screen. J. Nat. Cancer Inst. 1994, 86, 1239–1244. [Google Scholar]
- Shi, L.M.; Myers, T.G.; Fan, Y.; O'Connor, P.M.; Paull, K.D.; Friend, S.H.; Weinstein, J.N. Mining the National Cancer Institute Anticancer Drug Discovery Database: cluster analysis of ellipticine analogs with p53-inverse and central nervous system-selective patterns of activity. Mol. Pharmacol. 1998, 53, 241–251. [Google Scholar]
- Efferth, T.; Gebhart, E.; Ross, D.D.; Sauerbrey, A. Identification of gene expression profiles predicting tumor cell response to L-alanosine. Biochem. Pharmacol. 2003, 66, 613–621. [Google Scholar] [CrossRef]
- Efferth, T.; Olbrich, A.; Bauer, R. mRNA expression profiles for the response of human tumor cell lines to the antimalarial drugs artesunate, arteether, and artemether. Biochem. Pharmacol. 2002, 64, 617–623. [Google Scholar]
- Harris, E.H.; Burkhart, B.D.; Gillham, N.W.; Boynton, J.E. Antibiotic resistance mutations in the chloroplast 16S and 23S rRNA genes of Chlamydomonas reinhardtii: Correlation of genetic and physical maps of the chloroplast genome. Genetics 1989, 123, 281–292. [Google Scholar]
- Higgins, C.F. Multiple molecular mechanisms for multidrug resistance transporters. Nature 2007, 446, 749–757. [Google Scholar] [CrossRef]
- Efferth, T. The human ATP-binding cassette transporter genes: From the bench to the bedside. Curr. Mol. Med. 2001, 1, 45–65. [Google Scholar] [CrossRef]
- Gillet, J.P.; Efferth, T.; Remacle, J. Chemotherapy-induced resistance by ATP-binding cassette transporter genes. Biochim. Biophys. Acta 2007, 1775, 237–262. [Google Scholar]
- Kool, M.; van der Linden, M.; de Haas, M.; Scheffer, G.L.; de Vree, J.M.; Smith, A.J.; Jansen, G.; Peters, G.J.; Ponne, N.; Scheper, R.J.; Elferink, R.P.; Baas, F.; Borst, P. MRP3, an organic anion transporter able to transport anti-cancer drugs. Proc. Nat. Acad. Sci. USA 1999, 96, 6914–6919. [Google Scholar]
- Efferth, T.; Sauerbrey, A.; Olbrich, A.; Gebhart, E.; Rauch, P.; Weber, H.O.; Hengstler, J.G.; Halatsch, M.E.; Volm, M.; Tew, K.D.; Ross, D.D.; Funk, J.O. Molecular modes of action of artesunate in tumor cell lines. Mol. Pharmacol. 2003, 64, 382–394. [Google Scholar] [CrossRef]
- Sertel, S.; Eichhorn, T.; Sieber, S.; Sauer, A.; Weiss, J.; Plinkert, P.K.; Efferth, T. Factors determining sensitivity or resistance of tumor cell lines towards artesunate. Chem. Biol. Interact. 2010, 185, 42–52. [Google Scholar] [CrossRef]
- Grant, S.; Qiao, L.; Dent, P. Roles of ERBB family receptor tyrosine kinases, and downstream signaling pathways, in the control of cell growth and survival. Front. Biosci. 2002, 7, d376–d389. [Google Scholar]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; Stivala, F.; Libra, M.; Basecke, J.; Evangelisti, C.; Martelli, A.M.; Franklin, R.A. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim. Biophys. Acta 2007, 1773, 1263–1284. [Google Scholar] [CrossRef]
- Konkimalla, V.B.; McCubrey, J.A.; Efferth, T. The role of downstream signaling pathways of the epidermal growth factor receptor for Artesunate's activity in cancer cells. Curr. Cancer Drug Targets 2009, 9, 72–80. [Google Scholar] [CrossRef]
- Muller, P.A.; Klomp, L.W. ATOX1: a novel copper-responsive transcription factor in mammals? Int. J. Biochem. Cell Biol. 2009, 41, 1233–1236. [Google Scholar] [CrossRef]
- Boal, A.K.; Rosenzweig, A.C. Crystal structures of cisplatin bound to a human copper chaperone. J. Am. Chem. Soc. 2009, 131, 14196–14197. [Google Scholar] [CrossRef]
- Guay, D.; Evoy, A.A.; Paquet, E.; Garand, C.; Bachvarova, M.; Bachvarov, D.; Lebel, M. The strand separation and nuclease activities associated with YB-1 are dispensable for cisplatin resistance but overexpression of YB-1 in MCF7 and MDA-MB-231 breast tumor cells generates several chemoresistance signatures. Int. J. Biochem. Cell Biol. 2008, 40, 2492–2507. [Google Scholar] [CrossRef]
- Tay, W.L.; Yip, G.W.; Tan, P.H.; Matsumoto, K.; Yeo, R.; Ng, T.P.; Kumar, S.D.; Tsujimoto, M.; Bay, B.H. Y-Box-binding protein-1 is a promising predictive marker of radioresistance and chemoradioresistance in nasopharyngeal cancer. Mod. Pathol. 2009, 22, 282–290. [Google Scholar] [CrossRef]
- Shibahara, K.; Sugio, K.; Osaki, T.; Uchiumi, T.; Maehara, Y.; Kohno, K.; Yasumoto, K.; Sugimachi, K.; Kuwano, M. Nuclear expression of the Y-box binding protein, YB-1, as a novel marker of disease progression in non-small cell lung cancer. Clin. Cancer Res. 2001, 7, 3151–3155. [Google Scholar]
- Gessner, C.; Woischwill, C.; Schumacher, A.; Liebers, U.; Kuhn, H.; Stiehl, P.; Jurchott, K.; Royer, H.D.; Witt, C.; Wolff, G. Nuclear YB-1 expression as a negative prognostic marker in nonsmall cell lung cancer. Eur. Respir. J. 2004, 23, 14–19. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Pastan, I. Biochemistry of multidrug resistance mediated by the multidrug transporter. Ann. Rev. Biochem. 1993, 62, 385–427. [Google Scholar] [CrossRef]
- Huang, J.; Tan, P.H.; Li, K.B.; Matsumoto, K.; Tsujimoto, M.; Bay, B.H. Y-box binding protein, YB-1, as a marker of tumor aggressiveness and response to adjuvant chemotherapy in breast cancer. Int. J. Oncol. 2005, 26, 607–613. [Google Scholar]
- Plebuch, M.; Soldan, M.; Hungerer, C.; Koch, L.; Maser, E. Increased resistance of tumor cells to daunorubicin after transfection of cDNAs coding for anthracycline inactivating enzymes. Cancer Lett. 2007, 255, 49–56. [Google Scholar] [CrossRef]
- Penning, T.M. Dihydrodiol dehydrogenase and its role in polycyclic aromatic hydrocarbon metabolism. Chem. Biol. Interact. 1993, 89, 1–34. [Google Scholar] [CrossRef]
- Wang, H.W.; Lin, C.P.; Chiu, J.H.; Chow, K.C.; Kuo, K.T.; Lin, C.S.; Wang, L.S. Reversal of inflammation-associated dihydrodiol dehydrogenases (AKR1C1 and AKR1C2) overexpression and drug resistance in nonsmall cell lung cancer cells by wogonin and chrysin. Int. J. Cancer 2007, 120, 2019–2027. [Google Scholar] [CrossRef]
- Sreerama, L.; Sladek, N.E. Primary breast tumor levels of suspected molecular determinants of cellular sensitivity to cyclophosphamide, ifosfamide, and certain other anticancer agents as predictors of paired metastatic tumor levels of these determinant. Rational individualization of cancer chemotherapeutic regimens. Cancer Chemother. Pharmacol. 2001, 47, 255–262. [Google Scholar] [CrossRef]
- Hu, G.; Chong, R.A.; Yang, Q.; Wei, Y.; Blanco, M.A.; Li, F.; Reiss, M.; Au, J.L.; Haffty, B.G.; Kang, Y. MTDH activation by 8q22 genomic gain promotes chemoresistance and metastasis of poor-prognosis breast cancer. Cancer Cell 2009, 15, 9–20. [Google Scholar] [CrossRef]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef]
- Bharti, A.C.; Aggarwal, B.B. Nuclear factor-kappa B and cancer: Its role in prevention and therapy. Biochem. Pharmacol. 2002, 64, 883–888. [Google Scholar] [CrossRef]
- Kellen, J.A. Molecular interrelationships in multidrug resistance (review). Anticancer Res. 1994, 14, 433–435. [Google Scholar]
- Lonning, P.E. Genes causing inherited cancer as beacons to identify the mechanisms of chemoresistance. Trends Mol. Med. 2004, 10, 113–118. [Google Scholar] [CrossRef]
- Daum, G. Lipid Metabolism and Membrane Biogenesis; Springer-Verlag: Berlin-Heidelberg, Germany, 2004. [Google Scholar]
- Cornwell, M.M.; Safa, A.R.; Felsted, R.L.; Gottesman, M.M.; Pastan, I. Membrane vesicles from multidrug-resistant human cancer cells contain a specific 150- to 170-kDa protein detected by photoaffinity labeling. Proc. Nat. Acad. Sci. USA 1986, 83, 3847–3850. [Google Scholar] [CrossRef]
- Dietel, M.; Arps, H.; Lage, H.; Niendorf, A. Membrane vesicle formation due to acquired mitoxantrone resistance in human gastric carcinoma cell line EPG85-257. Cancer Res. 1990, 50, 6100–6106. [Google Scholar]
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.; Mills, G.B. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, J.; Yang, N.; Greshock, J.; Liang, S.; Hasegawa, K.; Giannakakis, A.; Poulos, N.; O'Brien-Jenkins, A.; Katsaros, D.; Butzow, R.; Weber, B.L.; Coukos, G. Integrative genomic analysis of phosphatidylinositol 3'-kinase family identifies PIK3R3 as a potential therapeutic target in epithelial ovarian cancer. Clin. Cancer Res. 2007, 13, 5314–5321. [Google Scholar] [CrossRef]
- Humtsoe, J.O.; Kramer, R.H. Differential epidermal growth factor receptor signaling regulates anchorage-independent growth by modulation of the PI3K/AKT pathway. Oncogene 2009, 29, 1214–1226. [Google Scholar] [CrossRef]
- Chen, Y.X.; Wang, Y.; Fu, C.C.; Diao, F.; Song, L.N.; Li, Z.B.; Yang, R.; Lu, J. Dexamethasone enhances cell resistance to chemotherapy through increasing adhesion to extracellular matrix in human ovarian cancer cells. Endocr. Relat. Cancer 2009, 17, 39–50. [Google Scholar]
- Samarakoon, R.; Higgins, C.E.; Higgins, S.P.; Higgins, P.J. TGF-beta1-Induced Expression of the Poor Prognosis SERPINE1/PAI-1 Gene Requires EGFR Signaling: A New Target for Anti-EGFR Therapy. J. Oncol. 2009, 2009, 342391. [Google Scholar]
- Maglich, J.M.; Stoltz, C.M.; Goodwin, B.; Hawkins-Brown, D.; Moore, J.T.; Kliewer, S.A. Nuclear pregnane x receptor and constitutive androstane receptor regulate overlapping but distinct sets of genes involved in xenobiotic detoxification. Mol. Pharmacol. 2002, 62, 638–646. [Google Scholar] [CrossRef]
- Traber, M.G. Vitamin E, nuclear receptors and xenobiotic metabolism. Arch. Biochem. Biophys. 2004, 423, 6–11. [Google Scholar] [CrossRef]
- Konkimalla, V.B.; Blunder, M.; Korn, B.; Soomro, S.A.; Jansen, H.; Chang, W.; Posner, G.H.; Bauer, R.; Efferth, T. Effect of artemisinins and other endoperoxides on nitric oxide-related signaling pathway in RAW 264.7 mouse macrophage cells. Nitric Oxide 2008, 19, 184–191. [Google Scholar] [CrossRef]
- Efferth, T.; Benakis, A.; Romero, M.R.; Tomicic, M.; Rauh, R.; Steinbach, D.; Hafer, R.; Stamminger, T.; Oesch, F.; Kaina, B.; Marschall, M. Enhancement of cytotoxicity of artemisinins toward cancer cells by ferrous iron. Free Radical Biol. Med. 2004, 37, 998–1009. [Google Scholar] [CrossRef]
- Cody, G.D.; Boctor, N.Z.; Filley, T.R.; Hazen, R.M.; Scott, J.H.; Sharma, A.; Yoder, H.S., Jr. Primordial carbonylated iron-sulfur compounds and the synthesis of pyruvate. Science 2000, 289, 1337–1340. [Google Scholar]
- Lu, C.W.; Lin, S.C.; Chen, K.F.; Lai, Y.Y.; Tsai, S.J. Induction of pyruvate dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and drug resistance. J. Biol. Chem. 2008, 283, 28106–28114. [Google Scholar] [CrossRef]
- Chen, H.H.; Zhou, H.J.; Wang, W.Q.; Wu, G.D. Antimalarial dihydroartemisinin also inhibits angiogenesis. Cancer Chemother. Pharmacol. 2004, 53, 423–432. [Google Scholar] [CrossRef]
- Dell'Eva, R.; Pfeffer, U.; Vene, R.; Anfosso, L.; Forlani, A.; Albini, A.; Efferth, T. Inhibition of angiogenesis in vivo and growth of Kaposi's sarcoma xenograft tumors by the anti-malarial artesunate. Biochem. Pharmacol. 2004, 68, 2359–2366. [Google Scholar] [CrossRef]
- Roberts, D.; Schick, J.; Conway, S.; Biade, S.; Laub, P.B.; Stevenson, J.P.; Hamilton, T.C.; O'Dwyer, P.J.; Johnson, S.W. Identification of genes associated with platinum drug sensitivity and resistance in human ovarian cancer cells. Br. J. Cancer 2005, 92, 1149–1158. [Google Scholar] [CrossRef]
- Johnson, S.W.; Swiggard, P.A.; Handel, L.M.; Brennan, J.M.; Godwin, A.K.; Ozols, R.F.; Hamilton, T.C. Relationship between platinum-DNA adduct formation and removal and cisplatin cytotoxicity in cisplatin-sensitive and -resistant human ovarian cancer cells. Cancer Res. 1994, 54, 5911–5916. [Google Scholar]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Chomczynski, P.; Mackey, K. Substitution of chloroform by bromo-chloropropane in the single-step method of RNA isolation. Anal. Biochem. 1995, 225, 163–164. [Google Scholar] [CrossRef]
- Paull, K.D.; Shoemaker, R.H.; Hodes, L.; Monks, A.; Scudiero, D.A.; Rubinstein, L.; Plowman, J.; Boyd, M.R. Display and analysis of patterns of differential activity of drugs against human tumor cell lines: Development of mean graph and COMPARE algorithm. J. Nat. Cancer Inst. 1989, 81, 1088–1092. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sertel, S.; Eichhorn, T.; Simon, C.H.; Plinkert, P.K.; Johnson, S.W.; Efferth, T. Pharmacogenomic Identification of c-Myc/Max-Regulated Genes Associated with Cytotoxicity of Artesunate towards Human Colon, Ovarian and Lung Cancer Cell Lines. Molecules 2010, 15, 2886-2910. https://doi.org/10.3390/molecules15042886
Sertel S, Eichhorn T, Simon CH, Plinkert PK, Johnson SW, Efferth T. Pharmacogenomic Identification of c-Myc/Max-Regulated Genes Associated with Cytotoxicity of Artesunate towards Human Colon, Ovarian and Lung Cancer Cell Lines. Molecules. 2010; 15(4):2886-2910. https://doi.org/10.3390/molecules15042886
Chicago/Turabian StyleSertel, Serkan, Tolga Eichhorn, Christian H. Simon, Peter K. Plinkert, Steven W. Johnson, and Thomas Efferth. 2010. "Pharmacogenomic Identification of c-Myc/Max-Regulated Genes Associated with Cytotoxicity of Artesunate towards Human Colon, Ovarian and Lung Cancer Cell Lines" Molecules 15, no. 4: 2886-2910. https://doi.org/10.3390/molecules15042886