A Novel Anti-Inflammatory Role for Ginkgolide B in Asthma via Inhibition of the ERK/MAPK Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
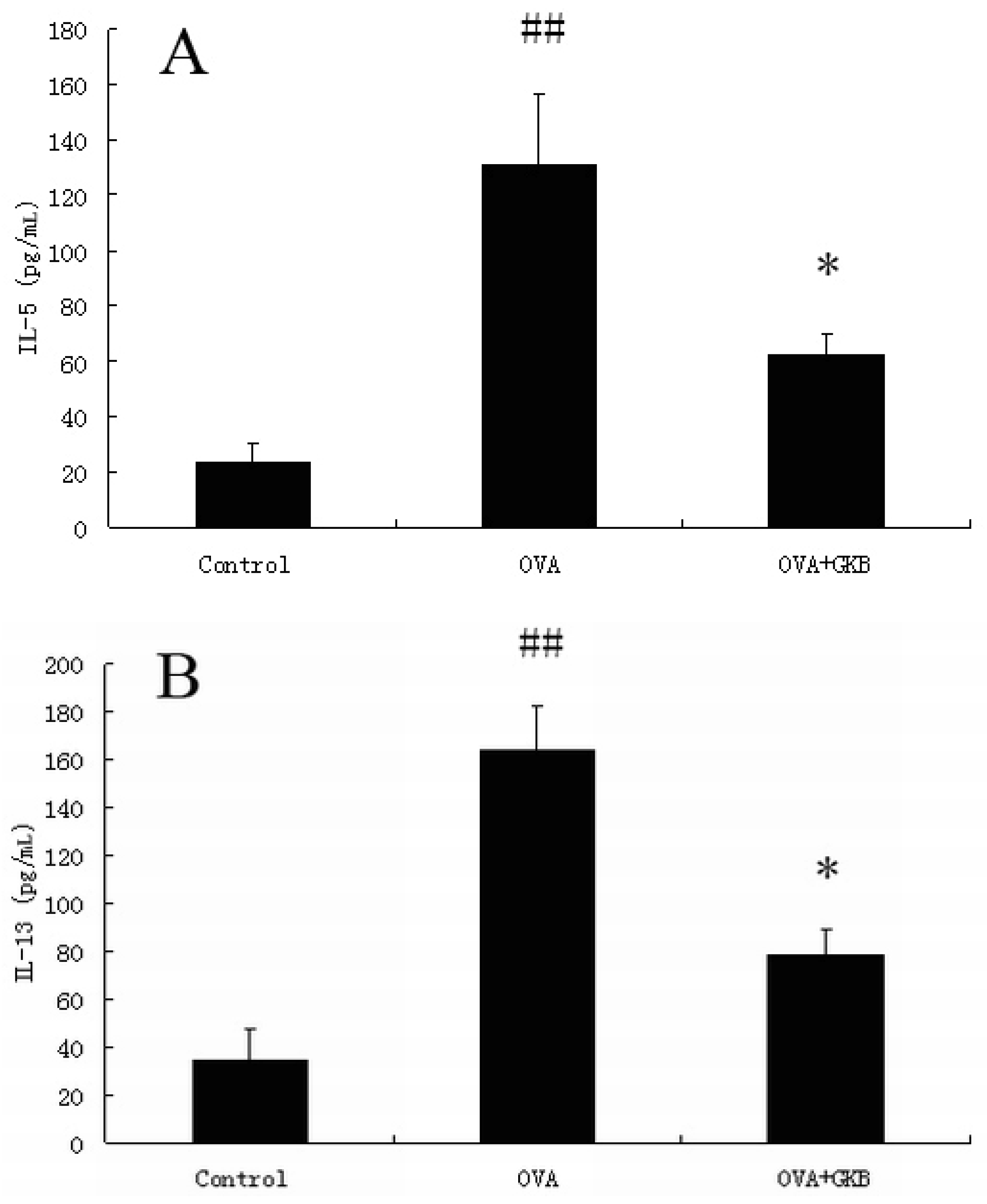
2.1. GKB Reduces Ovalbumin-induced Bronchoalveolar Lavage Fluid T Helper Type 2 Cytokine Levels

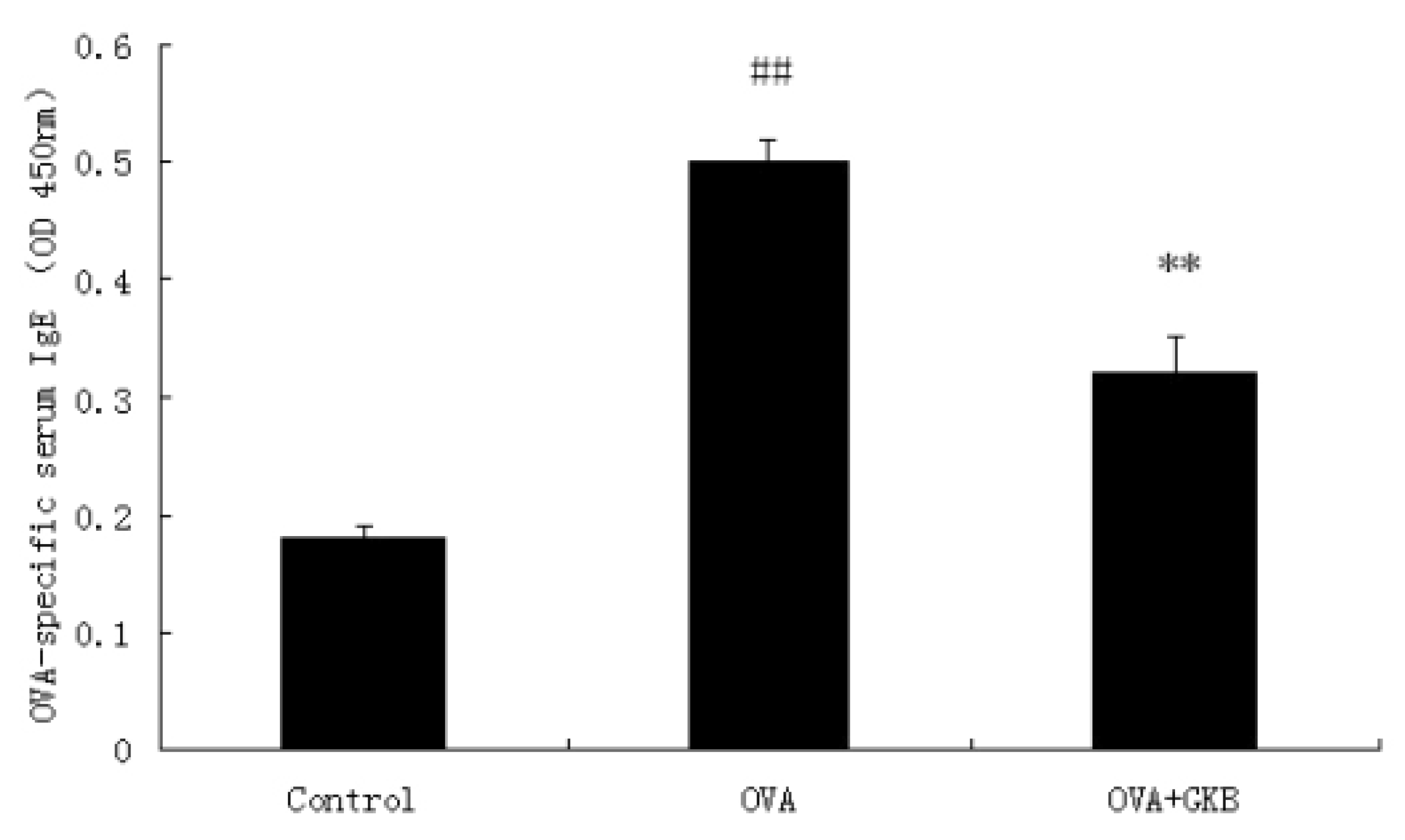
2.2. GKB Reduces OVA-Induced Serum Levels of OVA-specific IgE

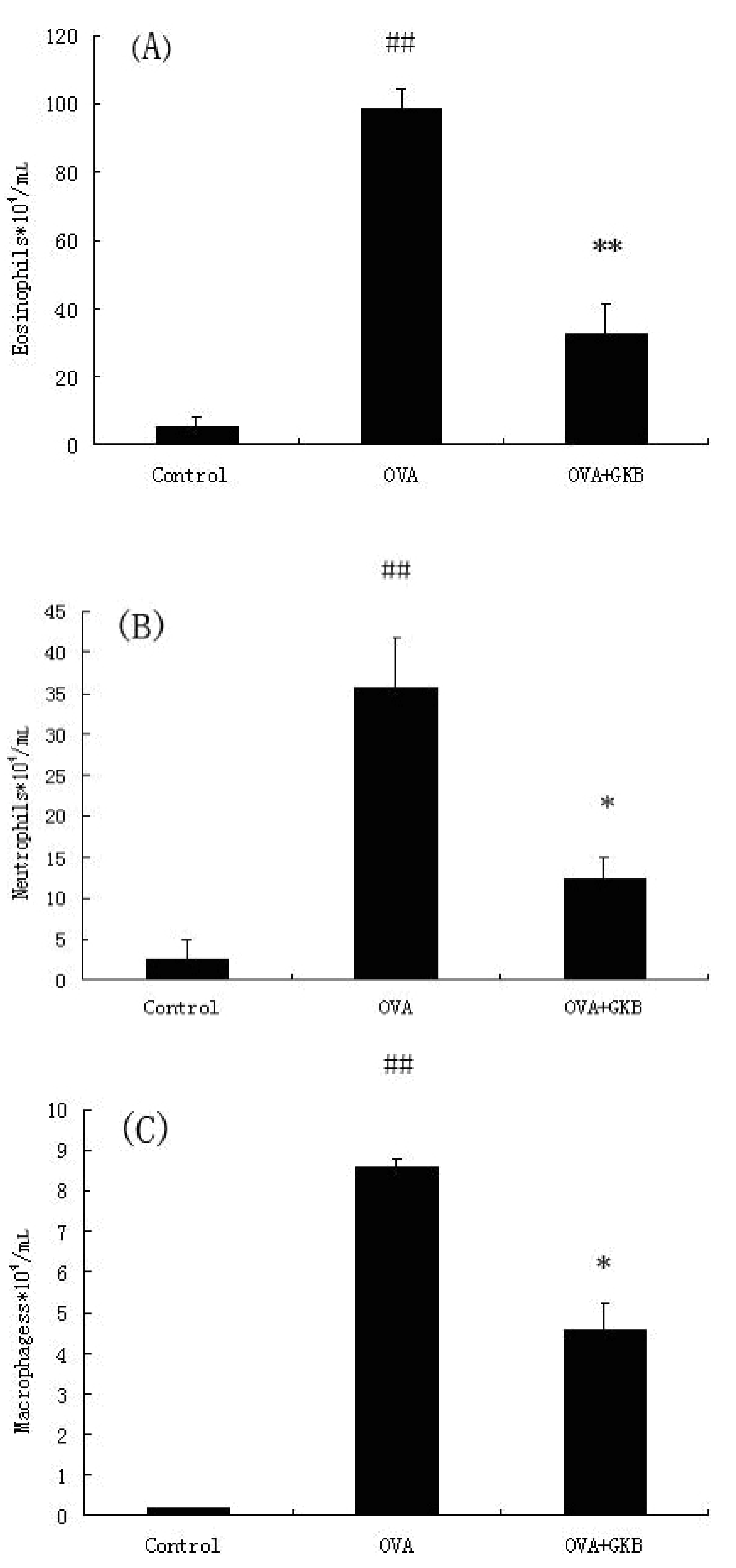
2.3. GKB Reduces OVA-Induced Bronchoalveolar Lavage Fluid (BALF) Inflammatory Cell Recruitment

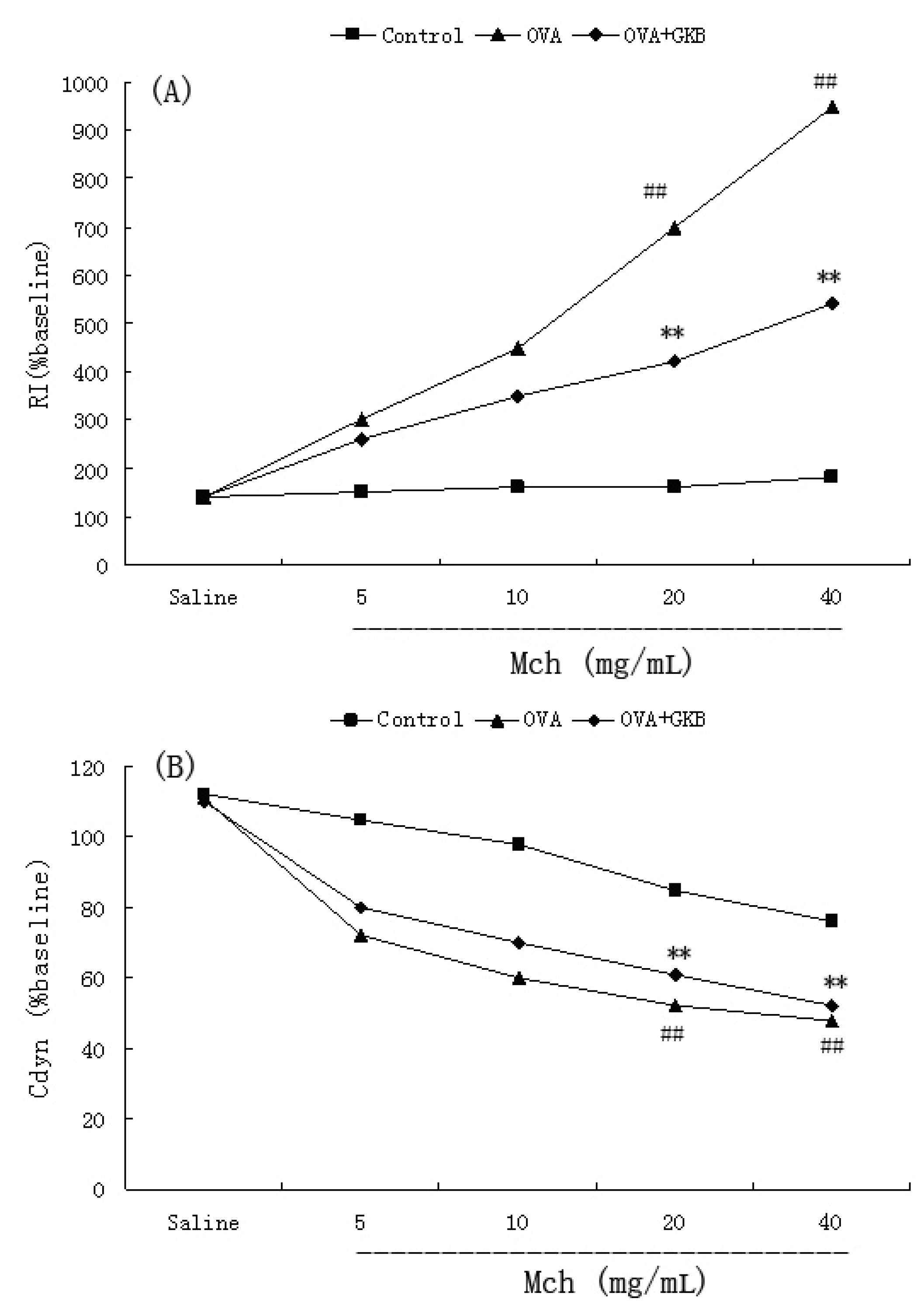
2.4. Effects of GKB on OVA-Induced Airway Hyper-Responsiveness
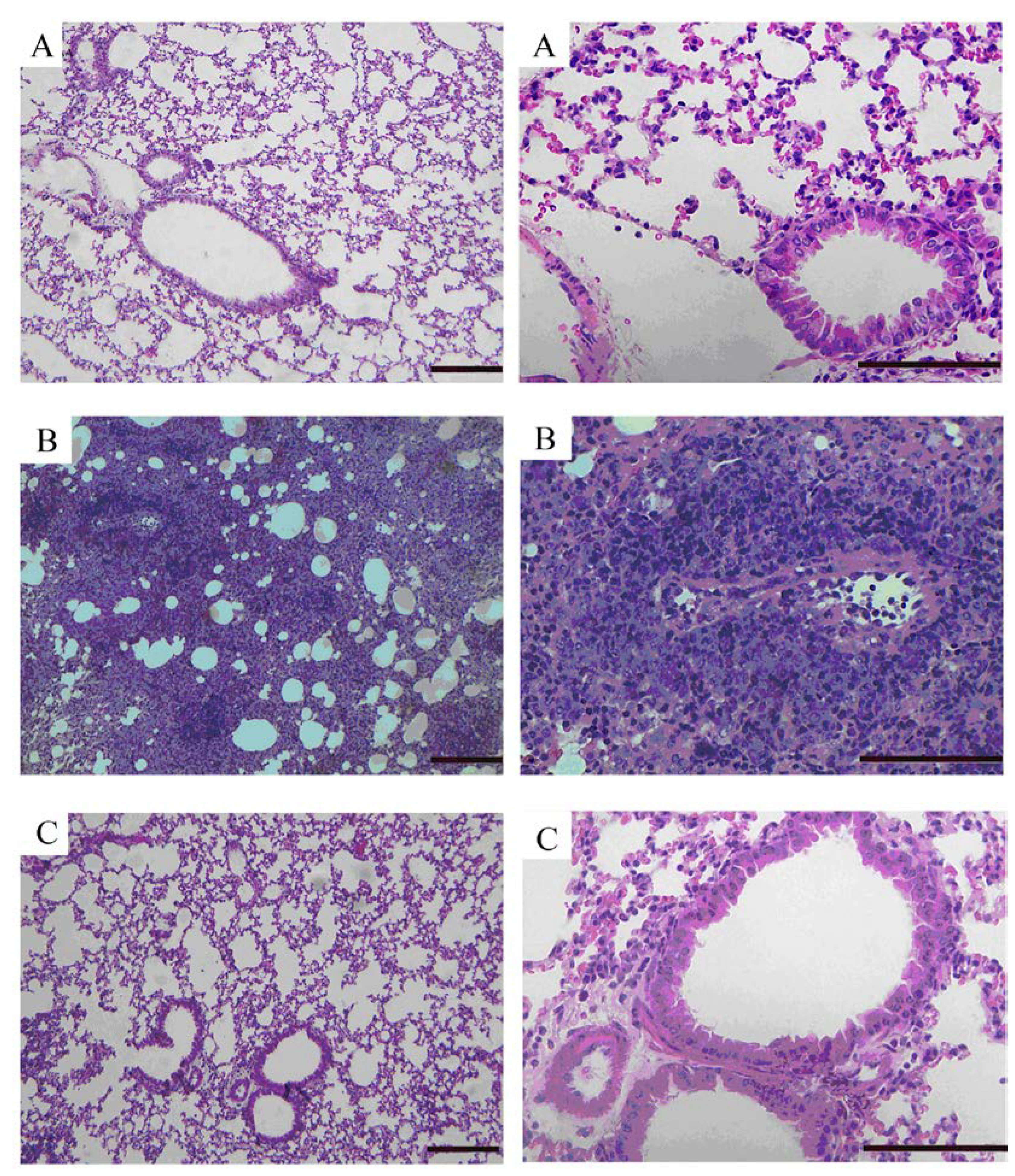
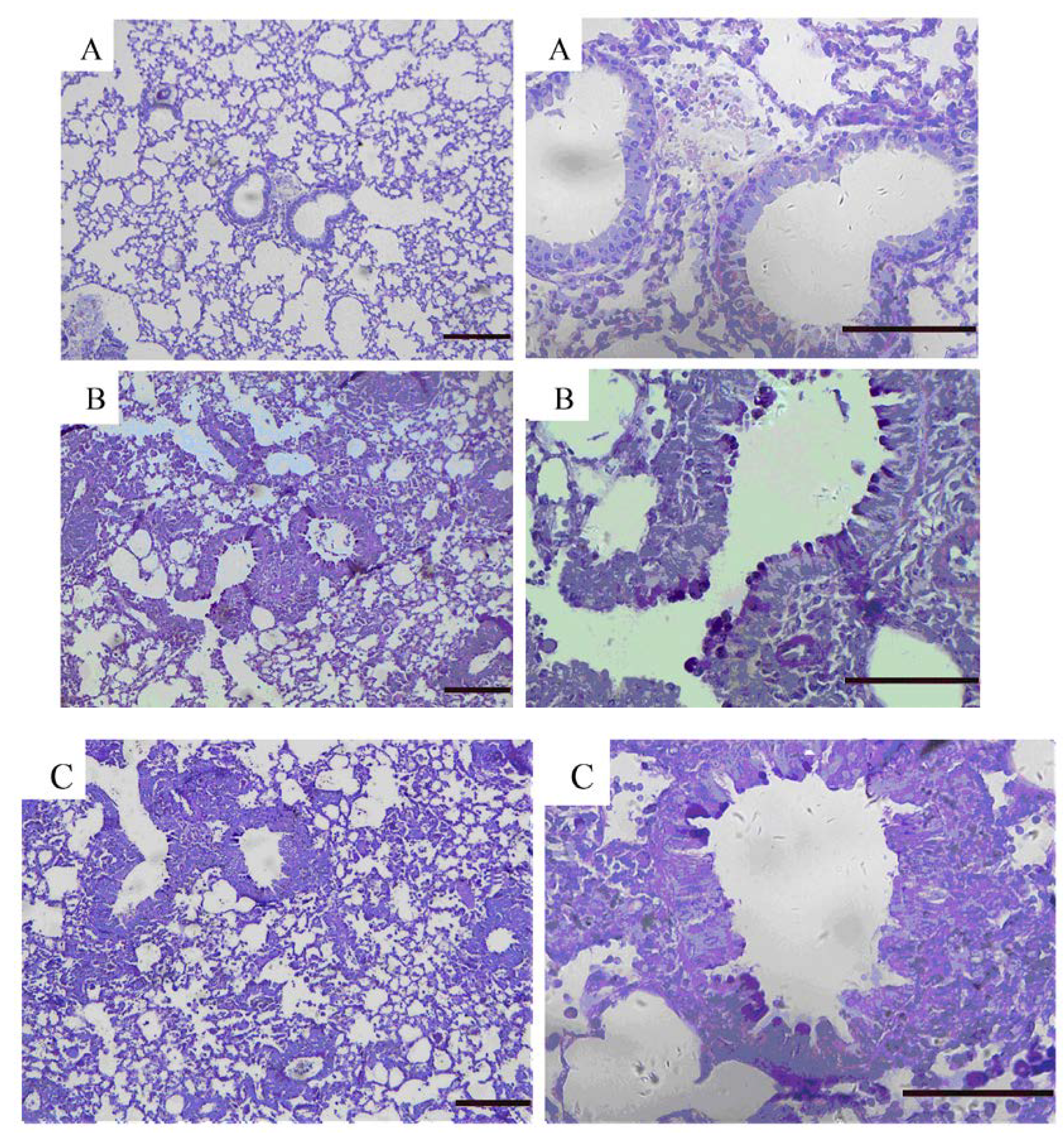
2.5. Effects of GKB on OVA-Induced Airway Goblet Cell Hyperplasia and Mucus Production




2.6. Effects of GKB on Activation of p38, ERK and JNK
2.7. Discussion
3. Experimental
3.1. Animals
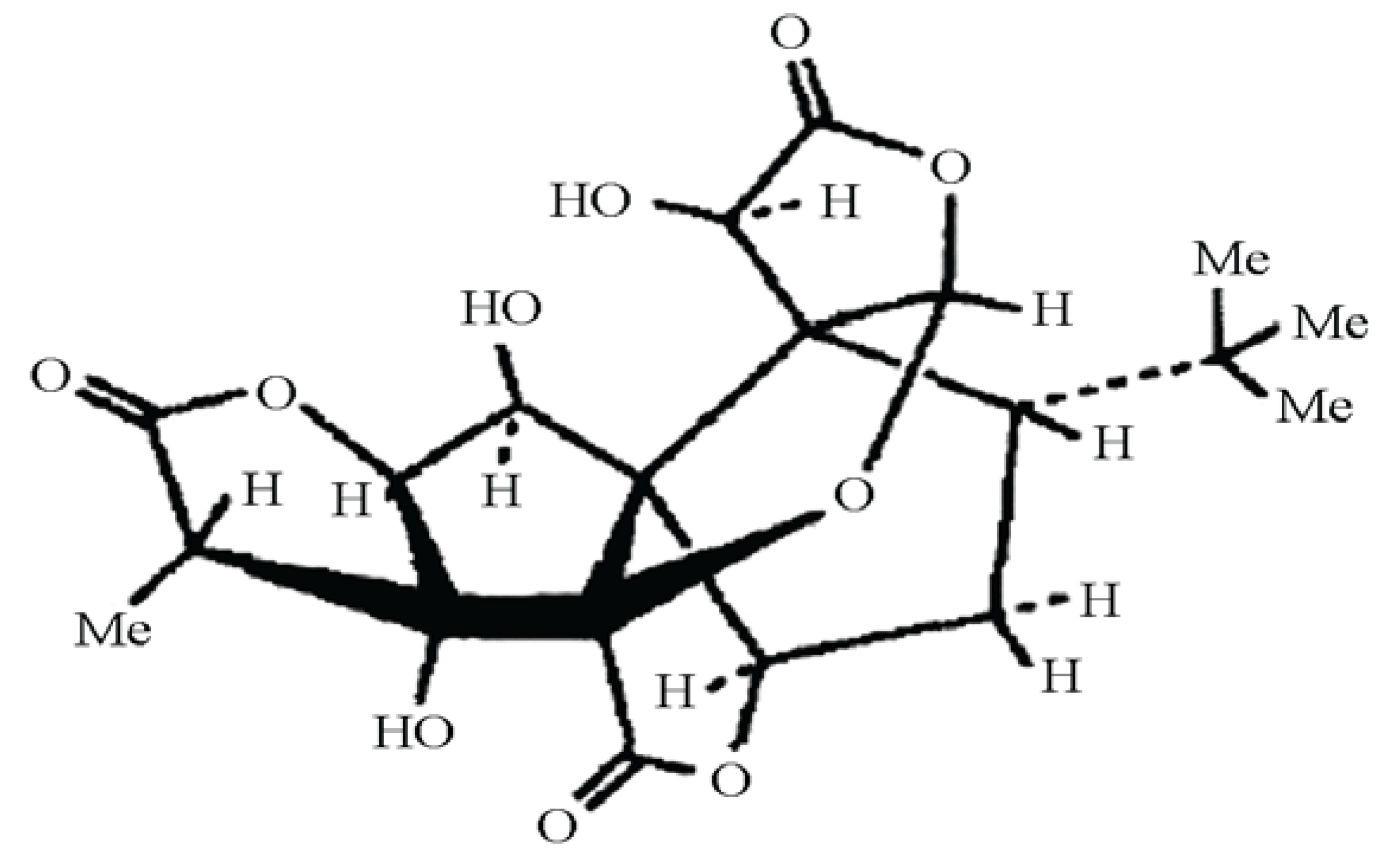
3.2. Reagent

3.3. Antigen Sensitization, Challenge and GKB Treatment
3.4. BALF and Serum Collection
3.5. Cytokine Levels in Lung Tissues
3.6. Mouse Anti-OVA IgE ELISA
3.7. Determination of Airway Hyper-Responsiveness
3.8. Histological Examination
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
- Samples Availability: Samples of the compounds are available from the authors (or from MDPI).
References
- Bloemen, K.; Verstraelen, S.; van Den Heuvel, R.; Witters, H.; Nelissen, I.; Schoeters, G. The allergic cascade: Review of the most important molecules in the asthmatic lung. Immunol. Lett. 2007, 113, 6–18. [Google Scholar] [CrossRef]
- Brightling, C.E.; Symon, F.A.; Birring, S.S.; Bradding, P.; Pavord, I.D.; Wardlaw, A.J. TH2 cytokine expression in bronchoalveolar lavage fluid T lymphocytes and bronchial submucosa is a feature of asthma and eosinophilic bronchitis. J. Allergy Clin. Immunol. 2002, 110, 899–905. [Google Scholar] [CrossRef]
- Sanderson, C.J. Interleukin-5, eosinophils, and disease. Blood 1992, 79, 3101–3109. [Google Scholar]
- Simon, D.; Braathen, L.R.; Simon, H.U. Eosinophils and atopic dermatitis. Allergy 2004, 59, 561–570. [Google Scholar] [CrossRef]
- Lopez, A.F.; Sanderson, C.J.; Gamble, J.R.; Campbell, H.D.; Young, I.G.; Vadas, M.A. Recombinant human interleukin 5 is a selective activator of human eosinophil function. J. Exp. Med. 1988, 167, 219–224. [Google Scholar] [CrossRef]
- Umetsu, D.T.; DeKruyff, R.H. TH1 and TH2 CD4+ cells in human allergic diseases. J. Allergy Clin. Immunol. 1997, 100, 1–6. [Google Scholar] [CrossRef]
- Wills-Karp, M.; Luyimbazi, J.; Xu, X.; Schofield, B.; Neben, T.Y.; Karp, C.L.; Donaldson, D.D. Interleukin-13: Central mediator of allergic asthma. Science 1998, 18, 2258–2261. [Google Scholar]
- Hershey, G.K. IL-13 receptors and signaling pathways: An evolving web. J. Allergy Clin. Immunol. 2003, 111, 677–690. [Google Scholar] [CrossRef]
- Hu, L.; Chen, Z.; Xie, Y.; Jiang, Y.; Zhen, H. Alkyl and alkoxycarbonyl derivatives of ginkgolide B: Synthesis and biological evaluation of PAF inhibitory activity. Bioorg. Med. Chem. 2000, 822, 1515–1521. [Google Scholar]
- Hsuuw, Y.D.; Kuo, T.F.; Lee, K.H.; Liu, Y.C.; Huang, Y.T.; Lai, C.Y.; Chan, W.H. Ginkgolide B Induces Apoptosis via Activation of JNK and p21-Activated Protein Kinase 2 in Mouse Embryonic Stem Cells. Ann. N.Y. Acad. Sci. 2009, 1171, 501–508. [Google Scholar]
- Muntaner, O.; Pedruzzi, E.; Roch-Arveiller, M.; Tissot, M.; Drieu, K.; Périanin, A. Ginkgolide B stimulates signaling events in neutrophils and primes defense activities. Biochem. Biophys. Res. Commun. 2005, 335, 1149–1154. [Google Scholar] [CrossRef]
- Tosaki, A.; Engelman, D.T.; Pali, T.; Engelman, R.M.; Droy-Lefaix, M.T. Ginkgo biloba extract (EGb 761) improves postischemic function in isolated preconditioned working rat hearts. Coronary Artery Dis. 1994, 5, 443–450. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Tsang, F.; Koh, A.H.; Ting, W.L.; Wong, P.T.; Wong, W.S. Effects of mitogen-activated protein kinase inhibitor PD 098059 on antigen challenge of guinea-pig airways in vitro. Br. J. Pharmacol. 1998, 25, 61–68. [Google Scholar]
- Tosaki, A.; Pali, T.; Droy-Lefaix, M.T. Effects of Ginkgo biloba extract and preconditioning on the diabetic rat myocardium. Diabetologia 1996, 39, 1255–1262. [Google Scholar] [CrossRef]
- Haines, D.D.; Varga, B.; Bak, I.; Juhasz, B.; Mahmoud, F.F.; Kalantari, H.; Gesztelyi, R.; Lekli, I.; Czompa, A.; Tosaki, A. Summative interaction between astaxanthin, Ginkgo biloba extract (EGb761) and vitamin C in suppression of respiratory inflammation: a comparison with ibuprofen. Phytother. Res. 2011, 25, 128–136. [Google Scholar] [CrossRef]
- Tosaki, A.; Droy-Lefaix, M.T.; Pali, T.; Das, D.K. Effects of SOD, catalase, and a novel antiarrhythmic drug, EGB 761, on reperfusion-induced arrhythmias in isolated rat hearts. Free Radic. Biol. Med. 1993, 14, 361–370. [Google Scholar] [CrossRef]
- Sanderson, C.J. Interleukin-5, eosinophils, and disease. Blood 1992, 79, 3101–3109. [Google Scholar]
- Dewson, G.; Cohen, G.M.; Wardlaw, A.J. Interleukin-5 inhibits translocation of Bax to the mitochondria, cytochrome c release, and activation of caspases in human eosinophils. Blood 2001, 98, 2239–2247. [Google Scholar] [CrossRef]
- Sher, A.; Coffman, R.L.; Hieny, S.; Cheever, A.W. Ablation of eosinophil and IgE responses with anti-IL-5 or anti-IL-4 antibodies fails to affect immunity against Schistosoma mansoni in the mouse. J. Immunol. 1990, 145, 3911–3916. [Google Scholar]
- Horie, S.; Okubo, Y.; Hossain, M.; Sato, E.; Nomura, H.; Koyama, S.; Suzuki, J.; Isobe, M.; Sekiguchi, M. Interleukin-13 but not interleukin-4 prolongs eosinophi survival and induces eosinophil chemotaxis. Internal Med. 1997, 36, 179–185. [Google Scholar] [CrossRef]
- Schleimer, R.P.; Sterbinsky, S.A.; Kaiser, J.; Bickel, C.A.; Klunk, D.A.; Tomioka, K.; Newman, W.; Luscinskas, F.W.; Gimbrone, M.A., Jr.; McIntyre, B.W. IL-4 induces adherence of human eosinophils and basophils but not neutrophils to endothelium. Association with expression of VCAM-1. J. Immunol. 1992, 148, 1086–1092. [Google Scholar]
- Green, R.H.; Brightling, C.E.; McKenna, S.; Hargadon, B.; Parker, D.; Bradding, P.; Wardlaw, A.J.; Pavord, I.D. Asthma exacerbations and sputum eosinophil counts: A randomized controlled trial. Lancet 2002, 360, 1715–1721. [Google Scholar] [CrossRef]
- Sont, J.K.; Willems, L.N.; Bel, E.H.; van Krieken, J.H.; Vandenbroucke, J.P.; Sterk, P.J. Clinical control and histopathologic outcome of asthma when using airway hyper-responsiveness as an additional guide to long-term treatment. Am. J. Respir. Crit. Care Med. 1999, 159, 1043–1051. [Google Scholar]
- Vargaftig, B.B.; Singer, M. Leukotrienes mediate murine bronchopulmonary hyperreactivity, inflammation, and part of mucosal metaplasia and tissue injury induced by recombinant murine interleukin-13. Am. J. Respir. Cell Mol. Biol. 2003, 28, 410–419. [Google Scholar] [CrossRef]
- Gleich, G.J. Mechanisms of eosinophil-associated inflammation. J. Allergy Clin. Immunol. 2000, 105, 651–663. [Google Scholar] [CrossRef]
- Hamelmann, E.; Tadeda, K.; Oshiba, A.; Gelfand, E.W. Role of IgE in the development of allergic airway inflammation and airway hyper-responsiveness in a murine model. Allergy 1999, 54, 297–305. [Google Scholar] [CrossRef]
- Holgate, S.; Casale, T.; Wenzel, S.; Bousquet, J.; Deniz, Y.; Reisner, C. The anti-inflammatory effects of omalizumab confirm the central role of IgE in allergic inflammation. J. Allergy Clin. Immunol. 2005, 115, 459–465. [Google Scholar] [CrossRef]
- Nel, A.E. T-cell activation through the antigen receptor. Part 1: Signaling components, signaling pathways, and signal integration at the T-cell antigen receptor synapse. J. Allergy Clin. Immunol. 2002, 109, 758–770. [Google Scholar] [CrossRef]
- Jacob, A.; Cooney, D.; Pradhan, M.; Coggeshall, K.M. Convergence of signaling pathways on the activation of ERK in B cells. J. Biol. Chem. 2002, 277, 23420–23426. [Google Scholar]
- Gauld, S.B.; Dal Porto, J.M.; Cambier, J.C. B cell antigen receptor signaling: Roles in cell development and disease. Science 2002, 296, 1641–1642. [Google Scholar] [CrossRef]
- Nadler, M.J.; Matthews, S.A.; Turner, H.; Kinet, J.P. Signal transduction by the high-affinity immunoglobulin E receptor FcεRI: coupling form to function. Adv. Immunol. 2000, 76, 325–355. [Google Scholar]
- Kumar, A.; Lnu, S.; Malya, R.; Barron, D.; Moore, J.; Corry, D.B.; Boriek, A.M. Mechanical stretch activates nuclear factor-κB, activator protein-1, and mitogen-activated protein kinases in lung parenchyma: implications in asthma. FASEB J. 2003, 17, 1800–1811. [Google Scholar] [CrossRef]
- Duan, W.; Chan, J.H.; Wong, C.H.; Leung, B.P.; Wong, W.S. Anti-inflammatory effects of mitogen-activated protein kinase kinase inhibitor U0126 in an asthma mouse model. J. Immunol. 2004, 172, 7053–7059. [Google Scholar]
- Bao, Z.; Lim, S.; Liao, W.; Lin, Y.; Thiemermann, C.; Leung, B.P.; Wong, W.S. Glycogen synthase kinase-3beta inhibition attenuates asthma in mice. Am. J. Respir. Crit. Care Med. 2007, 176, 431–438. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Biotechnology 1992, 24, 145–149. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chu, X.; Ci, X.; He, J.; Wei, M.; Yang, X.; Cao, Q.; Li, H.; Guan, S.; Deng, Y.; Pang, D.; et al. A Novel Anti-Inflammatory Role for Ginkgolide B in Asthma via Inhibition of the ERK/MAPK Signaling Pathway. Molecules 2011, 16, 7634-7648. https://doi.org/10.3390/molecules16097634
Chu X, Ci X, He J, Wei M, Yang X, Cao Q, Li H, Guan S, Deng Y, Pang D, et al. A Novel Anti-Inflammatory Role for Ginkgolide B in Asthma via Inhibition of the ERK/MAPK Signaling Pathway. Molecules. 2011; 16(9):7634-7648. https://doi.org/10.3390/molecules16097634
Chicago/Turabian StyleChu, Xiao, Xinxin Ci, Jiakang He, Miaomiao Wei, Xiaofeng Yang, Qingjun Cao, Hongyu Li, Shuang Guan, Yanhong Deng, Daxin Pang, and et al. 2011. "A Novel Anti-Inflammatory Role for Ginkgolide B in Asthma via Inhibition of the ERK/MAPK Signaling Pathway" Molecules 16, no. 9: 7634-7648. https://doi.org/10.3390/molecules16097634