Locked and Unlocked Nucleosides in Functional Nucleic Acids

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
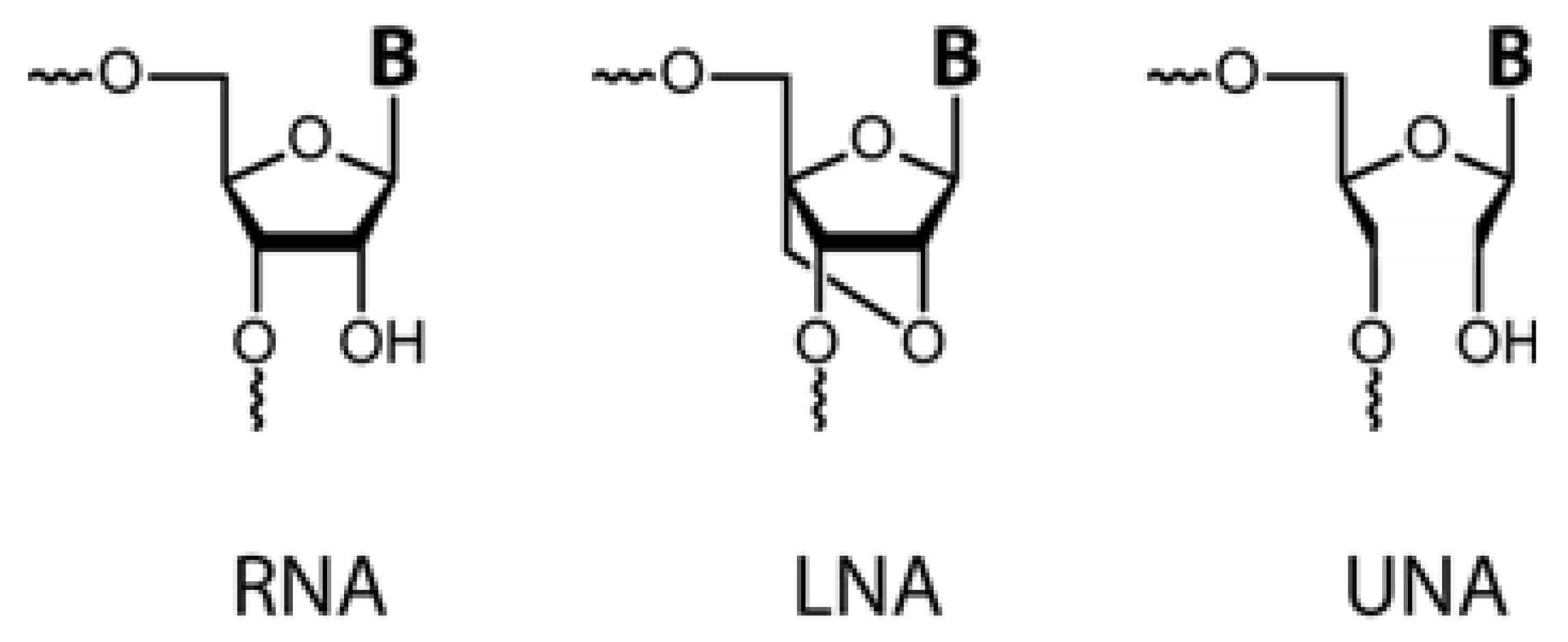
2. Locked Nucleic Acids
Structure and Properties
3. Functional Nucleic Acids with LNA
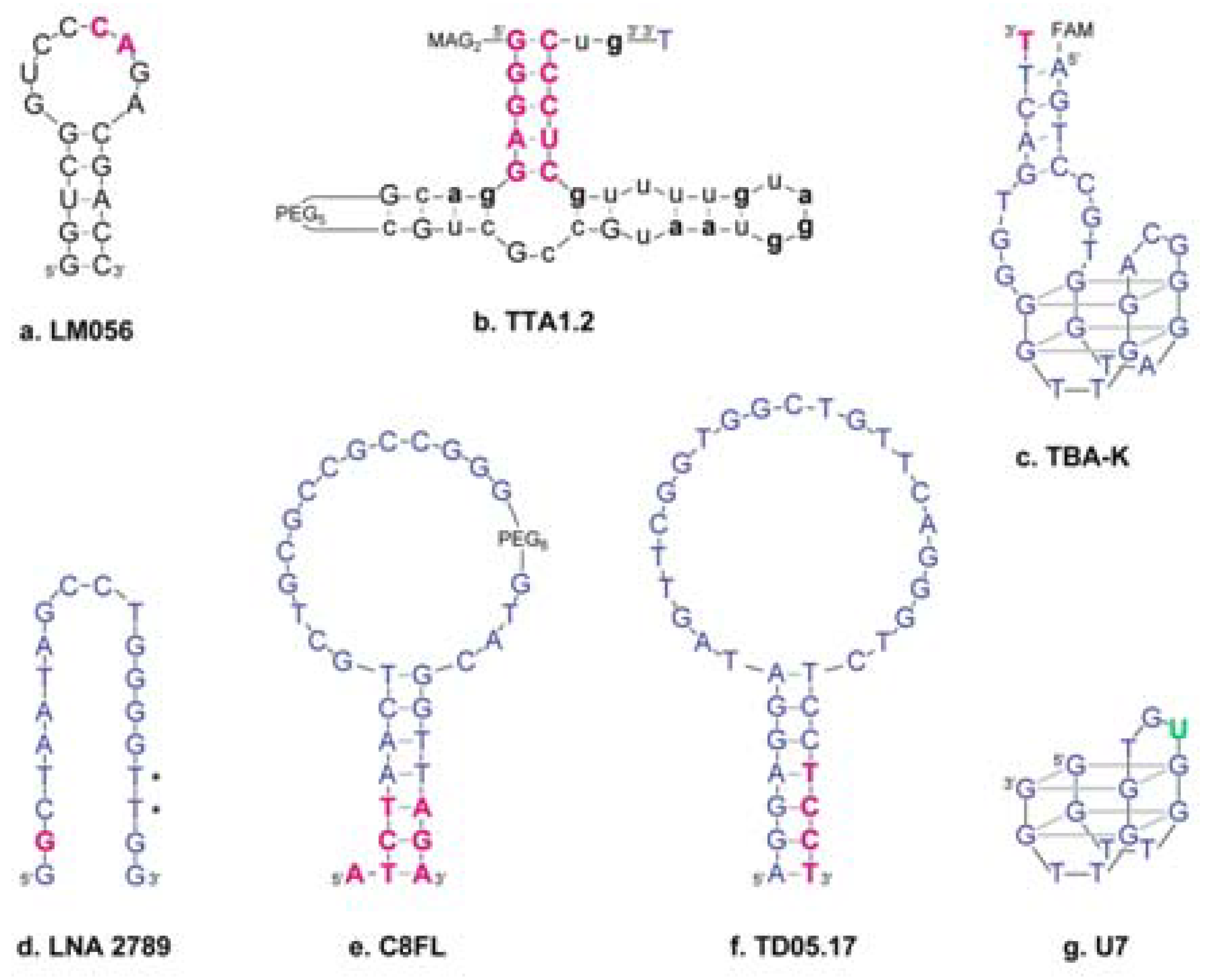
3.1. LNA in Aptamers
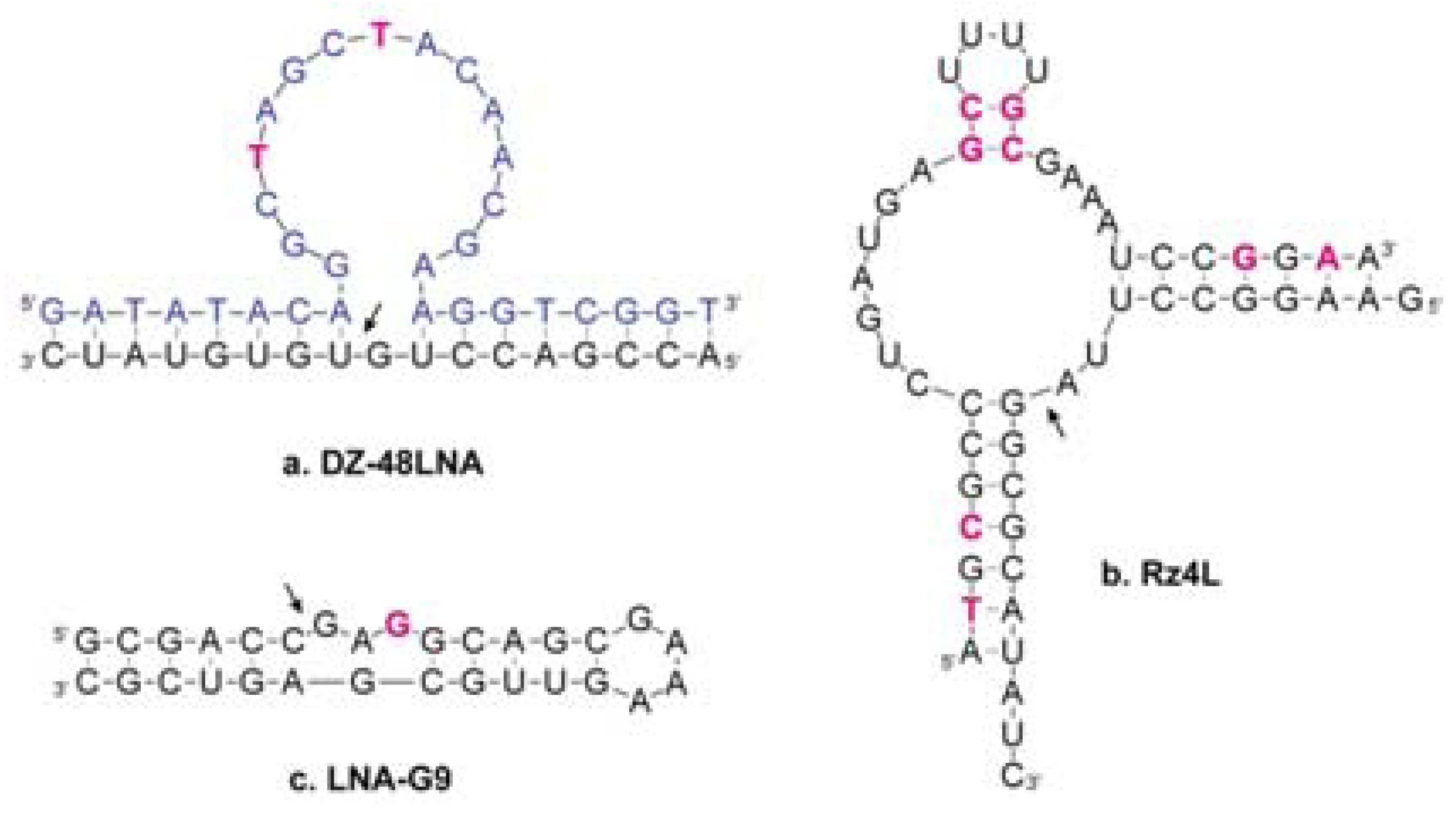
3.2. LNA in Catalytic Nucleic Acids
4. Unlocked Nucleic Acids
4.1. Properties
4.2. UNA in Aptamers
5. De Novo Selection of Modified Nucleic Acid Structures
6. Conclusions
Acknowledgments
References
- Hermann, T.; Patel, D.J. Stitching together RNA tertiary architectures. J. Mol. Biol. 1999, 294, 829–849. [Google Scholar] [CrossRef] [PubMed]
- Nagaswamy, U.; Voss, N.; Zhang, Z.; Fox, G.E. Database of non-canonical base pairs found in known RNA structures. Nucleic Acids Res. 2000, 28, 375–376. [Google Scholar] [CrossRef] [PubMed]
- Koshkin, A.A.; Singh, S.K.; Nielsen, P.; Rajwanshi, V.K.; Kumar, R.; Meldgaard, M.; Olsen, C.E.; Wengel, J. LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition. Tetrahedron 1998, 54, 3607–3630. [Google Scholar] [CrossRef]
- Singh, S.K.; Nielsen, P.; Koshkin, A.; Wengel, J. LNA (locked nucleic acids): Synthesis and high-affinity nucleic acid recognition. Chem. Commun. 1998, 4, 455–456. [Google Scholar] [CrossRef]
- Obika, S.; Nanbu, D.; Hari, Y.; Morio, K.-I.; In, Y.; Ishida, T.; Imanishi, T. Synthesis of 2’-O,4’-C-methyleneuridine and -cytidine. Novel bicyclic nucleosides having a fixed C3’-endo sugar puckering. Tetrahedron Lett. 1997, 38, 8735–8738. [Google Scholar] [CrossRef]
- Langkjaer, N.; Pasternak, A.; Wengel, J. UNA (unlocked nucleic acid): A flexible RNA mimic that allows engineering of nucleic acid duplex stability. Bioorg. Med. Chem. 2009, 17, 5420–5425. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.; Dreioe, L.H.; Wengel, J. Synthesis and evaluation of oligodeoxynucleotides containing acyclic nucleosides: Introduction of three novel analogues and a summary. Bioorg. Med. Chem. 1995, 3, 19–28. [Google Scholar] [CrossRef]
- Vester, B.; Wengel, J. LNA (locked nucleic acid): High-affinity targeting of complementary RNA and DNA. Biochemistry 2004, 43, 13233–13241. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.E.; Rasmussen, J.; Kumar, R.; Wengel, J.; Jacobsen, J.P.; Petersen, M. NMR studies of fully modified locked nucleic acid (LNA) hybrids: Solution structure of an LNA:RNA hybrid and characterization of an LNA:DNA hybrid. Bioconjug. Chem. 2004, 15, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Nielsen, C.B.; Nielsen, K.E.; Jensen, G.A.; Bondensgaard, K.; Singh, S.K.; Rajwanshi, V.K.; Koshkin, A.A.; Dahl, B.M.; Wengel, J.; et al. The conformations of locked nucleic acids (LNA). J. Mol. Recognit. 2000, 13, 44–53. [Google Scholar] [CrossRef]
- Obika, S.; Nanbu, D.; Hari, Y.; Andoh, J.; Morio, K.; Doi, T.; Imanishi, T. Stability and structural features of the duplexes containing nucleoside analogues with a fixed N-type conformation, 2’-O,4’-C-methyleneribonucleosides. Tetrahedron Lett. 1998, 39, 5401–5404. [Google Scholar] [CrossRef]
- Wengel, J. Synthesis of 3’-C- and 4’-C-branched oligodeoxynucleotides and the development of locked nucleic acid (LNA). Acc. Chem. Res. 1999, 32, 301–310. [Google Scholar] [CrossRef]
- Braasch, D.A.; Corey, D.R. Locked nucleic acid (LNA): Fine-tuning the recognition of DNA and RNA. Chem. Biol. 2001, 8, 1–7. [Google Scholar] [CrossRef]
- Kurreck, J.; Wyszko, E.; Gillen, C.; Erdmann, V.A. Design of antisense oligonucleotides stabilized by locked nucleic acids. Nucleic Acids Res. 2002, 30, 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Bondensgaard, K.; Petersen, M.; Singh, S.K.; Rajwanshi, V.K.; Kumar, R.; Wengel, J.; Jacobsen, J.P. Structural studies of LNA:RNA duplexes by NMR: Conformations and implications for RNase H activity. Chemistry 2000, 6, 2687–2695. [Google Scholar] [CrossRef]
- Jepsen, J.S.; Wengel, J. LNA-antisense rivals siRNA for gene silencing. Curr. Opin. Drug Discov. Devel. 2004, 7, 188–194. [Google Scholar] [PubMed]
- Roberts, J.; Palma, E.; Sazani, P.; Orum, H.; Cho, M.; Kole, R. Efficient and persistent splice switching by systemically delivered LNA oligonucleotides in mice. Mol. Ther. 2006, 14, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Graziewicz, M.A.; Tarrant, T.K.; Buckley, B.; Roberts, J.; Fulton, L.; Hansen, H.; Orum, H.; Kole, R.; Sazani, P. An endogenous TNF-alpha antagonist induced by splice-switching oligonucleotides reduces inflammation in hepatitis and arthritis mouse models. Mol. Ther. 2008, 16, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Guterstam, P.; Lindgren, M.; Johansson, H.; Tedebark, U.; Wengel, J.; El Andaloussi, S.; Langel, U. Splice-switching efficiency and specificity for oligonucleotides with locked nucleic acid monomers. Biochem. J. 2008, 412, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Childs, J.L.; Disney, M.D.; Turner, D.H. Oligonucleotide directed misfolding of RNA inhibits Candida albicans group I intron splicing. Proc. Natl. Acad. Sci. USA 2002, 99, 11091–11096. [Google Scholar] [CrossRef] [PubMed]
- Fluiter, K.; Mook, O.R.; Baas, F. The therapeutic potential of LNA-modified siRNAs: Reduction of off-target effects by chemical modification of the siRNA sequence. Methods Mol. Biol. 2009, 487, 189–203. [Google Scholar] [PubMed]
- Wang, L.; Yang, C.J.; Medley, C.D.; Benner, S.A.; Tan, W. Locked nucleic acid molecular beacons. J. Am. Chem. Soc. 2005, 127, 15664–15665. [Google Scholar] [CrossRef] [PubMed]
- Buh Gasparic, M.; Tengs, T.; La Paz, J.L.; Holst-Jensen, A.; Pla, M.; Esteve, T.; Zel, J.; Gruden, K. Comparison of nine different real-time PCR chemistries for qualitative and quantitative applications in GMO detection. Anal. Bioanal. Chem. 2010, 396, 2023–2029. [Google Scholar] [CrossRef] [PubMed]
- Martinez, K.; Estevez, M.C.; Wu, Y.; Phillips, J.A.; Medley, C.D.; Tan, W. Locked nucleic acid based beacons for surface interaction studies and biosensor development. Anal. Chem. 2009, 81, 3448–3454. [Google Scholar] [CrossRef] [PubMed]
- Morandi, L.; Ferrari, D.; Lombardo, C.; Pession, A.; Tallini, G. Monitoring HCV RNA viral load by locked nucleic acid molecular beacons real time PCR. J. Virol. Methods 2007, 140, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Stenvang, J.; Silahtaroglu, A.N.; Lindow, M.; Elmen, J.; Kauppinen, S. The utility of LNA in microRNA-based cancer diagnostics and therapeutics. Semin. Cancer Biol. 2008, 18, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Vester, B.; Lundberg, L.B.; Sorensen, M.D.; Babu, B.R.; Douthwaite, S.; Wengel, J. LNAzymes: Incorporation of LNA-type monomers into DNAzymes markedly increases RNA cleavage. J. Am. Chem. Soc. 2002, 124, 13682–13683. [Google Scholar] [CrossRef] [PubMed]
- Donini, S.; Clerici, M.; Wengel, J.; Vester, B.; Peracchi, A. The advantages of being locked. ASSESSING the cleavage of short and long RNAs by locked nucleic acid-containing 8–17 deoxyribozymes. J. Biol. Chem. 2007, 282, 35510–35518. [Google Scholar] [CrossRef] [PubMed]
- Fluiter, K.; Frieden, M.; Vreijling, J.; Koch, T.; Baas, F. Evaluation of LNA-modified DNAzymes targeting a single nucleotide polymorphism in the large subunit of RNA polymerase II. Oligonucleotides 2005, 15, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, R.G.; Khachigian, L.M. Locked nucleic acid modified DNA enzymes targeting early growth response-1 inhibit human vascular smooth muscle cell growth. Nucleic Acids Res. 2004, 32, 2281–2285. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, M.R.; Haasnoot, J.; Wengel, J.; Berkhout, B.; Kjems, J. Efficient inhibition of HIV-1 expression by LNA modified antisense oligonucleotides and DNAzymes targeted to functionally selected binding sites. Retrovirology 2007, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Scaria, V.; Maiti, S. “Locked onto the Target”: Increasing the efficiency of antagomirzymes using locked nucleic acid modifications. Biochemistry 2010, 49, 9449–9456. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Furste, J.P.; Werk, D.; Grunert, H.P.; Zeichhardt, H.; Erdmann, V.A.; Kurreck, J. Gaining target access for deoxyribozymes. J. Mol. Biol. 2004, 339, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Vester, B.; Hansen, L.H.; Lundberg, L.B.; Babu, B.R.; Sorensen, M.D.; Wengel, J.; Douthwaite, S. Locked nucleoside analogues expand the potential of DNAzymes to cleave structured RNA targets. BMC Mol. Biol. 2006, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Vester, B.; Lundberg, L.B.; Sorensen, M.D.; Babu, B.R.; Douthwaite, S.; Wengel, J. Improved RNA cleavage by LNAzyme derivatives of DNAzymes. Biochem. Soc. Trans. 2004, 32, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Gul, D.C.; Grunert, H.P.; Zeichhardt, H.; Erdmann, V.A.; Kurreck, J. RNA cleaving ‘10–23’ DNAzymes with enhanced stability and activity. Nucleic Acids Res. 2003, 31, 5982–5992. [Google Scholar] [CrossRef] [PubMed]
- Abdelgany, A.; Uddin, M.K.; Wood, M.; Taira, K.; Beeson, D. Design of efficient DNAzymes against muscle AChR alpha-subunit cRNA in vitro and in HEK 293 cells. J. RNAi Gene Silencing 2005, 1, 88–96. [Google Scholar] [PubMed]
- Wahlestedt, C.; Salmi, P.; Good, L.; Kela, J.; Johnsson, T.; Hokfelt, T.; Broberger, C.; Porreca, F.; Lai, J.; Ren, K.; et al. Potent and nontoxic antisense oligonucleotides containing locked nucleic acids. Proc. Natl. Acad. Sci. USA 2000, 97, 5633–5638. [Google Scholar] [CrossRef] [PubMed]
- Straarup, E.M.; Fisker, N.; Hedtjarn, M.; Lindholm, M.W.; Rosenbohm, C.; Aarup, V.; Hansen, H.F.; Orum, H.; Hansen, J.B.; Koch, T. Short locked nucleic acid antisense oligonucleotides potently reduce apolipoprotein B mRNA and serum cholesterol in mice and non-human primates. Nucleic Acids Res. 2010, 38, 7100–7111. [Google Scholar] [CrossRef] [PubMed]
- Fluiter, K.; ten Asbroek, A.L.; de Wissel, M.B.; Jakobs, M.E.; Wissenbach, M.; Olsson, H.; Olsen, O.; Oerum, H.; Baas, F. In vivo tumor growth inhibition and biodistribution studies of locked nucleic acid (LNA) antisense oligonucleotides. Nucleic Acids Res. 2003, 31, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Elmen, J.; Lindow, M.; Silahtaroglu, A.; Bak, M.; Christensen, M.; Lind-Thomsen, A.; Hedtjarn, M.; Hansen, J.B.; Hansen, H.F.; Straarup, E.M.; et al. Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Res. 2008, 36, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.A.; Hansen, J.B.; Lai, J.; Wu, S.; Voskresenskiy, A.; Hog, A.; Worm, J.; Hedtjarn, M.; Souleimanian, N.; Miller, P.; et al. Efficient gene silencing by delivery of locked nucleic acid antisense oligonucleotides, unassisted by transfection reagents. Nucleic Acids Res. 2010, 38, e3. [Google Scholar] [CrossRef] [PubMed]
- Swayze, E.E.; Siwkowski, A.M.; Wancewicz, E.V.; Migawa, M.T.; Wyrzykiewicz, T.K.; Hung, G.; Monia, B.P.; Bennett, C.F. Antisense oligonucleotides containing locked nucleic acid improve potency but cause significant hepatotoxicity in animals. Nucleic Acids Res. 2007, 35, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Santaris Pharma A/S. Santaris Pharma A/S advances miravirsen, the first microRNA-targeted drug to enter clinical trials, into Phase 2 to treat patients infected with Hepatitis C virus. Available online: http://www.santaris.com/news/2010/09/23/santaris-pharma-advances-miravirsen-first-microrna-targeted-drug-enter-clinical-tria/ (accessed on: May 16, 2011).
- Santaris Pharma A/S. Another LNA-based RNA Inhibitor Enters Clinical Trials. Available online: http://www.santaris.com/news/2009/03/20/another-lna-based-rna-inhibitor-enters-clinical-trials/ (accessed on: May 16, 2011).
- Elmen, J.; Thonberg, H.; Ljungberg, K.; Frieden, M.; Westergaard, M.; Xu, Y.; Wahren, B.; Liang, Z.; Orum, H.; Koch, T.; Wahlestedt, C. Locked nucleic acid (LNA) mediated improvements in siRNA stability and functionality. Nucleic Acids Res. 2005, 33, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Dagnaes-Hansen, F.; Nielsen, E.J.; Wengel, J.; Besenbacher, F.; Howard, K.A.; Kjems, J. The effect of chemical modification and nanoparticle formulation on stability and biodistribution of siRNA in mice. Mol. Ther. 2009, 17, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Mook, O.R.; Baas, F.; de Wissel, M.B.; Fluiter, K. Evaluation of locked nucleic acid-modified small interfering RNA in vitro and in vivo. Mol. Cancer Ther. 2007, 6, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Frieden, M.; Christensen, S.M.; Mikkelsen, N.D.; Rosenbohm, C.; Thrue, C.A.; Westergaard, M.; Hansen, H.F.; Orum, H.; Koch, T. Expanding the design horizon of antisense oligonucleotides with alpha-L-LNA. Nucleic Acids Res. 2003, 31, 6365–6372. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, K.; Veedu, R.N.; Wengel, J. Nuclease resistant methylphosphonate-DNA/LNA chimeric oligonucleotides. Bioorg. Med. Chem. Lett. 2009, 19, 2707–2709. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Takagi, M.; Hasegawa, C.; Kaneko, M.; Tsutsumi, S.; Sone, J.; Ishikawa, T.; Imanishi, T.; Koizumi, M. Synthesis and properties of 2’-O,4’-C-ethylene-bridged nucleic acids (ENA) as effective antisense oligonucleotides. Bioorg. Med. Chem. 2003, 11, 2211–2226. [Google Scholar] [CrossRef]
- Crinelli, R.; Bianchi, M.; Gentilini, L.; Magnani, M. Design and characterization of decoy oligonucleotides containing locked nucleic acids. Nucleic Acids Res. 2002, 30, 2435–2443. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. SELEX-A (r)evolutionary method to generate high-affinity nucleic acid ligands. Biomol. Eng. 2007, 4, 381–403. [Google Scholar] [CrossRef] [PubMed]
- Bunka, D.H.; Stockley, P.G. Aptamers come of age—At last. Nat. Rev. Microbiol. 2006, 4, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Syed, M.A.; Pervaiz, S. Advances in aptamers. Oligonucleotides 2010, 20, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Lee, J.W.; Ellington, A.D. Applications of aptamers as sensors. Annu. Rev. Anal. Chem. (Palo Alto Calif) 2009, 2, 241–264. [Google Scholar] [CrossRef] [PubMed]
- Duconge, F.; Toulme, J.J. In vitro selection identifies key determinants for loop-loop interactions: RNA aptamers selective for the TAR RNA element of HIV-1. RNA 1999, 5, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Darfeuille, F.; Hansen, J.B.; Orum, H.; Di Primo, C.; Toulme, J.J. LNA/DNA chimeric oligomers mimic RNA aptamers targeted to the TAR RNA element of HIV-1. Nucleic Acids Res. 2004, 32, 3101–3107. [Google Scholar] [CrossRef] [PubMed]
- Di Primo, C.; Rudloff, I.; Reigadas, S.; Arzumanov, A.A.; Gait, M.J.; Toulme, J.J. Systematic screening of LNA/2’-O-methyl chimeric derivatives of a TAR RNA aptamer. FEBS Lett. 2007, 581, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Hicke, B.J.; Marion, C.; Chang, Y.F.; Gould, T.; Lynott, C.K.; Parma, D.; Schmidt, P.G.; Warren, S. Tenascin-C aptamers are generated using tumor cells and purified protein. J. Biol. Chem. 2001, 276, 48644–48654. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.S.; Borkowski, S.; Kurreck, J.; Stephens, A.W.; Bald, R.; Hecht, M.; Friebe, M.; Dinkelborg, L.; Erdmann, V.A. Application of locked nucleic acids to improve aptamer in vivo stability and targeting function. Nucleic Acids Res. 2004, 32, 5757–5765. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, Y.; Kitadume, S.; Morihiro, K.; Kuwahara, M.; Ozaki, H.; Sawai, H.; Imanishi, T.; Obika, S. Effect of 3’-end capping of aptamer with various 2’,4’-bridged nucleotides: Enzymatic post-modification toward a practical use of polyclonal aptamers. Bioorg. Med. Chem. Lett. 2010, 20, 1626–1629. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, F.J.; Kalra, N.; Wengel, J.; Vester, B. Aptamers as a model for functional evaluation of LNA and 2’-amino LNA. Bioorg. Med. Chem. Lett. 2009, 19, 6585–6587. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, D.; Tang, Z.; Mallikaratchy, P.; Xiao, Z.; Tan, W. Optimization and modifications of aptamers selected from live cancer cell lines. Chembiochem 2007, 8, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Mallikaratchy, P.R.; Ruggiero, A.; Gardner, J.R.; Kuryavyi, V.; Maguire, W.F.; Heaney, M.L.; McDevitt, M.R.; Patel, D.J.; Scheinberg, D.A. A multivalent DNA aptamer specific for the B-cell receptor on human lymphoma and leukemia. Nucleic Acids Res. 2010, 39, 2458–2469. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, A.; Hernandez, F.J.; Rasmussen, L.M.; Vester, B.; Wengel, J. Improved thrombin binding aptamer by incorporation of a single unlocked nucleic acid monomer. Nucleic Acids Res. 2011, 39, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Bock, L.C.; Griffin, L.C.; Latham, J.A.; Vermaas, E.H.; Toole, J.J. Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature 1992, 355, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Virno, A.; Randazzo, A.; Giancola, C.; Bucci, M.; Cirino, G.; Mayol, L. A novel thrombin binding aptamer containing a G-LNA residue. Bioorg. Med. Chem. 2007, 15, 5710–5718. [Google Scholar] [CrossRef] [PubMed]
- Bonifacio, L.; Church, F.C.; Jarstfer, M.B. Effect of locked-nucleic acid on a biologically active g-quadruplex. A structure-activity relationship of the thrombin aptamer. Int. J. Mol. Sci. 2008, 9, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Tasset, D.M.; Kubik, M.F.; Steiner, W. Oligonucleotide inhibitors of human thrombin that bind distinct epitopes. J. Mol. Biol. 1997, 272, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, F.J.; Dondapati, S.K.; Ozalp, V.C.; Pinto, A.; O’Sullivan, C.K.; Klar, T.A.; Katakis, I. Label free optical sensor for Avidin based on single gold nanoparticles functionalized with aptamers. J. Biophotonics 2009, 2, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Mallikaratchy, P.; Tang, Z.; Kwame, S.; Meng, L.; Shangguan, D.; Tan, W. Aptamer directly evolved from live cells recognizes membrane bound immunoglobin heavy mu chain in Burkitt’s lymphoma cells. Mol. Cell. Proteomics 2007, 6, 2230–2238. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Shangguan, D.; Wang, K.; Shi, H.; Sefah, K.; Mallikratchy, P.; Chen, H.W.; Li, Y.; Tan, W. Selection of aptamers for molecular recognition and characterization of cancer cells. Anal. Chem. 2007, 79, 4900–4907. [Google Scholar] [CrossRef] [PubMed]
- Zaug, A.J.; Cech, T.R. In vitro splicing of the ribosomal RNA precursor in nuclei of Tetrahymena. Cell 1980, 19, 331–338. [Google Scholar] [CrossRef]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Weigand, B.-S.; Zerressen, A.; Schlatterer, J.C.; Helm, M.; Jaschke, A. In vitro selection of short, catalytically active oligonucleotides. In The Aptamer Handbook: Functional Oligonucleotides and Their Applications; Klussmann, S., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2006; Volume 1, pp. 211–227. [Google Scholar]
- Pan, W.; Clawson, G.A. Catalytic DNAzymes: Derivations and functions. Expert Opin. Biol. Ther. 2008, 8, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Santoro, S.W.; Joyce, G.F. A general purpose RNA-cleaving DNA enzyme. Proc. Natl. Acad. Sci. USA 1997, 94, 4262–4266. [Google Scholar] [CrossRef] [PubMed]
- Robaldo, L.; Montserrat, J.M.; Iribarren, A.M. 10-23 DNAzyme modified with (2’R)- and (2’S)-2’-deoxy-2’-C-methyluridine in the catalytic core. Bioorg. Med. Chem. Lett. 2010, 20, 4367–4370. [Google Scholar] [CrossRef] [PubMed]
- Lobedanz, S.; Vester, B. University of Southern Denmark, Campusvej 55 Odense M 5230. Denmark, 2011. [Google Scholar]
- Christiansen, J.K.; Lobedanz, S.; Arar, K.; Wengel, J.; Vester, B. LNA nucleotides improve cleavage efficiency of singular and binary hammerhead ribozymes. Bioorg. Med. Chem. 2007, 15, 6135–6143. [Google Scholar] [CrossRef] [PubMed]
- Persson, T.; Hartmann, R.K.; Eckstein, F. Selection of hammerhead ribozyme variants with low Mg2+ requirement: Importance of stem-loop II. Chembiochem 2002, 3, 1066–1071. [Google Scholar] [CrossRef]
- Fedoruk-Wyszomirska, A.; Szymanski, M.; Wyszko, E.; Barciszewska, M.Z.; Barciszewski, J. Highly active low magnesium hammerhead ribozyme. J. Biochem. 2009, 145, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Uhlenbeck, O.C. A small metalloribozyme with a two-step mechanism. Nature 1992, 358, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Dichtl, B.; Uhlenbeck, O.C. Properties of an in vitro selected Pb2+ cleavage motif. Biochemistry 1994, 33, 9561–9565. [Google Scholar] [CrossRef] [PubMed]
- Julien, K.R.; Sumita, M.; Chen, P.H.; Laird-Offringa, I.A.; Hoogstraten, C.G. Conformationally restricted nucleotides as a probe of structure-function relationships in RNA. RNA 2008, 14, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, A.; Wengel, J. Unlocked nucleic acid—An RNA modification with broad potential. Org. Biomol. Chem. 2011, 9, 3591–3597. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.B.; Langkjaer, N.; Wengel, J. Unlocked nucleic acid (UNA) and UNA derivatives: Thermal denaturation studies. Nucleic Acids Symp. Ser. (Oxf) 2008, 133–134. [Google Scholar] [CrossRef] [PubMed]
- Itkes, A.V.; Karpeisky, M.; Kartasheva, O.N.; Mikhailov, S.N.; Moiseyev, G.P.; Pfleiderer, W.; Charubala, R.; Yakovlev, G.I. A route to 2’,5’-oligoadenylates with increased stability towards phosphodiesterases. FEBS Lett. 1988, 236, 325–328. [Google Scholar] [CrossRef]
- Bramsen, J.B.; Laursen, M.B.; Nielsen, A.F.; Hansen, T.B.; Bus, C.; Langkjaer, N.; Babu, B.R.; Hojland, T.; Abramov, M.; Van Aerschot, A.; et al. A large-scale chemical modification screen identifies design rules to generate siRNAs with high activity, high stability and low toxicity. Nucleic Acids Res. 2009, 37, 2867–2881. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.B.; Pakula, M.M.; Gao, S.; Fluiter, K.; Mook, O.R.; Baas, F.; Langklaer, N.; Wengel, S.L.; Wengel, J.; Kjems, J.; et al. Utilization of unlocked nucleic acid (UNA) to enhance siRNA performance in vitro and in vivo. Mol. Biosyst. 2010, 6, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Bramsen, J.B.; Pakula, M.M.; Hansen, T.B.; Bus, C.; Langkjaer, N.; Odadzic, D.; Smicius, R.; Wengel, S.L.; Chattopadhyaya, J.; Engels, J.W.; et al. A screen of chemical modifications identifies position-specific modification by UNA to most potently reduce siRNA off-target effects. Nucleic Acids Res. 2010, 38, 5761–5773. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, K.; Padmanabhan, K.P.; Ferrara, J.D.; Sadler, J.E.; Tulinsky, A. The structure of alpha-thrombin inhibited by a 15-mer single-stranded DNA aptamer. J. Biol. Chem. 1993, 268, 17651–17654. [Google Scholar] [PubMed]
- Macaya, R.F.; Schultze, P.; Smith, F.W.; Roe, J.A.; Feigon, J. Thrombin-binding DNA aptamer forms a unimolecular quadruplex structure in solution. Proc. Natl. Acad. Sci. USA 1993, 90, 3745–3749. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.A.; Feigon, J.; Yeates, T.O. Reconciliation of the X-ray and NMR structures of the thrombin-binding aptamer d(GGTTGGTGTGGTTGG). J. Mol. Biol. 1996, 256, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S.; Szostak, J.W. In vitro selection of functional nucleic acids. Annu. Rev. Biochem. 1999, 68, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. SELEX—A (r)evolutionary method to generate high-affinity nucleic acid ligands. Biomol. Eng. 2007, 24, 381–403. [Google Scholar] [CrossRef] [PubMed]
- Keefe, A.D.; Cload, S.T. SELEX with modified nucleotides. Curr. Opin. Chem. Biol. 2008, 12, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Koshkin, A.A.; Fensholdt, J.; Pfundheller, H.M.; Lomholt, C. A simplified and efficient route to 2’-O, 4’-C-methylene-linked bicyclic ribonucleosides (locked nucleic acid). J. Org. Chem. 2001, 66, 8504–8512. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.S.; Kumar, T.S.; Wengel, J. LNA 5’-phosphoramidites for 5’→3’-oligonucleotide synthesis. Org. Biomol. Chem. 2010, 8, 5012–5016. [Google Scholar] [CrossRef] [PubMed]
- Barciszewski, J.; Medgaard, M.; Koch, T.; Kurreck, J.; Erdmann, V.A. Locked nucleic acid aptamers. Methods Mol. Biol. 2009, 535, 165–186. [Google Scholar] [PubMed]
- Kuwahara, M.; Obika, S.; Nagashima, J.; Ohta, Y.; Suto, Y.; Ozaki, H.; Sawai, H.; Imanishi, T. Systematic analysis of enzymatic DNA polymerization using oligo-DNA templates and triphosphate analogs involving 2’,4’-bridged nucleosides. Nucleic Acids Res. 2008, 36, 4257–4265. [Google Scholar] [CrossRef] [PubMed]
- Veedu, R.N.; Vester, B.; Wengel, J. Enzymatic incorporation of LNA nucleotides into DNA strands. Chembiochem 2007, 8, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Veedu, R.N.; Vester, B.; Wengel, J. In vitro incorporation of LNA nucleotides. Nucleos. Nucleot. Nucl. 2007, 26, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Veedu, R.N.; Vester, B.; Wengel, J. Polymerase chain reaction and transcription using locked nucleic acid nucleotide triphosphates. J. Am. Chem. Soc. 2008, 130, 8124–8125. [Google Scholar] [CrossRef] [PubMed]
- Veedu, R.N.; Vester, B.; Wengel, J. Novel applications of locked nucleic acids. Nucleic Acids Symp. Ser. (Oxf) 2007, 29–30. [Google Scholar] [CrossRef] [PubMed]
- Veedu, R.N.; Vester, B.; Wengel, J. Efficient enzymatic synthesis of LNA-modified DNA duplexes using KOD DNA polymerase. Org. Biomol. Chem. 2009, 7, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Veedu, R.N.; Vester, B.; Wengel, J. Polymerase directed incorporation studies of LNA-G nucleoside 5’-triphosphate and primer extension involving all four LNA nucleotides. New J. Chem. 2010, 34, 877–879. [Google Scholar] [CrossRef]
- Drabovich, A.; Berezovski, M.; Krylov, S.N. Selection of smart aptamers by equilibrium capillary electrophoresis of equilibrium mixtures (ECEEM). J. Am. Chem. Soc. 2005, 127, 11224–11225. [Google Scholar] [CrossRef] [PubMed]
- Berezovski, M.; Musheev, M.; Drabovich, A.; Krylov, S.N. Non-SELEX selection of aptamers. J. Am. Chem. Soc. 2006, 128, 1410–1411. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, S.D.; Bowser, M.T. In vitro selection of aptamers with affinity for neuropeptide Y using capillary electrophoresis. J. Am. Chem. Soc. 2005, 127, 9382–9383. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, S.D.; Bowser, M.T. In vitro evolution of functional DNA using capillary electrophoresis. J. Am. Chem. Soc. 2004, 126, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Mosing, R.K.; Mendonsa, S.D.; Bowser, M.T. Capillary electrophoresis-SELEX selection of aptamers with affinity for HIV-1 reverse transcriptase. Anal. Chem. 2005, 77, 6107–6112. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, S.D.; Bowser, M.T. In vitro selection of high-affinity DNA ligands for human IgE using capillary electrophoresis. Anal. Chem. 2004, 76, 5387–5392. [Google Scholar] [CrossRef] [PubMed]
- Berezovski, M.; Drabovich, A.; Krylova, S.M.; Musheev, M.; Okhonin, V.; Petrov, A.; Krylov, S.N. Nonequilibrium capillary electrophoresis of equilibrium mixtures: A universal tool for development of aptamers. J. Am. Chem. Soc. 2005, 127, 3165–3171. [Google Scholar] [CrossRef] [PubMed]
- Berezovski, M.V.; Musheev, M.U.; Drabovich, A.P.; Jitkova, J.V.; Krylov, S.N. Non-SELEX: Selection of aptamers without intermediate amplification of candidate oligonucleotides. Nat. Protoc. 2006, 1, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Bassett, S.E.; Li, X.; Luxon, B.A.; Herzog, N.K.; Shope, R.E.; Aronson, J.; Prow, T.W.; Leary, J.F.; Kirby, R.; et al. Construction and selection of bead-bound combinatorial oligonucleoside phosphorothioate and phosphorodithioate aptamer libraries designed for rapid PCR-based sequencing. Nucleic Acids Res. 2002, 30, e132. [Google Scholar] [CrossRef] [PubMed]
- Katilius, E.; Flores, C.; Woodbury, N.W. Exploring the sequence space of a DNA aptamer using microarrays. Nucleic Acids Res. 2007, 35, 7626–7635. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.G.; Platt, M.; Rowe, W.; Wedge, D.C.; Khan, F.; Day, P.J.; McShea, A.; Knowles, J.; Kell, D.B. Array-based evolution of DNA aptamers allows modelling of an explicit sequence-fitness landscape. Nucleic Acids Res. 2009, 37, e6. [Google Scholar] [CrossRef] [PubMed]
- Platt, M.; Rowe, W.; Wedge, D.C.; Kell, D.B.; Knowles, J.; Day, P.J. Aptamer evolution for array-based diagnostics. Anal. Biochem. 2009, 390, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Rowe, W.; Platt, M.; Wedge, D.C.; Day, P.J.; Kell, D.B.; Knowles, J.D. Convergent evolution to an aptamer observed in small populations on DNA microarrays. Phys. Biol. 2010, 7, 036007. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Kumar, R.; Wengel, J. Synthesis of 2’-amino-LNA: A novel conformationally restricted high-affinity oligonucleotide analogue with a handle. J. Org. Chem. 1998, 63, 10035–10039. [Google Scholar] [CrossRef]



© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Doessing, H.; Vester, B. Locked and Unlocked Nucleosides in Functional Nucleic Acids. Molecules 2011, 16, 4511-4526. https://doi.org/10.3390/molecules16064511
Doessing H, Vester B. Locked and Unlocked Nucleosides in Functional Nucleic Acids. Molecules. 2011; 16(6):4511-4526. https://doi.org/10.3390/molecules16064511
Chicago/Turabian StyleDoessing, Holger, and Birte Vester. 2011. "Locked and Unlocked Nucleosides in Functional Nucleic Acids" Molecules 16, no. 6: 4511-4526. https://doi.org/10.3390/molecules16064511