The Application of Ribozymes and DNAzymes in Muscle and Brain

Abstract
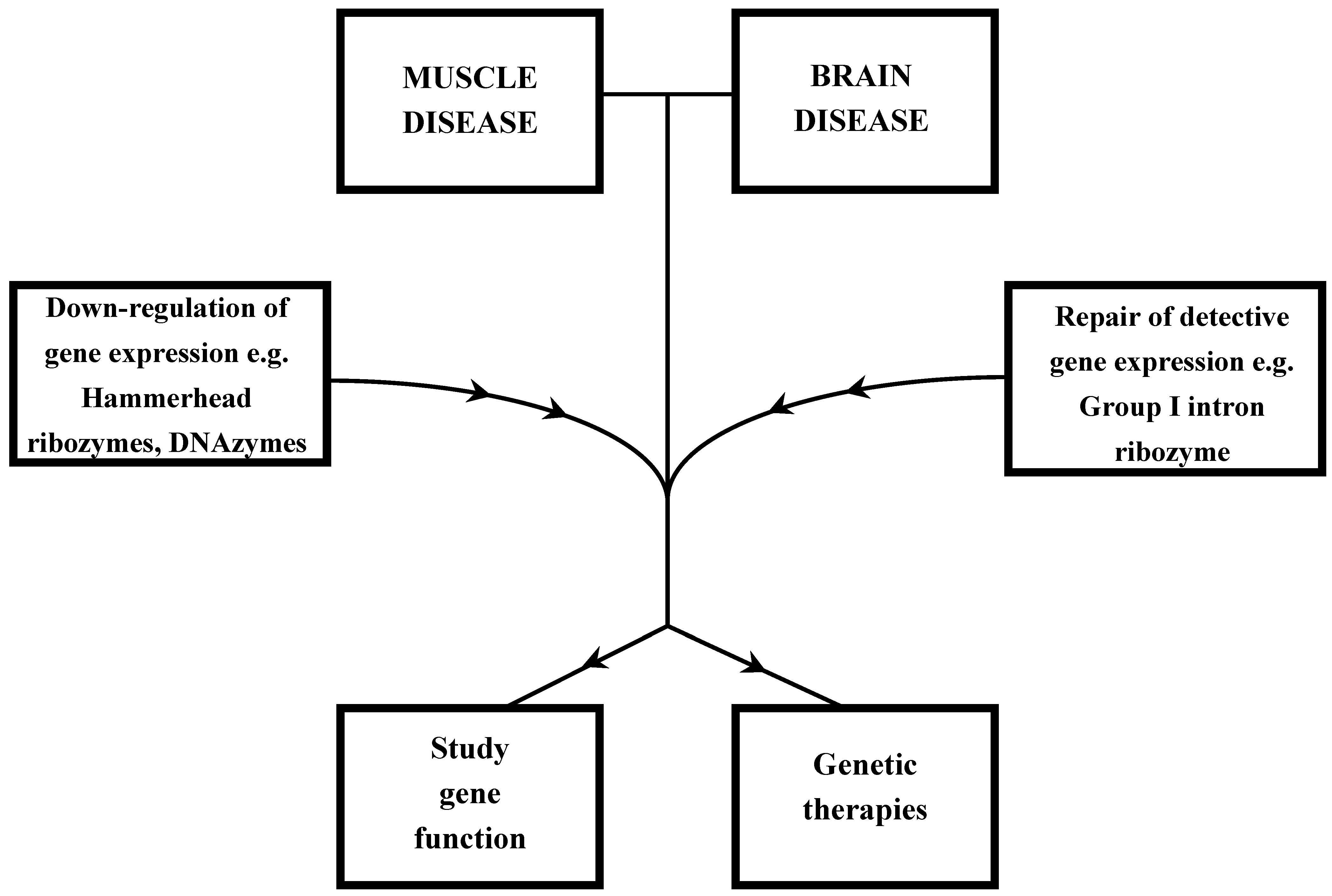
:1. Introduction
2. The Use of Ribozymes and DNAzymes in Muscle
3. The Use of Ribozymes and DNAzymes in Brain
4. Conclusions
Acknowledgements
References
- Rossi, J.J. Ribozymes. Curr. Opin. Biotechnol. 1992, 3, 3–7. [Google Scholar] [CrossRef]
- Haseloff, J.; Gerlach, W.L. Simple RNA enzymes with new and highly specific endoribonuclease activities. Nature 1988, 334, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, C.; Briguori, C.; De Rosa, R.; Ratti, G.; Cademartiri, F.; Sacco, M.; Borrelli, F.; Tammaro, P.; Midiri, M.; Capogrosso, P. Right coronary artery arising from pulmonary trunk: Assessment with conventional coronary angiography and multislice computed tomography coronary angiography. J. Cardiovasc. Med. 2009, 10, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Cech, T.R.; Zaug, A.J.; Grabowski, P.J. In vitro splicing of the ribosomal RNA precursor of Tetrahymena: Involvement of a guanosine nucleotide in the excision of the intervening sequence. Cell 1981, 27, 487–496. [Google Scholar] [CrossRef]
- Cech, T.R. Self-splicing of group I introns. Annu. Rev. Biochem. 1990, 59, 543–568. [Google Scholar] [CrossRef] [PubMed]
- Fiskaa, T.; Birgisdottir, A.B. RNA reprogramming and repair based on trans-splicing group I ribozymes. N. Biotechnol. 2010, 27, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Branch, A.D.; Robertson, H.D. Efficient trans cleavage and a common structural motif for the ribozymes of the human hepatitis delta agent. Proc. Natl. Acad. Sci. USA 1991, 88, 10163–10167. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.; Lande, C.; Cecchettini, A.; Citti, L. Hammerhead ribozymes in therapeutic target discovery and validation. Drug Discov. Today 2009, 14, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.L.; Choong, P.F.; Dass, C.R. DNAzyme delivery systems: Getting past first base. Expert Opin. Drug Deliv. 2009, 6, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Phylactou, L.A.; Kilpatrick, M.W.; Wood, M.J. Ribozymes as therapeutic tools for genetic disease. Hum. Mol. Genet. 1998, 7, 1649–1653. [Google Scholar] [CrossRef] [PubMed]
- Joyce, G.F. RNA cleavage by the 10-23 DNA enzyme. Methods Enzymol. 2001, 341, 503–517. [Google Scholar] [PubMed]
- Sun, L.Q.; Cairns, M.J.; Saravolac, E.G.; Baker, A.; Gerlach, W.L. Catalytic nucleic acids: From lab to applications. Pharmacol. Rev. 2000, 52, 325–347. [Google Scholar] [PubMed]
- Scanlon, K.J. Anti-genes: siRNA, ribozymes and antisense. Curr. Pharm. Biotechnol. 2004, 5, 415–420. [Google Scholar] [CrossRef] [PubMed]
- De Fougerolles, A.; Vornlocher, H.P.; Maraganore, J.; Lieberman, J. Interfering with disease: A progress report on siRNA-based therapeutics. Nat. Rev. Drug Discov. 2007, 6, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Moore, T.B.; Sakamoto, K.M. RNA interference and human disease. Mol. Genet. Metab. 2003, 80, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Dias, N.; Stein, C.A. Antisense oligonucleotides: Basic concepts and mechanisms. Mol. Cancer Ther. 2002, 1, 347–355. [Google Scholar] [PubMed]
- Bennett, C.F.; Swayze, E.E. RNA targeting therapeutics: Molecular mechanisms of antisense oligonucleotides as a therapeutic platform. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef] [PubMed]
- Van Oostrom, O.; Fledderus, J.O.; de Kleijn, D.; Pasterkamp, G.; Verhaar, M.C. Smooth muscle progenitor cells: Friend or foe in vascular disease? Curr. Stem. Cell Res. Ther. 2009, 4, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Mifune, M.; Sasamura, H.; Shimizu-Hirota, R.; Miyazaki, H.; Saruta, T. Angiotensin II type 2 receptors stimulate collagen synthesis in cultured vascular smooth muscle cells. Hypertension 2000, 36, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Nadler, J.L.; Natarajan, R.; Stern, N. Specific action of the lipoxygenase pathway in mediating angiotensin II-induced aldosterone synthesis in isolated adrenal glomerulosa cells. J. Clin. Invest. 1987, 80, 1763–1769. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.L.; Veerapanane, D.; Rossi, J.; Natarajan, R.; Thomas, L.; Nadler, J. Ribozyme-mediated inhibition of expression of leukocyte-type 12-lipoxygenase in porcine aortic vascular smooth muscle cells. Circ. Res. 1995, 77, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Sasson, S.; Eckel, J. Disparate effects of 12-lipoxygenase and 12-hydroxyeicosatetraenoic acid in vascular endothelial and smooth muscle cells and in cardiomyocytes. Arch. Physiol. Biochem. 2006, 112, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.S.; Fishbein, M.; Helfant, R.; Fagin, J. A paradigm for restenosis based on cell biology: Clues for the development of new preventive therapies. J. Am. Coll. Cardiol. 1991, 17, 758–769. [Google Scholar] [CrossRef]
- Jarvis, T.C.; Alby, L.J.; Beaudry, A.A.; Wincott, F.E.; Beigelman, L.; McSwiggen, J.A.; Usman, N.; Stinchcomb, D.T. Inhibition of vascular smooth muscle cell proliferation by ribozymes that cleave c-myb mRNA. RNA 1996, 2, 419–428. [Google Scholar] [PubMed]
- Dyson, N. The regulation of E2F by pRB-family proteins. Genes Dev. 1998, 12, 2245–2262. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, M.; Theodoras, A.M.; Schumacher, J.; Roberts, J.M.; Pagano, M. Human cyclin E, a nuclear protein essential for the G1-to-S phase transition. Mol. Cell Biol. 1995, 15, 2612–2624. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Schneider, A.; Engel, S.; Racchi, G.; Kandolf, R.; Kuhn, A. Hammerhead ribozymes targeted against cyclin E and E2F1 cooperate to down-regulate coronary smooth muscle cell proliferation. J. Gene Med. 2005, 7, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Mahadevan, M.; Tsilfidis, C.; Sabourin, L.; Shutler, G.; Amemiya, C.; Jansen, G.; Neville, C.; Narang, M.; Barcelo, J.; O’Hoy, K.; et al. Myotonic dystrophy mutation: An unstable CTG repeat in the 3’ untranslated region of the gene. Science 1992, 255, 1253–1255. [Google Scholar] [CrossRef] [PubMed]
- Brook, J.D.; McCurrach, M.E.; Harley, H.G.; Buckler, A.J.; Church, D.; Aburatani, H.; Hunter, K.; Stanton, V.P.; Thirion, J.P.; Hudson, T.; et al. Molecular basis of myotonic dystrophy: Expansion of a trinucleotide (CTG) repeat at the 3’ end of a transcript encoding a protein kinase family member. Cell 1992, 68, 799–808. [Google Scholar] [CrossRef]
- Mankodi, A.; Urbinati, C.R.; Yuan, Q.P.; Moxley, R.T.; Sansone, V.; Krym, M.; Henderson, D.; Schalling, M.; Swanson, M.S.; Thornton, C.A. Muscleblind localizes to nuclear foci of aberrant RNA in myotonic dystrophy types 1 and 2. Hum. Mol. Genet. 2001, 10, 2165–2170. [Google Scholar] [CrossRef] [PubMed]
- Timchenko, N.A.; Cai, Z.J.; Welm, A.L.; Reddy, S.; Ashizawa, T.; Timchenko, L.T. RNA CUG repeats sequester CUGBP1 and alter protein levels and activity of CUGBP1. J. Biol. Chem. 2001, 276, 7820–7826. [Google Scholar] [CrossRef] [PubMed]
- Langlois, M.A.; Lee, N.S.; Rossi, J.J.; Puymirat, J. Hammerhead ribozyme-mediated destruction of nuclear foci in myotonic dystrophy myoblasts. Mol. Ther. 2003, 7, 670–680. [Google Scholar] [CrossRef]
- Phylactou, L.A.; Darrah, C.; Wood, M.J. Ribozyme-mediated trans-splicing of a trinucleotide repeat. Nat. Genet. 1998, 18, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.S.; Vanoye, C.G.; Sullenger, B.A.; George, A.L., Jr. Functional repair of a mutant chloride channel using a trans-splicing ribozyme. J. Clin. Invest. 2002, 110, 1783–1789. [Google Scholar] [CrossRef] [PubMed]
- Croxen, R.; Newland, C.; Beeson, D.; Oosterhuis, H.; Chauplannaz, G.; Vincent, A.; Newsom-Davis, J. Mutations in different functional domains of the human muscle acetylcholine receptor alpha subunit in patients with the slow-channel congenital myasthenic syndrome. Hum. Mol. Genet. 1997, 6, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.J.; Mendes, Z.T.; Minatel, E.; Santo Neto, H. Acetylcholine receptors and nerve terminal distribution at the neuromuscular junction of long-term regenerated muscle fibers. J. Neurocytol. 2005, 34, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Abdelgany, A.; Ealing, J.; Wood, M.; Beeson, D. Selective cleavage of AChR cRNAs harbouring mutations underlying the slow channel myasthenic syndrome by hammerhead ribozymes. J. RNAi Gene Silencing 2005, 1, 26–31. [Google Scholar] [PubMed]
- Abdelgany, A.; Ealing, J.; Wood, M.; Beeson, D. Selective DNAzyme-mediated cleavage of AChR mutant transcripts by targeting the mutation site or through mismatches in the binding arm. J. RNAi Gene Silencing 2005, 1, 32–37. [Google Scholar] [PubMed]
- Kremer, B.; Goldberg, P.; Andrew, S.E.; Theilmann, J.; Telenius, H.; Zeisler, J.; Squitieri, F.; Lin, B.; Bassett, A.; Almqvist, E.; et al. A worldwide study of the Huntington’s disease mutation. The sensitivity and specificity of measuring CAG repeats. N. Engl. J. Med. 1994, 330, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Yen, L.; Strittmatter, S.M.; Kalb, R.G. Sequence-specific cleavage of Huntingtin mRNA by catalytic DNA. Ann. Neurol. 1999, 46, 366–373. [Google Scholar] [CrossRef]
- Wang, Y.L.; Liu, W.; Wada, E.; Murata, M.; Wada, K.; Kanazawa, I. Clinico-pathological rescue of a model mouse of Huntington’s disease by siRNA. Neurosci. Res. 2005, 53, 241–249. [Google Scholar] [CrossRef] [PubMed]
- DiFiglia, M.; Sena-Esteves, M.; Chase, K.; Sapp, E.; Pfister, E.; Sass, M.; Yoder, J.; Reeves, P.; Pandey, R.K.; Rajeev, K.G.; Manoharan, M.; Sah, D.W.; Zamore, P.D.; Aronin, N. Therapeutic silencing of mutant huntingtin with siRNA attenuates striatal and cortical neuropathology and behavioral deficits. Proc. Natl. Acad. Sci. USA 2007, 104, 17204–17209. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s disease. In the beginning. Nature 1991, 354, 432–433. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, B.; Antoszczyk, S.; Maszewska, M.; Kuwabara, T.; Warashina, M.; Taira, K.; Stec, W.J. Efficient inhibition of beta-secretase gene expression in HEK293 cells by tRNAVal-driven and CTE-helicase associated hammerhead ribozymes. Eur. J. Biochem. 2003, 270, 3962–3970. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, T.; Nishimaki, K.; Asoh, S.; Ishibashi, Y.; Takata, I.; Kuwabara, T.; Taira, K.; Yamaguchi, H.; Sugihara, S.; Yamazaki, T.; Ihara, Y.; Nakano, K.; Matuda, S.; Ohta, S. Truncated product of the bifunctional DLST gene involved in biogenesis of the respiratory chain. EMBO J. 2003, 22, 2913–2923. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S.; Ohsawa, I. Dysfunction of mitochondria and oxidative stress in the pathogenesis of Alzheimer’s disease: On defects in the cytochrome c oxidase complex and aldehyde detoxification. J. Alzheimers Dis. 2006, 9, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Onuki, R.; Bando, Y.; Suyama, E.; Katayama, T.; Kawasaki, H.; Baba, T.; Tohyama, M.; Taira, K. An RNA-dependent protein kinase is involved in tunicamycin-induced apoptosis and Alzheimer’s disease. EMBO J. 2004, 23, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Steece-Collier, K.; Maries, E.; Kordower, J.H. Etiology of Parkinson’s disease: Genetics and environment revisited. Proc. Natl. Acad. Sci. USA 2002, 99, 13972–13974. [Google Scholar] [CrossRef] [PubMed]
- Maries, E.; Dass, B.; Collier, T.J.; Kordower, J.H.; Steece-Collier, K. The role of alpha-synuclein in Parkinson’s disease: Insights from animal models. Nat. Rev. Neurosci. 2003, 4, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Hayashita-Kinoh, H.; Yamada, M.; Yokota, T.; Mizuno, Y.; Mochizuki, H. Down-regulation of alpha-synuclein expression can rescue dopaminergic cells from cell death in the substantia nigra of Parkinson’s disease rat model. Biochem. Biophys. Res. Commun. 2006, 341, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Kowall, N.W.; Hantraye, P.; Brouillet, E.; Beal, M.F.; McKee, A.C.; Ferrante, R.J. MPTP induces alpha-synuclein aggregation in the substantia nigra of baboons. Neuroreport 2000, 11, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Grzelinski, M.; Bader, N.; Czubayko, F.; Aigner, A. Ribozyme-targeting reveals the rate-limiting role of pleiotrophin in glioblastoma. Int. J. Cancer 2005, 117, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Benson, V.L.; Khachigian, L.M.; Lowe, H.C. DNAzymes and cardiovascular disease. Br. J. Pharmacol. 2008, 154, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Czubayko, F.; Riegel, A.T.; Wellstein, A. Ribozyme-targeting elucidates a direct role of pleiotrophin in tumor growth. J. Biol. Chem. 1994, 269, 21358–21363. [Google Scholar] [PubMed]
- Powers, C.; Aigner, A.; Stoica, G.E.; McDonnell, K.; Wellstein, A. Pleiotrophin signaling through anaplastic lymphoma kinase is rate-limiting for glioblastoma growth. J. Biol. Chem. 2002, 277, 14153–14158. [Google Scholar] [CrossRef] [PubMed]
- Bowden, E.T.; Stoica, G.E.; Wellstein, A. Anti-apoptotic signaling of pleiotrophin through its receptor, anaplastic lymphoma kinase. J. Biol. Chem. 2002, 277, 35862–35868. [Google Scholar] [CrossRef] [PubMed]
- Grzelinski, M.; Steinberg, F.; Martens, T.; Czubayko, F.; Lamszus, K.; Aigner, A. Enhanced antitumorigenic effects in glioblastoma on double targeting of pleiotrophin and its receptor ALK. Neoplasia 2009, 11, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Laaroubi, K.; Delbe, J.; Vacherot, F.; Desgranges, P.; Tardieu, M.; Jaye, M.; Barritault, D.; Courty, J. Mitogenic and in vitro angiogenic activity of human recombinant heparin affin regulatory peptide. Growth Factors 1994, 10, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Czubayko, F.; Schulte, A.M.; Berchem, G.J.; Wellstein, A. Melanoma angiogenesis and metastasis modulated by ribozyme targeting of the secreted growth factor pleiotrophin. Proc. Natl. Acad. Sci. USA 1996, 93, 14753–14758. [Google Scholar] [CrossRef] [PubMed]
- Vrati, S.; Giri, R.K.; Razdan, A.; Malik, P. Complete nucleotide sequence of an Indian strain of Japanese encephalitis virus: Sequence comparison with other strains and phylogenetic analysis. Am. J. Trop. Med. Hyg. 1999, 61, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Santoro, S.W.; Joyce, G.F. A general purpose RNA-cleaving DNA enzyme. Proc. Natl. Acad. Sci. USA 1997, 94, 4262–4266. [Google Scholar] [CrossRef] [PubMed]
- Appaiahgari, M.B.; Vrati, S. DNAzyme-mediated inhibition of Japanese encephalitis virus replication in mouse brain. Mol. Ther. 2007, 15, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available |


{kind=link}
| Ribozyme-DNAzyme | Disease | Gene-Gene target | Reference |
|---|---|---|---|
| Hammerhead ribozyme | Vascular smooth muscle cell hypertrophy-Restenosis | Leukocyte-type 12-lipoxygenase (12-LO) | Gu, J.L., et.al. Circ. Res. 1995, 77, 14-20 |
| Hammerhead ribozyme | Vascular smooth muscle cell hypertrophy-Restenosis | Proto-oncogene c-myb | Jarvis, T.C., et al. RNA, 1996, 2, 419-428. |
| Hammerhead ribozyme | Vascular smooth muscle cell hypertrophy-Restenosis | Cyclin E and Cyclin E2F1 | Grassi, G., et al., J Gene Med, 2005. 7(9): p. 1223-1234. |
| Hammerhead ribozyme | Myotonic Dystrophy type 1 (DM1) | Mutant DMPK 3’ UTR | Langlois, M.A., et al., Mol Ther, 2003. 7(5 Pt 1): p. 670-680. |
| Group I intron ribozyme | Myotonic Dystrophy type 1 (DM1) | Mutant DMPK 3’ UTR | Phylactou, L.A., et al., Nat Genet, 1998. 18(4): p. 378-381. |
| Hammerhead ribozyme | Slow channel congenital myasthenic syndrome | Acetylcholine receptor (AchR) | Abdelgany, A., et al., J RNAi Gene Silencing, 2005. 1(1): p. 26-31. |
| DNAzyme | Slow channel congenital myasthenic syndrome | Acetylcholine receptor (AchR) | Abdelgany, A., et al., J RNAi Gene Silencing, 2005. 1(1): p. 32-37. |
| DNAzyme | Huntington’s | Huntingtin (HTT) | Yen, L., et al., Ann Neurol, 1999. 46(3): p. 366-373. |
| Hammerhead ribozyme | Alzheimer’s | β-Secretase | Nawrot, B., et al., Eur J Biochem, 2003. 270(19): p. 3962-3970. |
| Hammerhead ribozyme | Alzheimer’s | Dihydrolipoamide succinyltransferase (DLST) | Kanamori, T., et al., EMBO J, 2003. 22(12): p. 2913-2923. |
| Hammerhead ribozyme | Parkinson’s | a-Synuclein | Hayashita-Kinoh, H., et al., Biochem Biophys Res Commun, 2006. 341(4): p. 1088-1095. |
| Hammerhead ribozyme | Glioblastomas | Pleiotrophin (PTN) | Grzelinski, M., et al., Int J Cancer, 2005. 117(6): p. 942-951. |
| Hammerhead ribozyme | Glioblastomas | Pleiotrophin (PTN) and anaplastic lymphoma kinase (ALK) | Grzelinski, M., et al., Neoplasia, 2009. 11(2): p. 145-156. |
| Hammerhead ribozyme | Encephalitis | Japanese encephalitis virus (JEV) | Appaiahgari, M.B. and S. Vrati. Mol Ther, 2007. 15(9): p. 1593-1599. |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mastroyiannopoulos, N.P.; Uney, J.B.; Phylactou, L.A. The Application of Ribozymes and DNAzymes in Muscle and Brain. Molecules 2010, 15, 5460-5472. https://doi.org/10.3390/molecules15085460
Mastroyiannopoulos NP, Uney JB, Phylactou LA. The Application of Ribozymes and DNAzymes in Muscle and Brain. Molecules. 2010; 15(8):5460-5472. https://doi.org/10.3390/molecules15085460
Chicago/Turabian StyleMastroyiannopoulos, Nikolaos P., James B. Uney, and Leonidas A. Phylactou. 2010. "The Application of Ribozymes and DNAzymes in Muscle and Brain" Molecules 15, no. 8: 5460-5472. https://doi.org/10.3390/molecules15085460